
進化精神医学:進化と精神衛生に関する現在の展望
リアド・アベッド
進化精神医学は、進化論的および適応論的理論のレンズを通して、精神疾患の発展と有病率を説明し検討することを試みます。この編集された巻では、国際的な進化学者たちが、精神疾患が「どのように」発生するかだけでなく「なぜ」発生するかを考えるよう読者を促す様々なダーウィニズム的視点を提示しています。比較動物進化学、動物行動学、人類学、文化、哲学、その他の人文科学からの洞察を用いて、進化的思考は精神医学の疫学、遺伝学、生化学、心理学を再評価するのに役立ちます。それは、選択とその他の進化プロセスによって形成された持続的で遺伝する特性の説明を求め、系統発生の歴史と神経科学からの洞察、そして現代環境の影響を用いて特性や障害をレビューします。精神医学への社会的アプローチと生物学的アプローチの間のギャップを埋め、主流の精神医学に進化的視点を取り入れることを奨励することにより、この本は精神疾患の原因と治療に関する新たな研究の道を開くのに役立つでしょう。
目次 執筆者一覧 序文 ジョージ・イッコス はじめに 1 進化精神医学入門 リアド・アベッドとポール・セント・ジョン-スミス 2 進化論によって進化した生物心理社会モデル アダム・ハント、ポール・セント・ジョン-スミス、リアド・アベッド 3 ホミニン進化 I:ホモ・サピエンスの起源 デレク・K・トレイシー 4 ホミニン進化 II:サピエンス、既知の宇宙のマスター デレク・K・トレイシー 5 狩猟採集民、ミスマッチ、そして精神障害 ニキル・チョードリーとグル・デニズ・サラリ 6 なぜ精神障害は持続するのか?精神医学の進化的基盤 ランドルフ・M・ネッセ 7 進化的視点から見た不安障害 ランドルフ・M・ネッセ 8 うつ病に関する進化的視点 マルクス・J・ランタラとセヴェリ・ルオト 9 自殺のランダム性について:自殺リスク評価を超えるための進化的、臨床的呼びかけ C・A・ソパー、パブロ・マロ・オセホ、マシュー・M・ラージ 10 統合失調症スペクトラム障害に関する進化的視点 マーティン・ブリューネ 11 摂食障害に関する進化的視点 リアド・アベッドとアグネス・エイトン 12 物質乱用と進化 ポール・セント・ジョン-スミスとリアド・アベッド 13 進化的視点から見たアルコールの社会的機能 ロビン・I・M・ダンバー 14 小児期トラウマに関する進化的視点 アニー・スワネポエル、マイケル・J・ライス、ジョン・ローナー、グラハム・ミュージック、バーナデット・レン 15 神経発達障害に関する進化的視点 アニー・スワネポエル、マイケル・J・ライス、ジョン・ローナー、グラハム・ミュージック、バーナデット・レン 16 母親のネガティブさと子どもの虐待:進化的視点が重層的で思いやりのある理解にどのように貢献するか ダニエラ・F・シーフ 17 生殖的ライフヒストリーに焦点を当てた進化的ミスマッチの疾患としてのアルツハイマー病 モリー・フォックス 18 精神薬理学と進化 ポール・セント・ジョン-スミス、リアド・アベッド、マーティン・ブリューネ 19 進化的・認知科学が犯罪と司法の科学に提供するもの ブライアン・B・ブートウェル、メーガン・スプラナント、トッド・K・シャケルフォード 20 精神疾患患者の臨床ケアにおける進化的思考 アルフォンソ・トロイシ
序文
当時の医学的環境でよくあったように、エミール・クレペリン(1856-1926)(Kraepelin, 1987)とジークムント・フロイト(1856-1939)(Sulloway, 1980)はともに人間の進化に強い関心を持っていました。クレペリンは大学の学部で精神医学的遺伝学の追求を先駆け、フロイトは晩年に精神分析の訓練において進化論の教育を明確に提唱しました。残念ながら、優生学の残酷さと社会的ダーウィニズムの悪意は、重度の神経精神医学的障害を持つ何万人もの障害のある市民の去勢(Porter, 1999)や殺害(Friedlander, 1995)を含む忌まわしい慣行へとつながりました。それにもかかわらず、精神医学的症候群の疫学における家族的要因の継続的な確認とDNAの理解における科学的革命は、精神医学的遺伝学への積極的な関心を維持するのに役立ちました。おそらく、そして恐らく理解できることですが、忌まわしい遺産と「つじつま合わせの話」(第6章参照)に対する正当な恐れ、さらに遺伝子技術の刺激的な進歩が、遺伝子への焦点を狭め、私たちの専門分野においてより広範な進化的視点を周辺化させました。臨床実践のニーズに導かれたこの広範囲にわたる巻は、バランスを是正するのに役立つでしょう。重要なことに、すべての章は明確に書かれ、精神科研修医(および関心のある他の人々)を引き込むレベルで設定されており、将来の同僚が診療所やその他の環境で出会う多様で複数の要因によって決定される現象についての深い理解を発展させるのに役立ちます。
主題の歴史を考慮すると、編者が単なる理論や解釈ではなく、仮説検証に強くコミットしていることを注目することが重要です。彼らはまた、精神的健康と疾患の生物学的理解と社会的理解の架け橋として進化論が果たす可能性について書いています – これは必要な野心です。しかし、ベネディクト・スピノザ(Lloyd, 1996)が400年以上前に主張し、これらの章が明らかにしているように、物質と心(生物学的なものと社会的なもの)はコインの表裏であり、コインと同様に、私たちは両面を同時に見ることができないように構成されています。私たちのこの認知的、感情的な欠点は、より野心的な目標への障害となっています:つまり、単に生物心理社会的なアプローチではなく、精神医学と精神保健サービスへの真に多元的なアプローチです。この巻の強みの一つ – ただしそれだけではありません – は、生物学、心理学、人類学、哲学の視点からの貢献を含む学際的な著者構成です。多元主義に関して仮説検証型の進化精神医学が約束するものは、新しい理解だけでなく、新しい問いかけです。
記録のために、そしてこの序文に寄稿するよう招待された唯一の資格として、私はポール・セント・ジョン-スミス博士を研修医、専門家仲間、共著者、知的スパーリングパートナー、進化精神医学のメンター、そして友人として何十年も知る幸運に恵まれました。私は彼に、そして今ではリアド・アベッド博士にも、私の継続的な専門的形成について恩義を感じています。2013年5月にロンドンのロイヤル・ソサエティ・オブ・メディシンで、私がアンジェロス・チャニオティス教授とクリストス・シデラス博士と共同で企画したイベント「感情と精神医学:神経科学、歴史と文化」の間に、ランドルフ・M・ネッセ教授を「愛着とうつ病の進化生物学」について講演するよう招待することをポールが提案しました。これがリアドを引き寄せ、彼とポールは知り合いになりました。彼らの共通の熱意は、王立精神科医師会の進化精神医学特別利益グループの設立につながり、これは世界中でそのような関心を持つ最も重要な精神医学組織に発展しました。ほぼ10年にわたる共同の努力の重要な成果は、この巻であり、その長所には、精神科医やその批評家によるものであれ、単純化に挑戦することが含まれています。
ジョージ・イッコス 王立国立整形外科病院
References
Friedlander, H. (1995) The Origins of Nazi Genocide: From Euthanasia to the Final
Solution. Chapel Hill: University of North Carolina Press.
Kraepelin, E. (1987) Memoirs. Berlin: Springer Verlag.
Lloyd, G. (1996) Spinoza and the Ethics. Abingdon-on-Thames: Routledge.
Porter, D. (1999) The quality of population and family welfare: human
reproduction, eugenics and social policy. In: Health, Civilization and the State:
A History of Public Health from Ancient to Modern Times. Abingdon-on-Thames:
Routledge, pp. 165-195.
Sulloway, F. J. (1980) Part two: Psychoanalysis: the birth of a genetic
psychobiology. In Freud, Biologist of the Mind: Beyond the Psychoanalytic Legend.
Cambridge, MA: Harvard University Press, pp. 135-416.
序文
リアド・アベッドとポール・セント・ジョン-スミス
このように、自然の戦いから、飢餓と死から、私たちが考えることのできる最も崇高な対象、すなわち高等動物の産出が、直接的に生じる。この生命観には壮大さがある。その様々な力は、最初はわずかな形態もしくは一つの形態に息を吹き込まれ、この惑星が重力の固定された法則に従って循環する間に、とても単純な始まりから最も美しく最も素晴らしい無限の形態が進化してきた、そして今も進化している。
-チャールズ・ダーウィン『種の起源』(第1版、1859年)
チャールズ・ダーウィンの重要な発見、つまり自然選択が複雑な特性が形作られ、生存と繁殖に有利な特性をその代替案よりも優先することによって種が差別化される手段であるという発見は、地球上の生命の複雑さと多様性について、初めて完全に自然主義的な、奇跡的でない説明を提供しました。しかしながら、自然選択はこのプロセスに関わる唯一のメカニズムではありません。偶然、特に確率的性質は、進化のプロセスにおいて中心的で、さらには本質的な役割を果たしています。特に、選択が作用しなければならない変異の唯一の源である突然変異のランダムなプロセスを通じて – フランスの生化学者でノーベル賞受賞者のジャック・モノーが「人間は計り知れない数の偶然の出来事の産物だった」と宣言したほどです(キャロル、2020:15より引用)。しかし、そのランダムな始まりにもかかわらず、さらに先には選択における強力な非ランダムな要素も存在します(ドーキンス、1996)。
真のダーウィン的なスタイルで、偶然の出来事は、両編者の最近の他の多くの進化的な取り組みと同様に、この巻の制作においても重要な役割を果たしました。最近まで、私たちは二人とも長年にわたり、英国の異なる地域でNHS(国民保健サービス)内で臨床医として働きながら、比較的孤立した状態で進化と精神衛生についての長期的な関心を追求してきました。しかし、2013年にジョージ・イッコスが企画し、ランドルフ・M・ネッセが講演したロイヤル・ソサエティ・オブ・メディシンでの一日会議に私たち二人が出席した後、状況は劇的に変わりました。ポールはリアドの名札から、彼が以前目にした強迫性障害と統合失調症に関する進化論的論文の著者だと認識し、自己紹介しました。それ以降、私たちは連絡を取り合い、会議、論文、書籍の章を含む多くの進化的プロジェクトで協力してきました。重要なことに、私たちは同僚とともに、王立精神科医師会の進化精神医学特別利益グループ(EPSIG)の設立にも協力し、それは2016年に日の目を見ました。そして同僚たちによる(そして時には私たち自身の予感による)精神科医は進化論を避けるだろうという多くの暗い予測に反して、EPSIGのメンバーシップは成長し続け、現在では1,900人を超えています。現在、世界中で最大の進化精神医学グループとなっています。それ以来、私たちは同僚と協力して、世界中の著名な進化論者が自分の研究を発表する4つの一日国際進化精神医学シンポジウムを組織し、いくつかの半日の科学的会議とウェビナーを開催し、YouTubeチャンネルを設置し、精神科研修医と医学生のためのチャールズ・ダーウィン・エッセイ賞を設立し、20以上のニュースレターを発行しました。この巻は、この協力の最新の成果です。もともとはEPSIG設立5周年を記念するものでしたが、出版される頃には6周年と重なりそうです。
興味深いことに、この巻の出版年(2022年)は、再び純粋な偶然により、ダーウィン主義者、特に人間の心理学に関心のある人々にとって幸運な機会と一致しています。つまり、チャールズ・ダーウィンの人間の心理学に関する唯一の著書『人間と動物における感情の表現』の出版150周年です。ダーウィンが今日生きていたら、彼は自分の世界を変える理論が精神障害の理解に適用されているのを見て喜ぶだろうと思います。しかし、他の生命科学とは異なり、精神医学や医学全般が進化を主流に取り入れるのがとても遅かったことに失望するかもしれません。
私たちは、精神医学の現在のダーウィン以前の状態は不幸な時代錯誤であり、この分野の将来の発展の障害だと考えています。したがって、EPSIGの目標は、世界中の進化論的志向を持つ他の精神科医と協力して、これを是正することです。しかし、私たちはこれには時間と努力が必要であることを十分に認識しています。ほとんどの精神科医(そして一般的に医師)は、進化を事実として、そして人間の起源に関する唯一の科学的説明として受け入れていたとしても、機能、機能不全、健康、疾病の理解に対する進化の深遠な結果についてほとんど考えを巡らせていません。その結果、進化精神医学は臨床医の間でマイノリティの関心事にとどまっており、世界中に散らばっているほんの数十人の精神科医だけが、精神障害の進化的理解を進めるために多大な時間とエネルギーを費やしています。これは進化の現実を受け入れている精神科医のほんの一部に過ぎません。
私たちは、多くのさらなる精神科医が、進化精神医学の利点と可能性についてより良く知るようになれば、進化精神医学の事業に加わる可能性が高いと信じています。人間の知識と技術を進歩させる上での人口規模(および臨界質量)の重要性はよく知られています。例えば、初期のヨーロッパ人の接触時期頃のオセアニアの島嶼集団に関する考古学的研究は、小さな人口を持つ島々の人々がより単純な海洋採集技術を持っていたことを示しました(Kline and Boyd, 2010)。また、都市の人口規模が知識やアイデアの生産と交換と正の相関関係にあることも知っています(Bettencourt and West, 2010)。したがって、知識の発展と蓄積に関しては、進化精神医学コミュニティの規模が重要です。そのため、国際的に精神科医の間で進化的知識がより広く普及することで、進化理論モデルの範囲と洗練さの指数関数的な拡大と、そのようなモデルに基づいた新しい、臨床的に関連のある実証データにつながると期待しています。この本が、世界中の進化精神医学者(この分野の多くの先駆者を含む)のアイデアと研究を紹介し、それらを一冊の本で簡単にアクセスできるようにすることで、この極めて重要なプロセスに貢献することを望んでいます。
私たちの目的は、精神医学、心理学、人類学、哲学など複数の分野にまたがる精神衛生と人間心理学に関する幅広い進化的思考の断面を読者に提示することです。進化科学はこの巻のすべての著者にとって基礎的な枠組みを表していますが、精神障害そのものの性質を含む様々な問題について意見の相違の余地は十分にあります。しかし、私たちはこれを弱点ではなく強みの表れと考えています。なぜなら、進化精神医学は理論と仮説を提唱し、それらの予測を検証し、経験的データに照らして受け入れ、拒絶し、または改良していくことで進歩する科学だからです。したがって、私たちの進化的な取り組みは、その性質上、修正可能で動的であるとともに、暫定的で進歩的でなければなりません。真の科学は教義でも固定された情報の集合体でもありません。進化精神医学は簡潔で、有用で、実用的であるとともに、内部的にも外部的にも一貫し、重要なことに、倫理的でなければなりません。私たちはこれらの目標を目指しています。したがって、進化的アプローチが主流に対して批判的である一方で、現在の精神医学の知識と実践を置き換えるのではなく、それらを補完することを目指していることを強調することが重要です。進化論者は現代医学の基礎を形成する科学的方法を完全に支持し、もしあるとすれば、それを弱体化するのではなく強化することを目指しています。しかし、進化論者は現在の精神医学と医学が近位(機械論的)説明とモデルに排他的に焦点を当て、究極的または進化的原因を無視していることを批判しています(第1章と第2章参照)。したがって、私たちは進化が精神医学にとって哲学が科学にとってそうであるものだと提案します:正しい質問をするための枠組みであり、既製の答えを得るための手段ではありません。答えは経験的証拠を分析することによってのみ得られるでしょう。
したがって、この巻は特定の精神障害の性質と治療に関する既製のレシピを提供するものではなく、人間の苦悩と苦しみを探索し理解するための思考方法と枠組みを促進し提唱するものです。進化の強みは、人間を地球上の生命の複雑なウェブの中にしっかりと位置づけ、それから離れたものとしてではなく、祖先の環境で果たした機能の観点から私たちの種の心理学的特性とそれらが生じさせる脆弱性についての理解を提供することです。
将来については、この本が精神科医だけでなく、心理学者、看護師、その他の精神保健専門家、そして心理学的志向を持つ医療・介護職の人々にも興味を持ってもらえることを望んでいます。また、哲学、人類学、生物学など様々な分野の学者や、進化について情報を持つ一般の方々にも興味を持ってもらえることを望んでいます。私たちは、進化的思考が精神機能不全のスティグマを解消するのに役立つと主張しており、これが実際にそうであるという臨床経験を持っています。特に、精神医学と関連職種の研修生がこのテーマに関心を持ち、進化的思考が最終的に標準的なカリキュラムと試験の一部になることを望んでいます。
この巻の章は、それぞれの主題についての独立した進化的レビューとして順不同で読むことができますが、進化文献に不慣れな読者は最初に序章を読むか、さらに良いのは最初の5章を読むことで恩恵を受けるでしょう。これらは読者に進化医学と人間の起源の基礎についての良い基盤を与え、精神障害の理解に進化的思考を適用することの主要な利点についての説明を提供するはずです。
私たちは、このプロジェクトを支援してくれた多くの情報源からの支援と励ましに感謝しています。進化医学の分野の共同創設者であるランドルフ・M・ネッセは、時間とアドバイスを惜しみなく提供してくれました。彼は2016年の私たち最初のEPSIGシンポジウムで講演することを申し出て、旅行計画を変更してロンドンに立ち寄り、私たちが彼の経費をカバーする資金がなかった時に講演を行いました。それ以来、私たちは彼と連絡を取り合い、他の会議の場で会い、彼の賢明なアドバイスから恩恵を受け続けています。さらに、私たちはEPSIGの最初の創設から励ましをくれ、4回の国際シンポジウムのそれぞれでセッションの議長を務めてくれたジョージ・イッコスに感謝しています。特に、現在のEPSIGニュースレター編集者であり、同僚や一般の人々の間で進化を促進することに関連するすべての事柄について私たちの親密な協力者であるアニー・スワネポエルに感謝しています。また、EPSIG執行委員会の同僚と、私たちのシンポジウムで講演したり、この巻に貢献したりした英国および世界中の進化論的同僚たちに、彼らの進化的研究と知識を共有してくれたことに感謝しています。また、この取り組みを組織するよう励ましてくれただけでなく、これらのアイデアの段階的な臨床的展開を支援してくれている、私たちのEPSIGの同僚たちに感謝します。また、私たちは「巨人の肩の上に立っている」¹ということを認識し、進化と精神医学に関する書籍の著者、特にランドルフ・M・ネッセ、ジョン・プライス、アルフォンソ・トロイシ、マーティン・ブリューネ、マルコ・デル・ジュディチェの多大な貢献を認識しています。彼らの書籍がなければ、この分野は大幅に縮小していたでしょう。
さらに、この題名の出版に同意してくれたケンブリッジ大学出版局と王立精神科医師会出版委員会、そして肯定的なフィードバックと励ましを提供してくれた匿名の査読者に特別な感謝を表します。さらに、EPSIGでの会員との連絡を維持し、進化的イベントの組織を手伝ってくれた、王立精神科医師会の過去と現在の管理スタッフ、それぞれキャサリン・ラングレーとケルシー・ヘンシェルに感謝します。また、私たちの章を親切に校正してくれたリンジー・エドワーズ、表紙のイラストとEPSIGのロゴをデザインし草案を作成してくれたハーレー・セント・ジョン-スミス、そして物質乱用に関する第12章をレビューしてくれたデビッド・ギーニーにも感謝します。最後に、この巻の制作において助けとサポートを提供してくれたケンブリッジ大学出版局のオリビア・ボールト、ジェシカ・パップワース、サスキア・プロンクに感謝します。
References
Bettencourt, L. and West, G. A. (2010). A unified theory of urban living. Nature,
467,912-913.
Carroll, S. B. (2020). A Series of Fortunate Events. Princeton, NJ: Princeton
University Press.
Dawkins, R. (1996). Climbing Mount Improbable. London: Norton.
Kline, M. A. and Boyd, R. (2010). Population size predicts technological complexity
in Oceania. Proceedings of the Royal Society B: Biological Sciences, 277, 2559-2564.
1 ‘If we see more and farther than our predecessors, it is not because we have
keener vision or greater height, but is because we are lifted up and borne aloft
on their gigantic stature’ (Bernard of Chartres, twelfth century, quoted by Isaac
Newton, 1675).
第1章
進化精神医学入門
リアド・アベッドとポール・セント・ジョン-スミス*
要旨
この序章は複数の目的を果たしています。その主な目的は、ダーウィン的思考に馴染みのない精神科医やその他の精神保健専門家に、進化科学の基本的な概念と用語のいくつかを紹介し、この巻の残りの章を通じての進行を容易にすることです。もう一つの目的は、精神医学と心理学に関連する様々な進化的分野における重要な理論的およびその他の発展の要約と更新を提供することであり、これは既存の進化論者を含むすべての読者に利益をもたらすでしょう。スペースの制約上、必然的に重要な省略があるでしょう。私たちは標準的な進化理論の基礎、および進化心理学と医学の基本原則のいくつかをカバーすることを選択しました。また、文化進化と関連分野に関する進化文献の最近の発展のいくつかを簡単に調査しています。章が用語集にならないように、可能な限り広い領域をカバーすることとバランスを取る必要があることを認識しています。専門的な進化用語に馴染みのない読者は、王立精神科医師会のウェブサイト上の進化精神医学特別利益グループの用語集を参照することをお勧めします:www.epsig.org (「About us」をクリックし、次に「Resources」)。
要点
· ダーウィン理論はすべての生命科学の組織的枠組みである。
· 進化的思考は、ティンバーゲンの4つの質問の適用を通じて、医学と精神医学における因果関係の理解を変革することができる。
· 進化なしでは、疾患の原因に関する私たちの理解は必然的に不完全である。
· 進化的視点は、疾患や障害に対する人間の脆弱性を理解するのに役立つ。
· 進化精神医学は主流の精神医学を補完し強化するものであり、それを置き換えようとするものではない。
· 進化はまた、人間の独自性、特に人間の体と心を形成する累積的文化と遺伝子-文化共進化の役割を理解するのに役立つ。
1.1 進化理論入門
1.1.1 背景 チャールズ・ダーウィンは1859年に二つの異なる革命的な提案をしました。第一に、すべての生物は共通の祖先を共有しているということ、第二に、自然選択が地球上のすべての生命の多様性が生じるメカニズムであるということでした(Nesse and Stein, 2019)。
これらの洞察は歴史上最大の科学革命の一つを動かしました。他の主要な科学的パラダイムシフトは物理科学で発生しましたが(例えば、コペルニクス、ニュートン、アインシュタイン、ハイゼンベルクのもの)、それらは専門分野以外では目立った影響をほとんど持ちませんでした。しかし、ダーウィニズムは、生物学から地質学に至るまでの複数の探究と信念の分野で深く根付いた前提に挑戦し、創造論、本質主義、人間中心主義への挑戦においてメタ科学的な結果を持ちました(Mayr, 1971)。しかし、生命科学の一部であるにもかかわらず、精神医学(および医学の多く)はそのアプローチにおいて大部分が前ダーウィン的なままです。この本では、精神科医、哲学者、人類学者、心理学者を含む様々な分野の進化学者が、現在の主流の精神医学を拒絶したり置き換えたりするのではなく、進化的視点の追加を通じてこれを是正しようとしています。それは分野により現代的で健全な科学的基盤を提供するはずです。
精神医学は、認知、感情、行動の障害を通じて現れる精神障害を扱う医学の一分野です。しかし、精神障害の病因を理解する上で精神医学が大きな進歩を遂げられなかったことは、主要な進化論者によって「危機」と表現されています(Brüne et al., 2012)- この事実はまた、統合失調症の治療において50年間、またうつ病の治療において20年間、大きな突破口がなかったと述べたScienceの記事でも認められています(Akil et al., 2010)。進化論者は、これは部分的に主流の精神医学が近位的因果関係にのみ焦点を当て、疾患と障害の機械論的説明を好むからだと主張するでしょう。しかし、人間の生理学が正常な機能の明確な参照点を提供する医学とは異なり、精神医学は正常な人間心理学の一貫した理論なしに障害と機能不全を特定しようと試みてきました(Nesse, 2016)。また、主流の精神科医によって機能の重要な問いと進化の役割が考慮される稀な場合でさえ、彼らはそのような思考とアプローチの根本的な転換の完全な意味を探求するところまでには至りません(例えば、Kendler, 2008)。進化精神医学者は、ダーウィン理論が精神医学にとって不可欠な、欠けている基礎科学として役立つと主張しています(Nesse, 2019)。
精神医学の前ダーウィン的状態は、いくつかの精神障害に対する進化モデルの発展と、過去数十年間の影響力のある進化精神医学的文献の出版により、非常に徐々に変化しているかもしれません(Baron-Cohen, 1997; Brüne, 2015; Del Giudice, 2018; Gilbert and Bailey, 2000; McGuire and Troisi, 1998; Nesse, 2019; Stevens and Price, 2000)。
その発展において、進化精神医学は二つの密接に関連する分野からの研究の恩恵を受けています。最初の分野は進化医学であり、これはNesseとWilliams(1994)の基礎的な著作の出版以来、大幅に拡大しました(これに先立つWilliamsとNesse(1991)の論文と1993年の進化医学に関するアメリカ科学振興協会のシンポジウム)、その後多くの研究が続きました(例えばGluckman et al., 2009; Trevathan et al., 2008)。もう一つの分野は、現在高度に成熟し急速に拡大している進化心理学の領域です。これは非常に影響力のある『Adapted Mind』(Barkow et al., 1992)の出版によって学術分野として告知され、その後多くの影響力のある文献や専門学術誌の出版、さらに西洋世界中の多数の大学の学部からの膨大な科学的成果が続きました。さらに、進化人類学者はこれらの学術的な流れ、特に進化医学の発展に大きな影響を与えました(Trevathan et al., 2008)。
この本では、進化が神経生物学的システムの表現型最終産物(これらは選択によって形作られた遺伝的に基づく行動的および心理学的特性です)を決定する上で、精神科医を導くのに理想的に位置づけられている理由を提供しています。重要なことに、機能に対する進化的な強調は、心理的、社会的、文化的領域を包含する生物学的概念の非還元主義的拡張のための科学的基盤を提供することができます(Abed and St John-Smith, 2021)。したがって、精神障害を脳障害(または脳回路障害)として見る主流の生物学的精神医学の狭い「脱文脈化された」見解(Insel and Cuthbert, 2015)とは対照的に、進化論者は環境的文脈が精神障害の存在と性質を決定する上で最も重要であると考えています(Nesse, 2019)。
したがって、進化論者はダーウィン理論を、単に他の多くの視点と並んで考慮されるべき一つの視点ではなく、生命科学全体を支える基本的な組織的枠組みあるいはメタ理論と考えています。進化精神医学は、現代の進化論を精神的健康と疾患の科学的理解に適用したものです。進化精神医学の目標は、人々がどのように病気になるかだけでなく、なぜ病気になるのかを理解することです。
この序章の残りの部分では、人間の健康と疾患の理解に関連する進化科学の基本的事項のいくつかを概観します。¹
1.1.2 進化、自然選択と適応
進化とは何か?進化は、多くの世代にわたる生物や個体群の特徴における正味の方向性のある変化、または累積的な変化と定義することができる—つまり、修正を伴う子孫への伝達である。個体群内の個体が将来の個体群への遺伝的貢献に影響を与える方法で変異する場合、個体群の平均的特性は変化する。
生物学者は進化が起こりうる多くの方法を認識していることを理解することが不可欠であり、自然選択による進化はそのうちの一つに過ぎないが、しばしば最も重要なものと考えられている。他の基本的な進化プロセスには、遺伝的浮動、突然変異、移住、そして性的および社会的選択が含まれる。
自然選択は種分化につながる可能性があり、一つの種が新しく明確に異なる種を生み出す。これは進化を推進するプロセスの一つであり、地球上の生命の多様性を説明するのに役立つ。自然選択は、生物の個体群が適応し変化するプロセスである。しかし、自然選択には予見、計画、目標は含まれない。したがって、個体群内での代替案との競争において繁殖上の利点を与える遺伝的(遺伝子に基づく)表現型形質は広がり、十分な時間が与えられれば、その形質は種全体の特性として固定される可能性がある。繁殖成功度の尺度は「適応度」と呼ばれる。自然選択の繰り返しのサイクルは、成功した変異体の保存と成功度の低い変異体の排除につながり、デザインの出現と生物の適応度を高める形質の形成をもたらす。これらは進化文献では「適応」と呼ばれる。
ダーウィンは遺伝子の存在や変異がどのように生じるかを知らなかったが、現在では変異は突然変異、つまり細胞分裂中のDNA配列のコピーエラーの結果として生じることが分かっている(NIH, 2020)。突然変異が体細胞ではなく生殖系列細胞で発生すると、子孫に伝達される可能性がある。
基本的なダーウィンの考え(変異、遺伝、自然選択)は、20世紀に「現代の総合説」と呼ばれるものによって強化された。これには、遺伝子、突然変異、メンデルの遺伝の概念を含む現代の遺伝学の科学が進化理論に組み込まれた。
現代の総合説は、変異を生み出す主要なメカニズムがランダム(突然変異)である一方で、異なる変異体の成功または失敗はそれらが与える適応度に依存し、まったくランダムではないという洞察につながった。したがって、自然選択は、ある表現型形質を他の形質よりも優先させることによって、生存と繁殖を助ける適応的かつ機能的なシステムを形成し、基礎となる遺伝子が個体群内に広がることにつながる。それにもかかわらず、機能的適応を形成するのと同じ進化プロセスが、逆説的かつ必然的に、不適応(Brady et al., 2019)だけでなく、疾患や障害への脆弱性(Nesse, 2019)を生み出す(ボックス1.1参照)。しかし、疾患と障害の持続の進化的原因に取り組む前に、まず進化的思考がいかに因果関係の理解を変えるかを探求し、その後、他の重要な進化的概念についての簡単な議論を行う。
ボックス1.1 疾患と障害の持続のための進化的経路(Crespi(2016)とGluckman et al.(2009)から改変)
・ミスマッチ
・生活史的要因
・過剰防御メカニズム
・共進化的考慮:病原体との軍拡競争の結果
・進化史によって課せられた制約
・トレードオフ
・性選択とその結果
・バランス選択とヘテロ接合体の優位性
・人口統計学的歴史とその結果
・選択は健康を犠牲にして繁殖成功を好む
・有害なアレル
・適応の極端
1.1.3 ティンバーゲンの因果的枠組み
進化理論の最も重要な意味の一つは、生物科学における因果関係の理解にある。この主題に関する画期的な論文で、ノーベル賞受賞者であり動物行動学の科学の共同創設者であるニコラース・ティンバーゲンは、現在「ティンバーゲンの4つの質問」として知られる因果システムを提案した(Tinbergen, 1963)。Mayr(1961)によって行われた近位(機構的)原因と究極(進化的)原因の区別に基づいて、ティンバーゲンは、あらゆる生物学的システム、形質、または器官の完全な理解には、その因果関係の4つのカテゴリーすべての理解が必要であると提案した(表1.1)。これらは、システムを機能させるメカニズム(生理学、構造)、生物の生涯にわたってシステムを形成する発達プロセス、システムの系統発生史、そして自然環境でそのシステムが生物に提供した機能である。表1.1では、ボックス(1)と(2)がMayrの分類による近位原因に対応し、ボックス(3)と(4)が究極原因に対応する。すべての4つの原因はすべての生物学的現象に同時に適用され、互いに代替するものではなく、これら4つの因果要素のいずれかを無視すると必然的に当該システムまたは形質の不完全な理解につながることに注意することが重要である。
- 表1.1 ティンバーゲンの四つの質問(Nesse, 2013より改変)
- 1.1.4 ダーウィン適応度と包括的適応度
- 1.1.5 進化と心理的メカニズムの概念
- 1.1.6 親の投資理論と親子間葛藤
- 1.1.7 表現型可塑性、カナライゼーション(発達的安定性)および差異的感受性
- 1.1.8 祖先環境または進化的適応環境
- 1.2 疾患と障害の持続のための進化的経路
- 1.2.2 ライフヒストリー理論
- 1.2.3 過剰に活性化した防御機構
- 1.2.4 共進化的考察(病原体と宿主の間の軍拡競争)
- 1.2.5 進化の歴史から生じる制約
- 1.2.6 トレードオフ
- 1.2.7 性選択とその結果
- 1.2.8 均衡選択とヘテロ接合体優位性
- 1.2.9 人口統計学的歴史とその結果
- 1.2.10 選択は健康よりも繁殖成功を優先する
- 1.2.11 有害対立遺伝子
- 1.2.12 機能的適応の極端な末端
- 1.3 進化、人間の独自性と文化の役割
表1.1 ティンバーゲンの四つの質問(Nesse, 2013より改変)
| 発達的/歴史的 | 形質/システムの特性 | |
|---|---|---|
| 近位的原因 | (2) 個体発生:その形質は生物の一生の間にどのように発達するか? | (1) メカニズム:それはどのように機能するか? |
| 進化的または究極的原因 | (3) 系統発生:その形質の系統発生的歴史は何か?(なぜその形質/システムはそのようになっているのか?) | (4) 適応的機能:その形質またはシステムは、自然環境における生物の包括的適応度にどのように貢献してきたか?(なぜその形質/システムは存在するのか?) |
疾患や障害は生物学的システムに影響を与える現象であるため、より伝統的な「どのように」という質問を補足する「なぜ」という質問をすることで、ティンバーゲンのシステムの適用から明らかに恩恵を受けるはずである(いくつかの臨床応用の詳細な議論については第2章を参照)。
近位的原因のみに焦点を当てること(現在の主流の精神医学ではそうである)は、機械に対する技術者の見方に似ており、一方、究極的原因も考慮することはエンジニアの見方に近い(Nesse, 2019)。進化論者は、苦痛を伴う感情状態の認識に熟練し、なぜ私たちがそのような感情を持っているのか、そして感情システムがどのように人々の現在の生活と相互作用するのかを理解している臨床医は、患者の苦痛をより深く理解し、患者の現状に寄与している可能性のある状況をより考慮できる可能性が高いと考えている(Abed and St John-Smith, 2021)。さらに、重要なことに、進化的考察は、精神病理を引き起こしているシステムの正常な機能が何であるかに関する仮説を検証することを通じて研究課題に影響を与える可能性がある;これらは主流の精神医学ではほとんど尋ねられない質問である(Brüne, 2015)。
1.1.4 ダーウィン適応度と包括的適応度
適応度は進化理論における中心的な概念である。ダーウィン適応度は繁殖成功の尺度であり、特定の環境における遺伝子型または表現型に関して定義することができる。これは、指定された遺伝子型または表現型の個体によって次世代の遺伝子プールに対して行われる平均的な貢献によって測定される。適応度が特定の遺伝子の様々なアレル間の違いに影響される場合、それらのアレルの相対的頻度は選択を通じて世代を超えて変化し、個体の適応度にとってより大きな正の効果を持つアレルは時間とともにより一般的になる。
前述したように、現代の遺伝学とダーウィン理論の統合は「現代の総合説」と「包括的適応度」の概念の定式化につながった(Hamilton, 1964)。ハミルトンの定式化によれば、適応度は自分の遺伝子のコピーを持つ直接的な子孫の数だけでなく、同じ遺伝子のコピーを持つ非子孫の親族の数を通じても測定されるべきである。したがって、親族に対して利他的に振る舞うことは、利他主義者への適応度のコストが、親族への適応度の利得に関係係数を掛けたものよりも低ければ、自分の全体的な適応度または包括的適応度(自分の遺伝子のコピーを持つ子孫および非子孫の親族の合計)を向上させることができる(これはハミルトンの法則としても知られている)。これは利他主義の進化と、競争と協力を生み出す条件の理解のための基礎を提供する(Del Giudice, 2018)。「血縁選択」は、ハミルトンの法則の適用を通じて包括的適応度を高める進化戦略に使用される用語である。
1.1.5 進化と心理的メカニズムの概念
自然選択と性選択は、複雑な機能的メカニズム(適応とも呼ばれる)を生み出すことができる唯一の既知の因果プロセスである。適応は、その進化の期間中に生存と繁殖を促進したため、自然選択を通じて種の特徴として存在するようになった遺伝的特性と定義することができる(Tooby and Cosmides, 1992)。繰り返し発生する適応問題を解決することが、任意の適応の機能である。適応は親から子孫への適応の移行に公理的に必要とされるため、適応にはどんな適応においても遺伝子が存在しなければならない。したがって、進化心理学者/精神科医は、すべての脳神経生物学的メカニズム/システムが特定の環境条件のセット内での長い選択プロセスを通じて形作られたという立場から始める(セクション1.1.8参照)(Buss, 2009)。
心理的メカニズムは、進化の歴史を通じて祖先の人間が直面した生存と繁殖の繰り返し発生する問題を解決するために選択によって形作られた専門的な神経生物学的システムとみなされる(Tooby and Cosmides, 1992)。進化したメカニズムの機能と系統発生の理解は、それらの適応的な出力だけでなく、どのようにそして何故これらのメカニズムが誤作動し、不適応反応につながるか(例えば、新しい環境条件下で;セクション1.2.1参照)についても独自の洞察を提供する。進化した心理的メカニズムの例には:恐怖、愛着、安全、地位、配偶、養育が含まれる(Del Giudice, 2018)。
進化したメカニズムの脱線の例示的な例は、カッコウのヒナが特定の鳥種の生得的な親の給餌メカニズムを悪用する方法である。孵化したカッコウのヒナは、その巨大に開いたくちばしを通じて、別の種(例えばオオヨシキリ)の巣にいるにもかかわらず、(親の)給餌反応を引き起こす超正常刺激を提供し、オオヨシキリが自分の子孫を犠牲にしてカッコウのヒナに餌を与えるよう誘導する(例えば、Tanaka et al., 2011)。同様に、人間の進化した心理的メカニズムは、新しい環境条件にさらされると脱線し、不適応反応を生み出し、一部の個人では精神障害につながる可能性がある(セクション1.2.1参照)。
1.1.6 親の投資理論と親子間葛藤
親の投資とは、親が子孫に対して行う投資であり、それによってその子孫の生存確率が高まるものである。定義上、そのような投資は、他の子孫(現在および将来)に投資する能力によって測定される親へのコストを課す。適応度の構成要素には、既存の子孫の幸福、親の将来の繁殖、そして血縁者への援助を通じた包括的適応度が含まれる(Hamilton, 1964; Trivers, 1972)。親の投資は、雄と雌の両方(両親による世話)、雌のみ(排他的な母親による世話)、または雄のみ(排他的な父親による世話)によって行われる可能性がある。世話は、子孫の生涯のどの段階でも提供される可能性があり、出生前(例えば、鳥類における卵の保護と孵化、哺乳類における胎盤栄養)から出生後(例えば、食物の提供と子孫の保護)までである。
親の投資理論は、平均して、配偶子のサイズ、妊娠、授乳、子育てを含む子孫により多く投資する性は、配偶者を選ぶ際により選択的になり、投資が少ない性は配偶者へのアクセスのためにより多くの同性間競争に従事すると予測する。この理論は、人間を含む動物界全体での性的選択と配偶者の好みにおける性差を説明する上で影響力を持ってきた。Trivers(1974)は親の投資理論を拡張して親子間葛藤を説明した:親の視点と子の視点からの最適な投資の間の葛藤である。
養育におけるさらなる複雑さは親子間葛藤によって生じる。これは受胎の瞬間から始まる可能性のある生物学的プロセスである。有性生殖種においてのみ発生するこの葛藤は、母親(または父親)が彼らの子孫と50%関連している一方で、胎児は自身と100%関連しているという事実に基づいている。これは、親と子孫の両方の立場から見た子孫への最適な親の投資の違いから生じる進化的葛藤を意味するために使用される(Trivers, 1974)。
同様に、各兄弟姉妹はそれらの完全な兄弟姉妹のいずれとも50%しか関連していないため、彼らは親の投資の公平な分け前以上、そして親が喜んで提供するよりも多くを獲得しようとする傾向がある。しかし、親子間葛藤は、包括的適応度に関連するプロセスによって機能的かつ統計的に相殺され、したがって親と子孫間の緊密な遺伝的関係によって制限される。これは、一人の子孫が兄弟姉妹を犠牲にして追加の親の投資を得ることにより、生存する兄弟姉妹の数を減少させ、包括的適応度を低下させる可能性があるためである。これにより、他の条件が同じであれば、親子間葛藤は完全な兄弟姉妹間よりも半兄弟姉妹間でより強くなるという予測につながる。これらの観察とモデルは、児童精神医学に関連する重要な影響を持つ可能性がある(第14章と第15章を参照)。
1.1.7 表現型可塑性、カナライゼーション(発達的安定性)および差異的感受性
可塑性は、個体の生涯内に生じる合理的に予測可能な環境変動への進化的適応であり、個体が異なる環境に自らの表現型を「適合」させることを可能にする。表現型可塑性は、特定の環境的手がかりに応じて発達経路を修正する可能性と、個体が環境条件の変動に応じて自らの表現型状態または活動(例えば、代謝)を変化させる能力を説明する(Garland and Kelly, 2006)。
表現型可塑性は、表現型を変化させることによってダーウィン適応度が増加する場合、進化する可能性がある。しかし、可塑性の適応度上の利益は、可塑的反応のコスト(例えば、新しいタンパク質の合成、アイソザイム変異体の発現比率の調整、[ここで文章が途切れています])のトレードオフによって制限される可能性がある。これはストレス素因モデルによって議論される。
変化を検出するための感覚機構の維持)のコストのトレードオフだけでなく、環境的手がかりの予測可能性と信頼性によっても制限される可能性がある。カナライゼーション(発達的安定性)は可塑性の逆であり、遺伝的および環境的干渉や摂動の両方に抵抗する発達的安定性を指す。カナライゼーションメカニズムは極めて重要であり、生物の形質が頑健性を示し、確実に発達することを保証する。しかし、その欠点は可塑性を制限することである(Haltigan et al., 2021; Waddington, 1942)。
伝統的にストレス素因モデルに従って議論されている心理学的知見の別の解釈は差異的感受性である(Belsky, 1997)。両モデルは、発達が環境の経験や質に対して差異的に感受性を持つ可能性があることを示唆している。ストレス素因モデルは明確でほとんど否定的感受性のある反応を示唆するのに対し、Belskyは逆境体験に敏感であるだけでなく、肯定的体験にも敏感なグループを記述している。一部の個人が二重にあるいは独自に肯定的感受性を持ち、他の個人が独自に否定的感受性を持つ場合、これらのモデルは補完的である可能性がある。
Bakermans-KranenburgとIJzendoorn(2006)は、遺伝的要因の関数として差異的感受性仮説を検証した最初の研究者であり、47家族における母親の感受性と外在化行動問題の関連におけるドーパミン受容体D4 7回反復多型(DRD4-7R)の調整効果を検討した。DRD4-7Rアレルを持ち「非感受性の母親」を持つ子どもは、同じアレルを持つが「感受性の」母親を持つ子どもよりも有意に多くの外在化行動を示した。DRD4-7Rアレルを持ち感受性の母親を持つ子どもは全体の中で最も外在化行動が少なく、一方、母親の感受性はDRD4-7Rアレルを持たない子どもには影響を与えなかった。
研究はまた、セロトニントランスポーター遺伝子HTTPLRの少なくとも1つのsアレルを持つことが、逆境に直面した際にうつ病を発症するリスクを高めることを実証している。しかし、同じ変異はいくつかの領域における優れた認知パフォーマンスと関連し、社会的同調性を高める(Homberg and Lesch, 2011)。
これらの例は単純な遺伝子決定論に対する証拠として役立ち、また単純に障害を治療するために遺伝子だけを変更することを目指すことは、状況が異なれば、そのような遺伝子の害や利益が変わるため、個人の最善の利益にならない可能性があることの兆候を提供する。
1.1.8 祖先環境または進化的適応環境
進化的適応環境(EEA)の概念は、愛着理論で有名なジョン・ボウルビー(1969)によって最初に提案された。広く言えば、EEAは現代人の特徴的な形質が形成された全体的な祖先人類環境を指す。それはしばしば単一の均一な時間と場所であるかのように誤って言及される。しかし、それはより適切に「祖先の個体群のメンバーが遭遇した祖先環境の適応関連特性の統計的複合体であり、その頻度と適応度結果によって重み付けされた」(Tooby and Cosmides, 1990: 386-387)と概念化される。したがって、EEAは特定の時間枠にわたる外部条件や刺激、脅威と機会、栄養素、社会的圧力、寄生虫や捕食者や競争相手からの脅威だけでなく気候や一般的な生息地への個体群の曝露の総和を表す複合的な概念である(Gluckman et al., 2009)。したがって、それは「種の最近の部分の環境特性の複合体であり、その現代の適応コレクションが現在の形を取った期間を包含する進化」(Tooby and Cosmides, 1990: 388)と考えることができる。「異なる適応は異なるEEAを持つことに注意することが重要である。言語のような一部は約200万年前にしっかりと固定されているが、幼児の愛着などの他のものははるかに長い進化史を反映している」(Durrant and Ellis, 2003: 10)。
EEAの概念の批評家たちは、私たちは遠い祖先がどのように生きていたかについてあまり知らないと主張し、これによりEEAの概念は非常に投機的で非科学的な前提になると主張している(Hagen, 2016)。Gould(1997)、Buller(2005)、LalandとBrown(2011)などの批評家たちもEEAの概念の使用に反対した。なぜなら、彼らは私たちが祖先の生活条件を十分な精度で特定することができないと仮定したからである。これらの懸念のいくつかは正当であり、真剣に考慮されるべきであるという疑いはない。しかし、もし彼らの主張が真実であり、私たちが祖先がどのように生きていたかについて何も知ることができず、将来もできないとすれば、進化的アプローチはどんな身体的または脳のシステムの正確な機能も確認できないだろう。すべての機能は過去の環境に対応して選択によって形作られた適応であるため、過去の環境に関する事実を発見することは進化的努力の重要な部分であり、現在の機能と機能不全を理解するための前提条件である。
私たちは過去についてあまり知ることができないという主張は現在もはや持続可能ではなく、過去の調査に完全に焦点を当てている幅広い学問分野と矛盾している。これらの分野には考古学、古生物学、古人類学、歴史学、宇宙論が含まれ、現在では化石や人工物の研究だけでなく、古代および絶滅種のDNA配列も含まれている(Hagen, 2020)。これにより大きな進歩が可能になり、過去に関する事実を発見することを目的とした科学的研究が、過去の環境の厳密で検証可能で反証可能なモデルを生み出すことができることを明確かつ決定的に示している(例えば、Dunbar, 2014)。過去の知識がなければ、進化科学は単に進歩することができず、したがってEEAまたは祖先人類環境の概念は不可欠である。これは人間の進化史に関する声明が盲目的または無批判に受け入れられるべきであることを意味するわけではない。そのようなすべての主張は、証拠によって支持または反証される可能性のある仮説として述べられるべきであり、同様のレベルの科学的厳密さも人間の心理的適応とその機能に関する仮説に適用されるべきである。
1.2 疾患と障害の持続のための進化的経路
前述のように、自然選択は何千世代にもわたって繁殖成功(適応度)を高めるように形作られた適応の詰め合わせを持つ身体と脳を生み出すが、必ずしも幸福や快適さを高めるわけではない。なぜ進化が人間を疾患や障害に対してこれほど脆弱にしたのかという難問の説明は、現代の進化医学の創始者たちによって最初に提起されて以来(Nesse and Williams, 1994)、それ自体が進化してきた。それに応じて、進化的プロセスが疾患や障害の存在と持続につながる可能性のある一連の経路が提案されており、ボックス1.1に示されている。
これらの経路のいくつかは精神医学にとって他よりも関連性が高く、それらは相互排他的ではない。いくつかは精神障害の起源において同時にまたは連続的に関与している可能性がある。それらは、精神障害を含む、疾患や障害に対する私たちの脆弱性の究極的/進化的原因のリストを表している。
1.2.1 ミスマッチ
ミスマッチは間違いなく進化医学の最も重要な洞察の一つであり、冠状動脈疾患、高血圧、肥満、2型糖尿病、うつ病、アルコール依存症、摂食障害など、現代環境で蔓延している一連の疾患や障害の理解には不可欠である(Nesse and Williams, 1994; Pollard, 2008)。ミスマッチの考え方は、適応が特定の環境内での選択によって形作られるという事実に基づいている。環境が急速かつ根本的に変化すると、一部の生物学的システムは新しい環境に不適合になるリスクを負う。これは「ゲノムの遅れ」とも呼ばれる(Li et al., 2018)。現代の人間環境が祖先の環境からいくつかの点で根本的な変化を遂げたことを考えると、これによりいくつかのシステムがこの新しい環境に不適合になり、一連の精神障害を含む機能不全の結果をもたらした(詳細な議論については第2章を参照)。現代環境で発生/増加している精神障害の例には、摂食障害(Rantala, 2019; Russell, 2000)および薬物やアルコール依存症(Nesse, 2005)が含まれる(栄養のミスマッチの図については図1.1を参照)。しかし、人間は現代環境の特定の側面(例えば、常に豊富な栄養素、特に超加工食品)に不適合かもしれないが、私たちは現代の条件の大部分によく適合している。人間は明らかに繁栄しており、絶滅していないからである(Hagen, 2020)。
現代環境における栄養的ミスマッチの図解
| (1) 祖先の環境 | (2) 現代の環境 | |
|---|---|---|
| 進化したエネルギー獲得システムのアルゴリズム | 「見つけられる最も甘い食べ物を食べる」 | 「見つけられる最も甘い食べ物を食べる」 |
| 利用可能な食品 | 単純糖は希少で非常に価値が高い<br>(適応的インプット) | 超加工食品:<br>単純糖が普遍的に存在し、高濃度で含まれている<br>(ミスマッチしたインプット) |
| 結果 | 適応的結果:<br>熟した果物、ベリー類、ヤム芋、蜂蜜など、<br>カロリー、ビタミン、食物繊維を提供 | 不適応的結果:<br>高糖度の清涼飲料水、菓子類、ペストリー、アイスクリーム、<br>過剰なカロリーだが栄養価が低い |
図1.1 現代環境における栄養的ミスマッチの図解(Li et al., 2018より改変)
1.2.2 ライフヒストリー理論
ライフヒストリー理論(LHT)は、個体の生涯にわたって変化する生存と繁殖に関連する問題に対する種の典型的な解決策を扱います(Brüne, 2015)。したがって、LHTは生物が生涯を通じて中心的な生物社会的目標を達成するために時間とエネルギーをどのように配分するかを理解するための枠組みを提供します。ライフヒストリー戦略には、性的成熟の時期や子孫の数と質、さらには寿命の長さなどの重要な生物学的発達を形作る一連のトレードオフが含まれます(Stearns, 1992)。LHTの適用により、これらのトレードオフがライフヒストリー戦略のスペクトルを生み出すことが示されており、そのトレードオフには体細胞対繁殖努力、現在対未来、子孫の質対量が含まれます。スペクトルの「速い」端は、より短い寿命、より速い成長、より早い成熟と繁殖、より多くの子孫によって特徴づけられ、一方でライフヒストリースペクトルの「遅い」端にいる生物は反対の特徴を示します(Del Giudice, 2018)。ライフヒストリーの速い-遅いスペクトルという考え方は、精神障害への脆弱性を含む個人差を理解するための枠組みとして提案されています(Del Giudice, 2018)。ライフヒストリー戦略の違いは部分的に遺伝的制御下にありますが、個人の初期環境の性質と質も重要であるように思われます(Belsky et al., 1991; Ellis et al., 2011)。種間ではなく個人間の特性変動へのLHTの適用は、最近批判的精査の対象となっています(例:Zietsch and Sidari, 2020)。その結果、この研究分野は方法論と理論的前提の両方について相当な改訂が進行中です(Del Giudice, 2020; Young et al., 2020)。
1.2.3 過剰に活性化した防御機構
気分や不安を調節するシステムなどの防御機構は過剰に活性化したり調節不全に陥ったりすることがあり、有害な結果をもたらし、防御活性化障害につながることがあります(Del Giudice, 2018; Nesse, 2019)。一般医学における防御の例には、痛み、下痢、嘔吐、発熱があり、同様の原則が適用されます。重要なことに、生物学的に進化したシステムであれ人工システムであれ、すべての防御機構には、誤警報(偽陽性とも呼ばれる)を許容するという共通の設計特性があります。これはリスクが存在するにもかかわらず活性化しない(偽陰性、通常は壊滅的な結果をもたらす;例えば、毒物を摂取しても嘔吐しない場合を想像してください)よりもはるかにコストが低いためです。これは「火災報知器の原則」と呼ばれ、すべての身体防御機構(嫌悪感情を含む)が過剰に活性化する可能性がある理由を説明します(Nesse, 2019)。すべての防御システムを特徴づける過剰な誤警報の傾向は、「用心するに越したことはない」という戦略に似ており(Blumstein, 2020)、反応が不必要あるいは逆効果であると確立された場合、防御を安全にブロックできる理由を説明します。
1.2.4 共進化的考察(病原体と宿主の間の軍拡競争)
宿主としての人間は、細菌やウイルスなどの急速に進化する病原体との終わりのない軍拡競争に従事してきており、今後も続けていきます(Ewald, 1994)。これは、ますます革新的な宿主防御(例えば、ますます洗練された免疫応答)が、さらに新しい防御回避方法と拮抗することを意味します。また、抗菌療法に耐性を持つ病原体が増えており、これは人間の健康にとってますます深刻な危険をもたらしています。この軍拡競争において、病原体は急速な複製者として、はるかに速い進化能力のために有利です(Nesse, 2005)。最近のCovid-19パンデミックは、新たに進化したウイルスが種を越えて人間集団に世界的に広がり、人命と生活に大きな犠牲を払った生々しい例であり、将来的にも同様のパンデミックが他にも発生することは間違いないようです。これは一般医学にとって間違いなく大きな問題ですが、メンタルヘルスにおける例はより限定的に見えます。例えば、強迫性障害(OCD)は連鎖球菌誘発性自己免疫疾患の結果として生じることがあり(Swedo et al., 1994)、トキソプラズマ・ゴンディと統合失調症との関連の示唆が継続的になされています(Fuglewicz et al., 2017)(第10章参照)。
1.2.5 進化の歴史から生じる制約
人間の設計者とは異なり、進化は図面に戻って一から始めることはできません。これは「経路依存性」と呼ばれ、人間の目が網膜の一部を覆う血管を持つ理由(Nesse, 2005)、人間の網膜の光受容体が間違った方向を向いている理由(Lents, 2020)、反回神経の曲がりくねった経路、そして四足歩行の設計計画を修正した二足歩行の骨格が、普遍的な背部問題から胎児の頭を通せないほど狭い産道までの無数の脆弱性を生み出す理由を説明します(Pavličev et al., 2020; Taylor, 2015)。これがまた、系統発生学的歴史と進化的適応環境(EEA)がとても重要である理由です。進化はこれまでのものを使って作業しなければなりません;複雑なシステムは無から作られるわけではありません。進化は工夫家として描写され、生存と繁殖にちょうど十分に機能する適応(利用可能な生物学的システムから)を形作ります(Jacobs, 1977)。したがって、進化は歴史的制約、複数のトレードオフ、および(遺伝的)エラーを通じて適応を形成するプロセスです(Nesse, 2005)。進化的思考は、したがって、生物学的システムに普遍的に見られる欠陥、癖、曲がりくねった複雑さを説明し、これらはすべて機能不全や障害への脆弱性を生み出す可能性があります。
1.2.6 トレードオフ
生物学的に進化したすべての適応、特性、システムはトレードオフを表していることを理解する必要があります。ある特性を向上させることは、しばしば別の特性のパフォーマンスを悪化させる犠牲を伴うからです。例えば、感染への抵抗力を高めると自己免疫疾患のリスクが高まります。また、これはエネルギー保存と飢餓抵抗力を向上させることが、食物が豊富になったときに肥満のリスクを高める理由を説明し、体のサイズ/筋肉量と動きの速さの間の相互トレードオフには最適なバランスがあり、一方を増やすともう一方が減少する可能性があることを説明します。
1.2.7 性選択とその結果
性選択はダーウィン(1871)によって、生存に役立たず、さらには生存に不利益となる可能性のある特性の進化を説明するために記述されました。性選択された特性の典型的な例は孔雀の尾であり、これは生存目的には役立ちませんが、雌孔雀を引き付けるものです。性選択された特性は、異性への魅力を高めることによって繁殖成功率を向上させるものです。性選択はヒトを含むすべての有性生殖生物に発生し、通常、偽装が困難で、したがって良好な健康状態と高品質の遺伝子の正直なマーカーとして機能する、コストがかかり贅沢な特性の表示を伴います。性選択は性別に分岐する特性を形成し、特に異性の好みをターゲットにする傾向があります。性選択された特性の進化は、しばしば性別比率が偏っている精神障害に対する特定の種類の脆弱性を生み出す可能性があります。性選択が重要な役割を果たす可能性のある精神障害の例には、摂食障害(Abed, 1998)(第11章参照)、性機能障害、統合失調症(Del Giudice, 2017)が含まれます。
1.2.8 均衡選択とヘテロ接合体優位性
ヒトなどの二倍体種では、2つの対立遺伝子は同一(ホモ接合体)または異なる(ヘテロ接合体)場合があります。医学におけるヘテロ接合体優位性の古典的な例は鎌状赤血球貧血であり、ヘテロ接合体状態はマラリアに対する免疫を付与します(マラリアが風土病である世界の一部では大きな利点)が、ホモ接合体状態は深刻で衰弱性の疾患である鎌状赤血球貧血を引き起こします(Gluckman et al., 2009)。この例では、ヘテロ接合体状態の利点はホモ接合体状態の有害な影響によって相殺されます。医学におけるヘテロ接合体優位性の他の例はより推測的です(例:嚢胞性線維症)。現在、メンタルヘルスに関連するこのプロセスの例はありません。
1.2.9 人口統計学的歴史とその結果
人間のアフリカからの移住は約70,000年前以降に行われました。それらは連続的な波で行われ、その過程で人間集団はしばしば障害物またはボトルネック(飢饉、疾病などによる)を通過し、それが遺伝的多様性を著しく減少させました(Henn et al., 2012)。このようなシナリオには、偶然の出来事によって孤立し、小さな孤立したコミュニティで生活を続ける小さな集団も含まれ、そこでは通常は稀な突然変異が「創始者効果」の結果として異常に広まる可能性があります(Gluckman et al., 2009)。このような偶然の出来事は遺伝的浮動とも呼ばれます。創始者効果の結果として稀な有害遺伝子が広まる例には、アシュケナージ・ユダヤ人にのみ影響するテイ・サックス病や、フランス系カナダ人に不釣り合いに多く見られるゴーシェ病が含まれます。興味深いことに、最近の発見は、世界的に見て、アフリカからの移住において人間集団は一連のボトルネック(および創始者効果)の影響を受け、アフリカ大陸から離れるほど増大し、その結果として遺伝的多様性が連続的に減少したことを示唆しています(Henn et al., 2012)。この現象はまた、ベネズエラ、コロンビア、ペルー、ブラジルでのハンチントン病の高い有病率を説明するかもしれません(Kay et al., 2017)。
1.2.10 選択は健康よりも繁殖成功を優先する
ダーウィン理論の基本的な原則は、選択が必ずしも良好な健康と幸福を通じてではなく、繁殖成功を通じて働くということです。したがって、健康と幸福を減少させるが繁殖成功を高める遺伝子は、それにもかかわらず集団内で広がるでしょう(Nesse, 2005)。したがって、高レベルの競争力、協力性の低下、嫉妬心・貪欲さ・羨望の増加、そして鎮めることのできない性的欲求は、自己と他者の健康と幸福に対する潜在的な悪影響にもかかわらず、繁殖成功に対するそれらの肯定的な効果のために広がるでしょう(Buss, 2000)。
1.2.11 有害対立遺伝子
生殖年齢を超えて生存を可能にし、晩年まで現れない有害遺伝子は、ハンチントン病の原因となるものなど、集団内に残る可能性があります。そのような対立遺伝子/突然変異は、選択がそれらを排除する能力が限られているため、集団内で低レベルで残ることがあります。また、致命的でない新生突然変異(親の生涯中に生殖細胞系列で発生し、親はその突然変異の影響を受けない突然変異)は子孫に伝達されます。そのような突然変異は最初の世代では選択に見えず、生殖年齢までの生存と両立する場合にのみ、後続の世代で選択の対象となります。
1.2.12 機能的適応の極端な末端
不安、恐怖、潔癖症などの機能的に適応的な特性は、スペクトルの極端な末端では機能不全になり、不適応になることがあり、そこでは全般性不安障害、恐怖症、強迫性障害(OCD)として現れます(Abed and De Pauw, 1998; Crespi, 2016)。同様に、適応的な性格特性の極端な末端は機能不全で不適応な状態につながる可能性があります(Trull and Widiger, 2013)。不適応な極端は両端で機能不全になることがあります(つまり、特性が低すぎる場合も高すぎる場合も)。これは、心理化(心の理論)のような生物学的に基づく特性にも当てはまり、過活動と活動不足の両方が精神障害(それぞれ統合失調症と自閉症スペクトラム障害)に関与していることが示されています(Crespi, 2016)。
したがって、進化的視点を取ることで、精神的苦痛が機能的システム(例えば過剰に活性化した防御機構やミスマッチ)から生じうるという重要な洞察が得られます(Abed et al., 2019)。したがって、医療専門家による介入を依然として正当化すべき望ましくない状態は、真の生物学的機能不全を必ずしも伴わない様々な状況から生じる可能性があります。さらに、進化的分析は、機能的または機能不全のシステムから生じる精神的苦痛と精神障害の状態を区別することを可能にする理論的枠組みを提供し、また環境的文脈の役割をより効果的に理解する方法を提供します(有害な機能不全の議論については第2章を参照)。
1.3 進化、人間の独自性と文化の役割
1.3.1 社会的および文化的脳
人間の脳は体重の2%を占めますが、体が必要とする総エネルギーの15-20%を消費します(Brüne, 2015)。このような印象的な事実は進化的説明を必要とします。なぜなら、そのようにエネルギー的に高価な器官は、その利益がコストを上回る場合にのみ進化することができるからです(Aiello and Wheeler, 1995)。これに対する最も説得力があり実証的に支持されている説明は、社会的環境の要求が人間の進化史の過程での脳サイズの増加の主な推進力であったということです(Dunbar, 2003a; Humphrey, 1976)。
社会的脳とは、社会的領域に関連する情報の処理を専門とする神経生物学的システムのネットワークを指します(Brothers, 1997; Dunbar, 2003b)。種の脳のサイズとその社会集団の平均サイズの間には、種を超えて当てはまる定量的な関係があるようです(Dunbar, 2014)。しかし、脳のサイズと集団のサイズの関係(厳密に言えば新皮質の体積)は、結合した社会的関係を持ち、高度に構造化された集団を形成する種にのみ適用されます。これらの社会的絆は友情の形で現れます—ペアの絆に似た、感情的に密接な関係ですが、性と生殖を伴いません。このような複雑な社会的配置は、持続的な絆を形成しない群れ動物の一時的で短命な相互作用とは鋭く対照的です(Dunbar, 2014)。
狩猟採集民の社会組織は、30-50人の集団、150人の結合したコミュニティ、500人の内婚的コミュニティ、そして1,500人の民族言語的単位(部族)から始まる入れ子状の階層的集団によって特徴づけられていたようです(Dunbar, 2014)。興味深いことに、これらの様々なレベルの組織は、現代の人間の「知人の同心円」にもまだ明らかです(Dunbar, 2014)。
このような社会的複雑性の増加は、最も特徴的な人間の特性の一つである言語なしには不可能だったでしょう(Del Giudice, 2018; Pinker, 1994)。これにより、累積的文化の基礎を築いた前例のない情報と知識の移転レベルが可能になりました(セクション1.3.2参照)。何百万人(現代の国家では数億人にも)から成るメガグループへの人間の社会組織の拡大は、大規模な文化獲得と伝達のための人間の能力なしには不可能だったでしょう(Henrich, 2016)。社会的脳の機能と機能不全を理解することは、統合失調症(Burns, 2007)や自閉症スペクトラム障害(Baron-Cohen, 1994)を含む、社会性の障害を伴ういくつかの精神障害に関連する可能性があります。
1.3.2 文化的進化と遺伝子-文化共進化
人間は、他の生物とは異なり、平行して相互作用する2つの継承システム、つまり遺伝子と文化を持っています。ある種の文化は広範囲の種に存在し、人間に固有のものではありませんが、累積的文化進化、つまり一人の個人だけでは達成できない多くの世代にわたる累積的改善(時に文化的ラチェット効果と呼ばれる;Tomasello, 1990)は、広く我々の種に固有のものと考えられています(Dean et al., 2014)。累積的文化進化は、遺伝子に選択圧をもたらす環境の変化を生み出します。さらに、文化的に進化した社会環境は、そのような環境に適した継承された心理を持つことを好みます(Richerson and Boyd, 2005)。現代人における遺伝子-文化共進化の結果は、一部の著者によって記述されている「自己家畜化」の現象に関連しています(例:Brüne, 2007)。並行して実行されているこれら2つの進化し相互作用するシステム(遺伝子と文化)の存在は、「二重継承システム」と呼ばれています。
行動的にも形態的にも人間を形作る上での文化の役割は過大評価できません。例えば、チンパンジーと人間の最後の共通祖先(LCA)(約700万年前に生存していた)がどのように見えたかは分かりませんが、影響力のある見解ではLCAはチンパンジーのような類人猿だったと示唆しています(Pilbeam and Lieberman, 2017)。もしそうであれば、人間はLCA以来真に急進的な行動的・形態的変容を遂げたのに対し、チンパンジーは同じ期間中にほとんど変化していないように見えます。同様に、累積的文化や遺伝子-文化共進化がなければ、ゴリラ(形態的に過成長したチンパンジーに似ています)はさらに長い進化的時間スケールでも著しく変化していません(人間はゴリラと約1000万年前に共通祖先を共有していました)。この一つの妥当な説明は、人間が持続的かつ長期的な遺伝子-文化共進化の影響を受けてきたのに対し、チンパンジー(ボノボを含む)系統や他の大型類人猿の系統にはそのような影響の証拠がないことです(Henrich and Tennie, 2017)。したがって、遺伝子-文化共進化が人間の心、人間の身体、そして人間の社会構造を形作ってきたと示唆することは過言ではないかもしれません。最終的に、文化自体が環境であり、したがって選択プロセスの一部となるため、文化がどのように遺伝子頻度に影響を与えることができ、実際に影響を与えてきたかを考慮する価値があります。
進化的時間スケールにわたる遺伝子選択に対する文化の影響に加えて、文化的実践が個人の一生の間に私たちの脳の再配線をもたらすことができるように思われます(Henrich, 2020)。広範囲に研究されてきた例は読み書き能力であり、読み書きができる人は読み書きができない人とは有意に異なる脳を持っており、より厚い脳梁、顔の処理を右半球へのシフト、顔認識の障害、そして言語記憶の改善が示されています(Dehaene et al., 2015)。
現代人における遺伝子-文化共進化の影響の顕著な例は、消化システムの形成です。私たちの消化システムは、小さな口、小さな歯、小さな胃、そして短い結腸を持つ私たちのサイズの生物にとっては異常であり、これらはすべて人間の系統における食物準備の文化的習慣と料理における火の使用の結果として進化してきました(Wrangham, 2009)。また、文化的に影響を受けた、人間の進化的時間スケールにわたる狩猟や戦闘における投射物を正確に使用するユニークな人間の能力は、もはや近接戦闘で狩猟や戦闘をする必要がなくなった現代人の身体的頑健性の減少の説明として提案されています(Richerson and Boyd, 2005)。遺伝子-文化共進化のより進化的に最近の結果には、乳製品文化を持つ祖先の子孫である集団(主に北ヨーロッパ、中東の一部、アフリカの一部)に発生するラクターゼ持続性(乳糖耐性)の進化が含まれます。これらの集団は、6,000-7,000年前に発生した突然変異を持ち、成人が乳糖を消化することを可能にします(Cavali-Sforza et al., 1994)。文化はまた人間の心理を形作り、規範違反者を排除し、社交的で従順な従順者に報酬を与えることにより、私たちを「文化中毒者」にしました。人間はまた、私たちの内集団のメンバーから正確に学習する能力を進化させ、「集団脳」と呼ばれるものを発達させました(Henrich, 2016)。
人間の進化史にわたる生態学的、社会的、文化的要因を分離しようとする試みで、代謝的アプローチを取った最近の研究は、社会的課題ではなく生態学的課題が重要な役割を果たし、人間の系統における非凡な脳の成長は文化によって強く促進されたことを示唆しています(Gonzalez-Forero and Gardner, 2018)。この発見は人間の脳進化の文化的仮説と一致しています(Henrich, 2016; Laland, 2017)。
興味深いことに、最近、人間の進化に関する新しい理論が提案され、累積的文化進化というユニークな人間現象に直接関係する可能性があります。これは、70,000年から100,000年前の間に現代人が「システム化メカニズム」を進化させ、これによって人間が他の種には隠されている世界のパターンを検出することができるようになったという提案です(Baron-Cohen, 2020)。この主張は、このメカニズムが人間に「生成的創造性」という才能を独自に付与し、約50,000年前以降の考古学的記録に見られる文化的/芸術的/実用的工芸品の非凡な多様化をもたらし、それ以前の長期間の文化的停滞と鋭く対照をなしていると述べられています(Baron-Cohen, 2020)。システム化メカニズムの極端な変種は、自閉症スペクトラム障害を持つ個人に明らかであると主張されています。この理論は累積的文化進化の現象に明確な含意を持っています。なぜなら、システム化メカニズムは人間に存在し他の種には存在しない新しい発明(文化的変種)の連続的な生産のためのエンジンを提供するからです。
しかし、Baron-Cohenが最近提案した自閉症スペクトラム障害とシステム化メカニズムの間の関連を除いて、最近の文化進化モデルと遺伝子-文化共進化、および文化的実践の発達的影響に関する最近の研究結果のメンタルヘルスへの影響は、大部分が未探索のままであり、進化心理学者と精神科医からのさらなる注目に値します。
1.3.3 精神障害と自然選択圧リラクゼーションモデル
最近発表された一つの進化論は、人間における精神障害の出現が自然選択圧のリラクゼーション(RNSP)の結果であると説明しようとします(Fuchs, 2019)。これは、人間が何千世代もの進化史の間に、彼らの周りに広範囲の危険(例えば捕食)から彼らを守る環境を構築してきた結果、著者が「人間の本能的衝動の多様化」と呼んでいるものをもたらしたと提案する興味深く斬新なアイデアです。著者は、本能的衝動の分化/拡散、開放的および閉鎖的遺伝的本能プログラム(Mayr, 1974)、能動的対反応的行動、および本能的衝動の欲求不満の結果として空虚な行動、置換、攻撃性、不快感を含む、一連の健全な進化的および動物行動学的アイデアと概念に彼の仮定を基づけています。保護環境の人間による構築に関する彼のアイデアは、「ニッチ構築」と遺伝子-文化共進化の両方の概念に密接に関連しています。興味深いことに、Laland et al.(2017)はニッチ構築の議論において、この現象を自然選択と人工選択の間のどこかに位置づけ、ニッチ構築は遺伝的多様性を減少させるが人工選択ほどではないと考えました。この見解はFuchsのRNSPモデルと矛盾するように思われます。
それにもかかわらず、特定の自然選択圧のリラクゼーションは観察可能な現象であり、特定の選択圧から解放された特性に明確な結果をもたらします。これは洞窟魚(Calderoni et al., 2016)で実証されており、彼らは視力を失うだけでなく(完全な暗闇では視力がないことは不利ではなく、視力の維持にはコストがかかるため、不完全な構造はもはや排除されません)、昼光の欠如は彼らの生物時計の劣化にもつながります。RNSPの効果は人間(および多くの他の霊長類)でも明らかであり、植物源からのビタミンCの広範な利用可能性により、GULO遺伝子の機能喪失が、ビタミンCを合成する能力の喪失(大多数の哺乳類とは異なり)をもたらし、したがって壊血病のリスクに対する人間の感受性を生じさせました(Lents, 2020)。
Fuchsは、RNSPが人間の進化の間に有用な人間の適応を形作るために利用されたが、これらの多様化した適応の極端が機能不全の行動形態と様々な形の精神障害につながったと主張しています。精神障害の理解のためのFuchsの斬新な理論の長所と含意は大部分が未探索のままであり、進化論者からのさらなる注目に値します。
現在の文化、特に医学にFuchsのRNSPモデルを適用する際、注意すべきは、捕食者や多くの感染症のリスクなど一部の領域では選択圧が緩和されている一方で、他の領域では新しい選択圧が生じているということです。もちろん、人間は微生物(古いものも新しいものも)などの古い脅威に対して依然として脆弱であり、現代生活やテクノロジーのストレスなどの新しい圧力が、現在のところ不明な適応度への影響、ひいては将来の世代の人間ゲノムの構成に影響を与えています。したがって、選択圧は異なるのであって、存在しないわけではありません。
1.4 意義と結論
1.4.1 機会
本章では、進化科学が精神医学にもたらす様々な利点について論じてきました。これらの利点は以下のようにまとめることができます(Nesse, 2005):
(1) 進化がなぜ私たち全員を精神疾患に対して脆弱にしたのかについて、新たな問いを投げかけること;
(2) 発達と、幼少期の経験がその後の特性にどのように影響するかについて明確に考える方法を提供すること;
(3) 感情とその調節を理解するための基盤を提供すること;
(4) 科学的診断システムの基盤を提供すること;
(5) なぜある人々は精神疾患になり、他の人々はならないのかを説明する複数の因果要因を組み込むための枠組みを提供すること。
しかし、進化精神医学は主流の精神医学に取って代わることを目指しているわけではないことを強調することが重要です。それは主流の精神医学的思考を補完し、情報を提供し、拡張するものです。進化論者は、個々の患者の利益、彼らの福祉、そして患者や他者への害の軽減が中心的関心事である現在の精神医学の実践を支配する倫理原則を完全に受け入れています。さらに、進化精神医学者は証拠に基づく医療の原則に全面的に賛同しており、純粋に理論的な定式化に基づいて未検証の治療法を患者に提案したり処方したりすることはありません。進化的思考は、新しい治療法の提案につながる可能性のある因果関係に関する理論を生み出すことができます。しかし、そのような治療法は、現在の主流医学および精神医学で使用されている標準的な科学的方法論を用いて同じ厳格な評価を受けるべきです。進化精神医学は、社会的ダーウィニズムや優生学のイデオロギー的で非科学的な教義とは関連がありません(Wilson, 2019参照)。したがって、進化精神医学は、進化過程から生じる人間の脆弱性についての理解を活用して、あらゆる倫理的かつ証拠に基づいた方法で個々の患者の苦痛を軽減し、回復を支援することを目指しています(Troisi, 2015)。
1.4.2 臨床的有用性
現在、進化理論に基づく心理学的/精神医学的介入の例はほんの数例しかありません。これには、思いやり焦点療法(Gilbert, 2020)や新しいタイプの認知行動療法(Abrams, 2020)が含まれます。それにもかかわらず、進化的知識は、特定の療法を実施していない場合でも、臨床実践において患者と臨床医の両方にとって有用であると私たちは主張します(例えば、共感の強化、文脈へのより大きな注意、感情の進化した機能についてのより深い理解を通じて)。しかし、進化の主な有用性は、個人よりも集団における健康と疾病を理解する手助けをすることにあります。したがって、進化科学は治療学よりも疫学に類似しています。他のすべての健康と疾病のモデルと同様に、進化理論がすべての未解決の問題を即座に解決するわけではないことを理解することが重要です。精神医学における問題は特に複雑であるため、特定のアプローチの有効性に関する広範な主張は単に信頼できません。しかし、進化理論はすでに生物学の複数の領域で非常に価値があることが証明されています。人間のゲノムのより深い理解が医学を個別化された治療へと導くにつれて、進化的アプローチは貴重な洞察を提供することができます。例えば、進化は私たちに、正常なゲノムというものは存在しないことを思い出させます。存在するのは、特定の環境において高いまたは低い生殖的成功をもたらす表現型を構築する遺伝子だけです(Nesse and Dawkins, 2010)。
1.4.3 研究への影響
進化的アプローチは、疾患の病因に関する新しい種類の問いを提起します。これらの問いに答えるための研究により、最終的には精神医学文献が各疾患に対する進化的考察を提供できるようになるはずです(Nesse and Dawkins, 2010)。そのような問いや仮説を定式化するための戦略はまだ確立されておらず、進化仮説を検証するための方法は医学の多くの人々にとって馴染みがありません。Nesse(2011)は、最近の例を用いて成功した戦略と一般的な課題の両方を説明する、適切な進化研究の構造を提案しています。彼は進化仮説を検証する際に考慮すべき適切な問いを特定しています。これらに体系的に取り組むことで、混乱や誤りを最小限に抑えることができます。
1.4.4 結論
進化の初心者と進化的学者の両方に対応することを望む進化精神医学の巻への導入を提供することが課題になることを私たちは認識しています。進化理論の基本をカバーしながら、含まれるテーマに関する現在の最新の考え方を提示することを目指しました。特に、進化に初めて触れる人々がこの導入を読んだ結果、本巻の残りの部分に取り組むための準備がより整い、また本章の参考文献を参照したり、進化心理学および精神医学に関する豊かで魅力的な文献についてより広く読んだりすることに興味を持つことを願っています。
References
Abed, R. (1998). The sexual competition hypothesis for eating disorders. British
Journal of Medical Psychology, 71, 525-547.
Abed, R. and De Pauw, K. W. (1998). An evolutionary hypothesis for obsessive-
compulsive disorder: a psychological immune system? Behavioural Neurology, 11,
245-250.
Abed, R. and St John-Smith, P. (2021). Evolutionary psychology and psychiatry. In
T. K. Shackleford (ed.), The Sage Handbook of Evolutionary Psychology: Applications
of Evolutionary Psychology (pp. 24-50). London: Sage.
Abed, R., Ayton, A., St John-Smith, P., Swanepoel, A. and Tracy, D. (2019).
Evolutionary biology: an essential basic science for the training of the next
generation of psychiatrists. British Journal of Psychiatry, 215, 699-701.
Abrams, M. (2020). The New CBT: Clinical Evolutionary Psychology. San Diego, CA:
Cognella Academic Publishing.
Aiello, L. C. and Wheeler, P. (1995). The expensive tissue hypothesis – the brain
and the digestive-system in human and primate evolution. Current Anthropology,
36, 199-221.
Akil, H., Brenner, S., Kandel, E., Kendler, K. S., King, M .- C., Scolnick, E., Watson, J. D.
and Zoghby, H. Y. (2010). The future of psychiatric research: genomes and neural
circuits. Science, 327, 1580-1581.
Bakermans-Kranenburg, M. J. and van IJzendoorn, M. H. (2006). Gene-
environment interaction of the dopamine D4 receptor (DRD4) and observed
maternal insensitivity predicting externalizing behavior in preschoolers.
Developmental Psychobiology, 48, 406-409.
Barkow, J. H., Cosmides, L. and Tooby, J. (1992). The Adapted Mind: Evolutionary
Psychology and the Generation of Culture. New York: Oxford University Press.
Baron-Cohen, S. (1994). How to build a baby that reads minds: cognitive
mechanisms in mindreading. Current Psychology of Cognition, 13, 513-552.
Baron-Cohen, S. (1997). The Maladapted Mind: Classic Readings in Evolutionary
Psychopathology. Hove: Psychology Press.
Baron-Cohen, S. (2020). The Pattern Seekers: How Autism Drives Human Invention.
New York: Basic Books.
Belsky, J. (1997). Variation in susceptibility to rearing influences: an evolutionary
argument. Psychological Inquiry, 8, 182-186.
Belsky, J., Steinberg, L. and Draper, P. (1991). Childhood experience, interpersonal
development and reproductive strategy: an evolutionary theory of socialization.
Child Development, 62, 647-670.
Blumstein, D. (2020). The Nature of Fear: Survival Lessons from the Wild.
Cambridge, MA: Harvard University Press.
Bowlby, J. (1969). Attachment and Loss, Vol. 1: Attachment. New York: Basic Books.
Brady, S. P., Bolnick, D. I., Angert, A. L., Gonzalez, A., Barrett, R. D. H., Crispo, E.,
Derry, A. M., Eckert, C. G., Fraser, D. J., Fussmann, G. F., Guichard, F., Lamy, T.,
McAdam, A. G., Newman, A. E. M., Paccard, A., Rolshausen, G., Simons, A. M. and
Hendry, A. P. (2019). Causes of maladaptation. Evolutionary Applications, 12,
1229-1242.
Brothers, L. (1997). Friday’s Footprint: How Society Shapes the Human Mind.
Oxford: Oxford University Press.
Brüne, M. (2007). On human self-domestication, psychiatry and eugenics.
Philosophy, Ethics and Humanities in Medicine, 2, 21.
Brüne, M. (2015). Textbook of Evolutionary Psychiatry & Psychosomatic Medicine:
The Origins of Psychopathology. Oxford: Oxford University Press.
Brüne, M., Belsky, J., Fabrega, H., Feierman, H. R., Gilbert, P., Glantz, K., Polimeni, J.,
Price, J. S., Sanjuan, J., Sullivan, R., Troisi, A. and Wilson, D. R. (2012). The crisis of
psychiatry – insights and prospects from evolutionary theory. World Psychiatry,
11,55-57.
Buller, D. J. (2005). Adapting Minds: Evolutionary Psychology and the Persistent
Quest for Human Nature. Cambridge, MA: MIT Press.
Burns, J. (2007). The Descent of Madness: Evolutionary Roots of Psychosis and the
Social Brain. London: Routledge.
Buss, D. M. (2000). The evolution of happiness. American Psychologist, 55, 15-23.
Buss, D. M. (2009). The great struggles of life: Darwin and the emergence of
evolutionary psychology. American Psychologist, 64, 140-148.
Calderoni, L., Rota-Stabelli, O., Frigato, E., Panziera, A., Kirchner, S., Foulkes,
N. S., Kruckenhauser, L., Bertolucci, C. and Fuselli, S. (2016). Relaxed selective
constraints drove functional modifications in peripheral photoreception of
the cavefish P. andruzzii and provide insight into the time of cave colonization.
Heredity, 117, 383-392.
Cavali-Sforza, L., Menozzi, P. and Piazza, A. (1994). The History and Geography of
Human Genes. Princeton, NJ: Princeton University Press.
Crespi, B. J. (2016). The evolutionary aetiologies of autism spectrum and psychotic
affective spectrum disorders. In A. Alvergne, C. Jenkinson and C. Faurie (eds.),
Evolutionary Thinking in Medicine: From Research to Policy and Practice (pp. 299-
327). Cham: Springer.
Darwin, C. (1859). On the Origin of Species by Means of Natural Selection. London:
Murray.
Darwin, C. (1871). On the Descent of Man, and Selection in Relation to Sex. London:
Murray.
Dean, L. G., Vale, G. L., Laland, K. N., Flynn, E. and Kendal, R. L. (2014). Human
cumulative culture: a comparative perspective. Biological Reviews, 89, 284-301.
Dehaene, S., Cohen, L., Morris, J. and Kolinsky, R. (2015). Illiterate to literate:
behavioural to cerebral changes induced by reading acquisition. Nature Reviews:
Neuroscience, 16, 234-244.
Del Giudice, M. (2017). Mating, sexual selection, and the evolution of
schizophrenia. World Psychiatry, 16, 141-142.
Del Giudice, M. (2018). Evolutionary Psychopathology: A Unified Approach. New
York: Oxford University Press.
Del Giudice, M. (2020). Rethinking the fast-slow continuum of individual
differences. Evolution and Human Behavior, 41, 536-549.
Dunbar, R. (2003a). Evolution of the social brain. Science, 302, 1160-1161.
Dunbar, R. (2003b). The social brain: mind language and society in evolutionary
perspective. Annual Review of Anthropology, 32, 163-181.
Dunbar, R. (2014). Human Evolution: A Pelican Introduction. London: Pelican.
Durrant, R. and Ellis, B. (2003). Evolutionary psychology: core assumptions and
methodology. In M. Gallagher and R. Nelson (eds.), Comprehensive Handbook of
Psychology. Vol. 3: Biological Psychology (pp. 1-35). New York: John Wiley and Sons.
Ellis, B. J., Shirtcliff, E. A., Boyce, W. T., Deardorff, J. and Essex, M. J. (2011). Quality
of early family relationships and the timing and tempo of puberty: effects depend
on biological sensitivity to context. Developmental Psychopathology, 23, 85-99.
Ewald, P. W. (1994). Evolution of Infectious Disease. Oxford: Oxford University
Press.
Fuchs, I. (2019). The Evolutionary Mechanism of Human Dysfunctional Behavior.
New York: Radius Book Group.
Fuglewicz, A. J., Piotrowski, P. and Stodolak, A. (2017). Relationship between
toxoplasmosis and schizophrenia: a review. Advances in Clinical and Experimental
Medicine, 26, 1031-1036.
Garland, T., Jr and Kelly S. A. (2006). Phenotypic plasticity and experimental
evolution. Journal of Experimental Biology, 209, 2344-2361.
Gilbert, P. (2020). Compassion: from its evolution to a psychotherapy. Frontiers in
Psychology, 11, 1-31.
Gilbert, P. and Bailey, K. (2000). Genes on the Couch: Explorations in Evolutionary
Psychotherapy. Hove: Brunner-Routledge.
Gluckman, P. D., Beedle, A. S. and Hanson, M. A. (2009). Principles of Evolutionary
Medicine. Oxford: Oxford University Press.
Gonzalez-Forero, M. and Gardner, A. (2018). Inference of ecological and social
drivers of brain size evolution. Nature, 557, 554-557.
Gould, S. J. (1997). Evolution: the pleasures of pluralism. New York Review of Books,
44, 47-52.
Hagen, E. H. (2016). Evolutionary psychology and its critics. In D. Buss (ed.),
Handbook of Evolutionary Psychology, Vol. 1, 2nd Ed. (pp. 136-160). Hoboken, NJ:
Wiley.
Hagen, E. H. (2020). Is evolutionary psychology impossible? https://
thisviewoflife.com/is-evolutionary-psychology-impossible/ (accessed 30
November 2020).
Haltigan, J., Del Giudice, M. and Khorsand, S. (2021). Growing points in
attachment disorganization: looking back to advance forward. Attachment and
Human Development, 23, 438-454.
Hamilton, W. D. (1964). The genetical evolution of social behaviour. I. Journal of
Theoretical Biology, 7, 1-16.
Henn, B. M., Cavalli-Sforza, L. L. and Feldman, M. W. (2012). The great human
expansion. Proceedings of the National Academy of Sciences of the United States of
America, 109, 17758-17764.
Henrich, J. (2016). The Secret of Our Success: How Culture Is Driving Human
Evolution, Domesticating Our Species, and Making Us Smarter. Princeton, NJ:
Princeton University Press.
Henrich, J. (2020). The Weirdest People in the World: How the West Became
Psychologically Peculiar and Peculiarly Prosperous. London: Allen Lane.
Henrich, J. and Tennie, C. (2017). Cultural evolution in chimpanzees and humans.
In M. Muller, R. Wrangham and D. Pilbeam (eds.), Chimpanzees and Human
Evolution (pp. 645-702). Cambridge, MA: Belknap Press of Harvard University
Press.
Homberg, J. R. and Lesch, K. P. (2011). Looking on the bright side of serotonin
transporter gene variation. Biological Psychiatry, 69, 513-519.
Humphrey, N. K. (1976). The social function of intellect. In P. P. G. Bateson and R.
A. Hinde (eds.), Growing Points in Ethology (pp. 303-317). Cambridge: Cambridge
University Press.
Insel, T. R. and Cuthbert, B. N. (2015). Brain disorders? Precisely. Science, 348, 499-
500.
Jacobs, F. (1977). Evolution as tinkering. Science, 169, 1161-1166.
Kay, C., Tirado-Hurtado, I., Cornejo-Olivas, M., Collins, J. A., Wright, G., Inca-
Martinez, M., Veliz-Otani, D., Ketelaar, M. E., Slama, R. A., Ross, C. J., Mazzetti, P.
and Hayden, M. R. (2017). The targetable A1 Huntington disease haplotype has
distinct Amerindian and European origins in Latin America. European Journal of
Human Genetics, 25, 332-340.
Kendler, K. S. (2008). Explanatory models for psychiatric illness. American Journal
of Psychiatry, 165, 695-702.
Laland, K. (2017). Darwin’s Unfinished Symphony: How Culture Made the Human
Mind. Princeton, NJ: Princeton University Press
Laland, K. and Brown, G. (2011). Sense and Nonsense: Evolutionary Perspectives on
Human Behaviour (2nd Ed.). Oxford: Oxford University Press.
Laland, K., Odling-Smee, J. and Endler, J. (2017). Niche construction, sources of
selection and trait coevolution. Interface Focus, 7, 20160147.
Lents, N. H. (2020). Human Errors: A Panorama of Our Glitches from Pointless Bone
to Broken Genes. London: Weidenfeld and Nicholson.
Li, N. P., van Vugt, M. and Colarelli, S. M. (2018). The evolutionary mismatch
hypothesis: implications for psychological science. Current Directions in
Psychological Science, 27, 38-44.
Mayr, E. (1961). Cause and effect in biology. Science, 134, 1501-1506.
Mayr, E. (1971). The nature of the Darwinian revolution. Science, 176, 981-989.
Mayr, E. (1974). Behavior programs and evolutionary strategies. American
Scientist, 62, 650-659.
McGuire, M. T. and Troisi, A. (1998). Darwinian Psychiatry. New York: Oxford
University Press.
Nesse, R. M. (2005). Maladaptation and natural selection. Quarterly Review of
Biology, 80, 62-70.
Nesse, R. M. (2011). Ten questions for evolutionary studies of disease
vulnerability. Evolutionary Applications, 4, 264-277.
Nesse, R. M. (2013) Tinbergen’s four questions organized: a response to Bateson
and Laland. Trends in Ecology & Evolution, 28, 681-682.
Nesse, R. M. (2016). Evolutionary psychology and mental health. In D. Buss (ed.),
Handbook of Evolutionary Psychology, Vol. 2, 2nd Ed. (pp. 1007-1026). Hoboken,
NJ: Wiley.
Nesse, R. M. (2019). Good Reasons for Bad Feelings: Insights from the Frontiers of
Evolutionary Psychiatry. London: Allen Lane.
Nesse, R. M. and Dawkins, R. (2010). Evolution: medicine’s most basic science.
In D. A. Warrell, T. M. Cox, J. D. Firth and E. J. J. Benz (eds.), Oxford Textbook of
Medicine, 5th Ed. (pp. 12-15). Oxford: Oxford University Press.
Nesse, R. M. and Stein, D. (2019). How evolutionary psychiatry can advance
psychopharmacology. Dialogues in Clinical Neuroscience, 21, 167-175.
Nesse, R. M. and Williams, G. (1994). Why We Get Sick: The New Science of
Darwinian Medicine. New York: Times Books.
NIH (2020) National Human Genome Research Institute. www.genome.gov/
genetics-glossary/Mutation (accessed 30 November 2020).
Pavličev, M., Romero, R. and Mitteroecker, P. (2020). Evolution of the human pelvis
and obstructed labor: new explanations of an old obstetrical dilemma. American
Journal of Obstetrics and Gynecology, 222, 3-16.
Pilbeam, D. R. and Lieberman, D. E. (2017). Reconstructing the last common
ancestor of chimpanzees and humans. In M. Muller, R. Wrangham and D. Pilbeam
(eds.), Chimpanzees and Human Evolution (pp. 22-141). Cambridge, MA: Belknap
Press of Harvard University Press.
Pinker, S. (1994). The Language Instinct: How the Mind Creates Language. New York:
Harper Collins.
Pollard, T. (2008). Western Diseases: An Evolutionary Perspective. Cambridge:
Cambridge University Press.
Rantala, M. J., Luoto, S., Krama, T. and Krams, I. (2019). Eating disorders: an
evolutionary psychoneuroimmunological approach. Frontiers in Psychology, 10,
2200.
Richerson, P. and Boyd, R. (2005). Not by Genes Alone: How Culture Transformed
Human Evolution. Chicago, IL: University of Chicago Press.
Russell, G. (2000). Disorders of eating. In M. G. Gelder, J. J. Lopez-Ibor Jr and N. C.
Andreasen (eds.), New Oxford Textbook of Psychiatry, Vol. 1 (pp. 835-855). Oxford:
Oxford University Press.
Stearns, S. C. (1992). The Evolution of Life Histories. Oxford: Oxford University
Press.
Stevens, A. and Price, J. (2000). Evolutionary Psychiatry: A New Beginning, 2nd Ed.
London: Routledge.
Swedo, S., Leonard, H. and Kiessling, L. S. (1994). Speculations on anti-neuronal
antibody-mediated neuropsychiatric disorders of childhood. Pediatrics, 93, 323-
326.
Tanaka, K. D., Morimoto, G., Stevens, M. and Ueda, K. (2011). Rethinking visual
supernormal stimuli in cuckoos: visual modeling of host and parasite signals.
Behavioral Ecology, 22, 1012-1019.
Taylor, J. (2015). Body by Darwin: How Evolution Shapes our Health and Transforms
Medicine. Chicago, IL: University of Chicago Press.
Tinbergen, N. (1963). On aims and methods of ethology. Zeitschrift Für
Tierpsychologie, 20, 410-433.
Tomasello, M. (1990). Cultural transmission in tool use and communicatory
signaling of chimpanzees? In S. Parker and K. Gibson (eds.), ‘Language’ and
Intelligence in Monkeys and Apes: Comparative Developmental Perspectives (pp. 274-
311). Cambridge: Cambridge University Press.
Tooby, J. and Cosmides, L. (1990). The past explains the present: emotional
adaptions and the structure of ancestral environments. Ethology and Sociobiology,
11, 375-424.
Tooby, J. and Cosmides, L. (1992). The psychological foundations of culture. In
J. H. Barkow, L. Cosmides and J. Tooby (eds.), The Adapted Mind: Evolutionary
Psychology and the Generation of Culture (pp. 19-136). New York: Oxford
University Press.
Trevathan, W. R., Smith, E. O. and McKenna, J. J. (2008). Evolutionary Medicine and
Health: New Perspectives. New York: Oxford University Press.
Trivers, R. L. (1972). Parental investment and sexual selection. In B. Campbell
(ed.), Sexual Selection and the Descent of Man, 1871-1971 (pp. 136-179). Chicago,
IL: Aldine.
Trivers, R. L. (1974). Parent-offspring conflict. American Zoologist, 14, 249-264.
Troisi, A. (2015). The evolutionary diagnosis of mental disorder. WIREs Cognitive
Science, 6, 323-331.
Trull, T. J. and Widiger, T. A. (2013). Dimensional models of personality: the five-
factor model and the DSM-5. Dialogues in Clinical Neuroscience, 15, 135-146.
Waddington, C. H. (1942). Canalization of development and the inheritance of
acquired characters. Nature, 150, 563-565.
Williams, G. C. and Nesse, R. M. (1991). The dawn of Darwinian medicine.
Quarterly Review of Biology, 66, 1-22.
Wilson, D. S. (2019). This View of Life: Completing the Darwinian Revolution. New
York: Pantheon Books.
Wrangham, R. (2009). Catching Fire: How Cooking Made Us Human. London: Profile
Books.
Young, E. S., Frankenhuis, W. E. and Ellis, B. J. (2020). Theory and measurement of
environmental unpredictability. Evolution and Human Behavior, 41, 550-556.
Zietsch, B. P. and Sidari, M. J. (2020). A critique of life history approaches to human
trait covariation. Evolution and Human Behavior, 41, 527-535.
- We are grateful to Lindsey Edwards, Adam Hunt and Annie Swanepoel for
providing valuable comments on previous drafts of this chapter.
1 Readers interested in a more detailed introduction to evolutionary science
relevant to medicine may wish to consult Principles of Evolutionary Medicine
(Gluckman et al., 2009) or later editions.
第2章
進化理論によって深化された生物・心理・社会モデル
Adam Hunt、Paul St John-Smith、Riadh Abed
要約
現在、精神医学と医学において健康と疾病を説明する主流モデルは、1970年代にEngelが提唱した生物・心理・社会(BPS)モデルである。これは、還元主義的な生物医学に心理的・社会的要因を統合するために提案された。BPSモデルは進歩ではあったが、その科学的・哲学的基盤には疑問が残り、完全あるいは十分なモデルとは言えない。本章では、BPSモデルの歴史的・概念的分析を行った後、進化理論の統合がBPSモデルの次の段階として適切であることを示す。進化理論は、BPSモデルが認識する多層的な因果関係を正当化し強化するとともに、究極的因果関係と近接的因果関係の両方を認識することでモデルを拡張する。これにより、生物学的機能と機能障害の明確な区別が可能となり、系統発生学的視点が生物学研究を新たな方向に導く。健康のモデルを生物学の最も基本的な理論と結びつけるこのアプローチは、BPSモデルが深刻に欠いていた哲学的・科学的整合性を提供する。
キーポイント
・生物・心理・社会モデルは、主流の還元主義的生物医学モデルを前進させた。
・進化的視点は、還元主義的生物医学アプローチの限界を明らかにする。
・進化理論は、機能と系統発生の分析を統合することで生物・心理・社会モデルを拡張する。
・進化理論は、生物学的機能と機能障害を理解するための優れた科学的基盤を提供する。
2.1 現代医学モデル
2.1.1 生物医学の導入
英国では「生物医学(biomedicine)」という用語が初めて登場したのは1923年のDorland医学辞典で(Quirke and Gaudillière, 2008)、「生理学と生化学の原理に基づく臨床医学」を意味した。20世紀の医学は次第に実験室科学と結びつき、健康研究はより厳密な実験手法(Bynum et al., 2006)と「根拠に基づく医療(EBM)」へと向かった。現代の規制当局は、治療法の承認にあたり、生化学的またはその他の病態生理学的経路および薬理学的機序に関する科学的根拠を要求する(Van Norman, 2016)。こうして医学は、何世紀にもわたって体液説や精神的病弊など有害性の高い治療を生んだ呪術的医療(Wootton, 2006)から、生物学的変化と介入を伴う実践へと転換した。
疾患を機能障害的な生物学的(以下「身体的」)プロセスとして定義するこのアプローチが主流精神医学に遅れて到達した。20世紀前半は精神分析が支配的だった精神医学(Shorter, 1997)は、1980年の『精神障害の診断と統計マニュアル第3版(DSM-III)』の出版により生物医学的アプローチへ移行した。この転換はRobert Spitzerの指導のもと行われ、他の医学領域で成功したモデルを模倣することが目的だった(Surís et al., 2016)。この生物医学的アプローチの支持者は「生物学的精神科医」と呼ばれることもある。
1970年代に精神医学に入り込んだ還元主義的生物医学アプローチは、「反精神医学(antipsychiatry)」(Pilgrim, 2002)から強力な反発を受けた。最も有名なのはThomas Szaszで、彼は「精神疾患」は人間の生活上の問題を表す単なる比喩に過ぎず、社会的に混乱をきたす行動や思考はあるが、アルツハイマー病やハンチントン病など一部の脳疾患を除けば、物理的疾患としての「病気」ではないと主張した。Szaszは、「診断を確定するための生物学的・化学的検査も、生検や剖検所見も存在しない」(Szasz, 2008: 2-5)と指摘した。
こうした批判をかわす一つの方法は、神経科学と遺伝学の研究がまもなく精神疾患を確実に説明する疾病プロセスを実証するという宣言だった。この立場は約50年間続いている(Linden, 2012)が、還元主義的生物医学的説明が発見されるという単純な考えへの信頼は薄れつつある。特に、還元主義的分析を拒む複雑な相互作用ネットワークを考慮する場合そうだ(Borsboom et al., 2019)。医療現場では、還元主義的生物医学モデルからの最も広く認知され採用された進歩は、George Engelの「生物・心理・社会(BPS)」モデル(Engel, 1977, 1980)であり、「現代精神医学の現状(status quo)」(Ghaemi, 2009)と評されている。
2.1.2 EngelのBPSモデルの起源と基盤
EngelはBPSモデルをシステム理論(von Bertalanffy, 1968; Weiss, 1969)の理論モデルに基づき、相互作用する組織の異なるレベルにおける物理的システムの階層を特定した:素粒子、原子、分子、細胞小器官、細胞、組織、器官・器官系、神経系、個人、二人、家族、地域社会、文化・下位文化、社会・国家、生物圏。階層の各レベルは、組織化された動的全体を表すとされた。極端な生物医学モデルでは、医師は単なるバイオマーカーの探索者と生理状態の修復者(例:Andreasen, 1984)に過ぎない。Engelは、これは「個人」以下のレベルにしか当てはまらず、人間が置かれた関係、家族、地域社会、文化といった上位階層の下方への影響を無視していると主張した。
抑うつを例にとると(本章で繰り返し取り上げる)、Engelの不満は、抑うつをセロトニン取り込みの異常な変動などの脳障害としてのみ扱う視点に向けられる。彼は、心理社会的レベルで容易に観察できる要因によって引き起こされる多くの抑うつ事例を見落としていると主張するだろう(第8章参照)。
Engelの当初の2つの論文は、政治的目的(精神分析学者と新興の生物医学研究者の統合)に触発されたもの(Shorter, 2005)とも言えるが、身体的レベルと同様に心理社会的レベルでの観察と介入の重要性を強調した。Bolton and Gillett(2019)などの最近のBPSモデルの更新とは異なり、この当初の定式化は「生物」「心理」「社会」を厳密に定義し区別することに関心がなかった。Engelは、還元主義と生物医学的独断への反対以外の哲学的議論を避け、適切なケアと健康障害の完全な理解には患者の心理状態と社会的状況の認識が必要だという単純な点を主張した。抑うつに関連する神経学的変化を完全に理解したとしても、その人が最近職を失ったか離婚したかといった基本的な情報を見逃す可能性がある。Engelはまた、生物医学的アプローチが特定の状態を変えるのに成功したとしても、患者は独自の精神的状態と社会的状況を持つ主体として考慮されるべきだと強調した。これはBPSモデルの批評家(Borell-Carrió et al., 2004)の間でさえ議論の余地がなく、現在の治療と研究の多くに(体系的でないにせよ)反映されている。心理的プロセスとプラセボ反応は医療介入における一般的な関連要因として受け入れられ、さらに社会的状況の健康への影響は広く認識されている(Braveman and Gottlieb, 2014)。
実際、今日の社会的状況と健康の関係の探究は著しく広がり、公衆衛生学者(Smith, 2000)、疫学者(Syme, 1987)、人類学者(Hoke and McDade, 2014)、社会学者(Conrad and Kern, 1981)などによる個別の広範な研究が行われている。この研究はさまざまな分野に広がり、「健康の社会的決定要因(social determinants of health)」(Braveman and Gottlieb, 2014)、「健康の政治経済学(political economy of health)」(Doyal, 1979)、「社会疫学(social epidemiology)」(Krieger, 2001)、「生態社会(ecosocial)」(Krieger, 1994)、「生物社会(biosocial)」(Hoke and McDade, 2014)、「生物文化(biocultural)」(McElroy, 1990)などの代替用語が導入されることが多い。タイトルや本文で「生物・心理・社会」という用語に言及されないことも多いが、直接関連している。この研究はEngelと同様に、国際的・国家的な公衆衛生アジェンダ(Kelly et al., 2009; Marmot, 2005)で引用され、ストレス、幼少期の逆境、社会的排除、仕事、失業、社会的支援、中毒、栄養と食事、交通(Marmot and Wilkinson, 1999)、社会的地位(Marmot, 2006)、人種差別(Harrell et al., 2011)、負債(Sweet et al., 2013)、近隣環境(Ludwig et al., 2011, 2012)など、身体的・精神的健康に影響する多数の社会的要因が指摘されている。この研究の量は「生物・心理・社会」という表現を使用した論文数をはるかに超えているが(Álvarez et al., 2012)、「生物・心理・社会」は、さまざまな大規模機関(Alonso, 2004)、医学部、精神医学(Adler, 2009; Mai, 1995; Pilgrim, 2002)において、多次元的医療の一般的概念を説明するための選択用語として残っている。
2.1.3 BPSモデルへの批判と更新
大多数の著者(批判派のSadler and Hulgus, 1990; Suls and Rothman, 2004を含む)は、Engelと同様に、純粋な生物医学的医療は過度な還元主義に陥りがちだという点で一致している。しかし、BPSモデルに対しては主に2種類の批判(Bolton and Gillett, 2019)が提起されている。第一に、哲学的明確性や科学的妥当性に欠けるという点。第二に、曖昧すぎて臨床的に実践不可能だという点である。
第一に、BPSモデルの哲学的・科学的価値は複数の著者(例:Benning, 2015)によって問われてきた。科学的理論やモデルとしての基準を満たしていない(McLaren, 1998)と批判され、精神分析を「裏口から」存続させようとする試み(Shorter, 2005)とも評された。さらに「社会」次元は、公衆衛生上の懸念事項(Kelly et al., 2009)である汚染、放射線、有害物質、感染症、騒音、食事などの重要な環境要素を除外しているように見える。このため近年の提唱者らは環境要因を付録として追加し(Bolton and Gillett, 2019)、モデルの完全性を高めようと試みている。重要なのは、Engelの主眼は哲学的妥当性ではなかった(Álvarez et al., 2012)と考えられる点だ。彼の関心は生物医学が特定の要素を過小評価していることであり、BPSモデルは医療実践への時宜を得た批判として提案され、独立した健康問題の理論モデルとしてではなかった。
第二の批判領域であるBPSモデルの曖昧さと臨床的非実用性は、このモデルが規範的でも精確でもない(Sadler and Hulgus, 1992)という観察に基づく。相互作用するレベルが疾病を引き起こすと認識する以上に、Engelは特定患者に対する診断・治療アプローチについて臨床医に明確な指針を提供していない(ただし近年の著者らはこのプロセスを形式化・例示しようと試みている;例:Evers et al., 2014; Smith et al., 2013)。BPSモデルの著名な批判者Ghaemi(2009, 2010)は、このモデルが本質的に恣意的な治療法に退化し、個々の臨床医が「生物」「心理」「社会」のいずれに重点を置くも自由に正当化され得ると指摘する。この曖昧さは研究課題の開発困難(Pilgrim, 2002)にも寄与している。「生物・心理・社会」という用語は現在広く建前として用いられるが、生物医学的研究アジェンダが依然支配的であり、Engelの発表以来ほとんど変化がない(Alonso, 2004; Suls and Rothman, 2004)。このため精神医学内外の研究者から懸念が表明され、精神医学は危機の学問分野(Bracken et al., 2012; Deacon, 2013)と評されるに至っている。
Bolton and Gillett(2019)は最近、BPSモデルを更新しこれらの批判に対処しようと試み、「生物」「心理」「社会」の区別と定義を正当化する哲学的議論を展開した。我々はFrazier(2020)と同様、これは「やや狭量な哲学的評価」でありBPSモデルを真に前進させるものではなく、多くの新たな批判に晒されると考える。例えばBolton and Gillett(2019)はBPSモデルの心理領域を定義する際「主体性(agency)」という単一概念を導入するが、内的葛藤、無意識的プロセスなど重要な心理過程の全範囲から遊離しているように見える。我々の見解では、彼らの試みは実践的行動計画として意図されたモデルの隙間を埋めるための後付け議論であり、厳密な科学理論としてBPSモデルを正当化できていない。BPSモデルを複雑化するだけで、健康の厳密で包括的なモデルとしての正当性を確立できていないのである。我々はここで、臨床的有用性のみならず理論的整合性と科学的有益性を備えた健康・疾病モデルに向かうためには、こうした哲学的議論が決定的に欠いているのは、すべての生物科学を変革した鍵となる科学的洞察――すなわちダーウィンの自然選択による進化論(Darwin, 1859)――だと提言する。
2.2 BPSモデルを超えて
2.2.1 生物医学からBPSへ、そして進化論へ
第1章で探究したように、チャールズ・ダーウィンの『種の起源』は自然選択による進化論を提示し、生物科学にパラダイムシフトを引き起こした。生物学において進化論は本質的に回避不能な自然法則であり、時空を超えて全ての生命形態に適用される。生物学における因果関係は複数のレベル(Mayr, 1961)を含み、アミノ酸、タンパク質、RNA、DNA、細胞、微細構造、細胞小器官、器官、生物体が相互作用する複雑なシステムを構成する(第1章参照)。EngelのBPSモデルはシステム理論内でこれらの相互作用レベルを取り込み、生物医学的アプローチに対するBPSモデルの優位性を正当化するとともに、この複雑な因果関係が生物医学的還元主義の完全な説明可能性を排除することを認めた。近年のネットワークモデル(Barabási and Oltvai, 2004; Borsboom et al, 2019)は、相互作用する多レベル複雑性という文脈におけるこの因果関係観の現代的バージョンと言える。
EngelがBPSモデルを提案した目的は、生物医学を超えた因果関係と介入レベルに注意を向けることにあった。彼の関心次元は微視的効果から巨視的効果、細胞レベルから社会全体まで、空間分析の多レベル認識だった。しかし生物学における因果関係は、歴史的プロセスの産物として時間的にも理解される必要がある――機序から個体発生、進化的機能の歴史へ、数百万年にわたって分岐する複数種にわたって。これらはティンバーゲンの4問(Tinbergen, 1963;詳細は本巻第1章参照)によって進化科学者に認識され、これはマイヤーの近接因果と究極因果の区別(Mayr, 1961)をさらに展開したものだった。
表2.1ではMedicus(2005)に着想を得て、抑うつの進化的生物・心理・社会像を得るため、ティンバーゲンの4問とEngelの3レベルからマトリクスを作成した。これはBPSモデルが生物医学をどう前進させたか(機序と発達という近接問を心理・社会レベルに拡張)、そして進化論がBPSをどう前進させるか(全ての分析レベルにおいて機能と系統発生という究極問を拡張)を示している。
表2.1
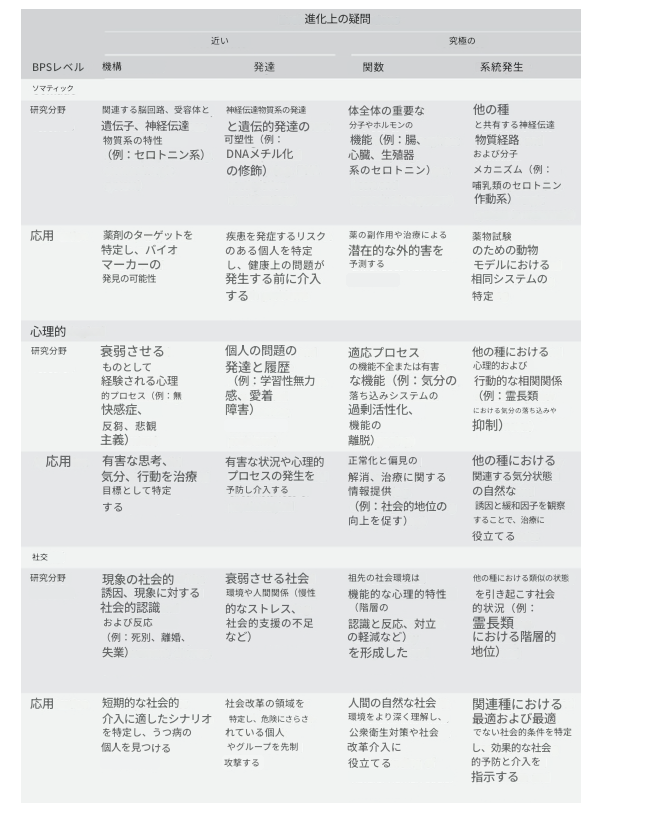
バイオメディカルの枠組みでは、左上の「体細胞-近位」の原因が主な関心事です。
エンゲルのBPSモデルは、この近位の列を下に拡張し、「心理的-近位」と「社会的-近位」を含めることで、パラダイムを進歩させました。これらには、心理的および社会的な分析の領域とその結果としての応用が含まれており、BPSモデルは、開業医のより包括的な役割を奨励し、公衆衛生の研究と社会レベルでの介入への扉を開きました。
進化医学と進化精神医学は、機能と系統発生の「究極」の分析を含めるように、3つのレベルすべてを拡張し、各レベルでの研究と治療に関連性を持たせています。体細胞レベルの究極の分析は、医学研究においてすでに実用的な有用性を持っていることに注意してください。研究者は、主要な分子が全身に存在し、私たちの生物学が他の種と比較可能であることを認識しています。これにより、介入の副作用を理解し、動物モデルを使用した実験への扉が開かれます。しかし、この慣行は、自然選択による進化がなぜ副作用が例外ではなく規範であるのか(進化は新しい機能のために既存の分子やシステムを流用する)と、動物モデルが有用である理由(生物学的システムは系統発生全体で共有されているため、非ヒト動物での実験により、ヒトでの結果を予測できる)を説明する進化論を認識することなく発展してきました。この説明理論をより明確に認識することで、副作用のより良い予測と軽減、そして動物モデルのより優れた使用につながる可能性があります。
健康に関連する心理的および社会的レベルの研究は、進化論の潜在的な恩恵をほとんど受けていません。これはおそらく、これらのレベルでの理論化を支配している哲学的、心理学的、社会学的視点が、進化論および比較種間研究とほとんど結びついていないことによる部分的な結果でしょう。
進化心理学、進化人類学、霊長類学は、逆に、ほとんど非医学的であり、機能不全ではなく、見かけの機能に焦点を当ててきました。したがって、これは依然としてほとんど未開拓の研究分野です。
2.2.2 心理社会的な究極の理解
第1章では、人類の文化的進化と社会脳の概要を示しました。これは、進化論的アプローチがうつ病の心理的および社会的分析をどのように強化するかを検討する上で関連するトピックです。
狩猟採集民の研究は、私たちの祖先がかなり平等主義的な小規模社会に住んでいたことを示唆しています。ほとんどの狩猟採集民の平等主義的な性質にもかかわらず、社会階層とある程度の不平等が存在し(Boehm, 2012; Chagnon, 2013)、これらは生殖成功に影響を与えました(Alami et al., 2020; von Rueden and Jaeggi, 2016)。これらの選択圧は、地位の低下後の気分の落ち込みが、無益で危険な争いを減らすリスクを減らすなど(Price et al., 1994; この巻の第8章も参照)、集団内の社会階層に関連する心理的適応を促したと考えられます。ミスマッチは、この自然な気分の落ち込みの傾向が、現代社会において臨床的うつ病に発展する原因となる可能性があります。現代の社会的流動性は制約されており、地位の喪失は、深刻な経済不況や技術革新の間に特定の職業が失われるなど、変化させることができない現代的な変数による可能性があります。狩猟採集社会では、そのようなシステム上の問題は存在しませんでした。個人は気分の落ち込みに対応して、別の集団に移ったり、称賛に値する偉業を成し遂げたり、新しい同盟を結んだりするなどして、状況を改善することができました。したがって、気分の落ち込みは、臨床的うつ病に見られるように、持続的に深まり麻痺させるのではなく、生産的で自己限定的である可能性があります。祖先環境では、複数の社会的支援の利用可能性が、気分の落ち込みの期間と強度を緩和し、したがって病的なうつ病状態への進行の可能性を減らすことができました(Rantala, 2018)。
もちろん、健康問題の理解への進化論的アプローチは、人間の自然な生息地と人類の進化の歴史の考察だけでなく、他の種との系統発生学的比較も奨励します。社会的敗北後の抑うつ気分は、配偶者や資源をめぐって対立する近隣の霊長類の集団という祖先環境において機能的であった可能性があり、さらなる紛争からの離脱を促すことで保護的な目的を果たしました(Gilbert and Allan, 1998; Sloman et al., 2003)。霊長類の研究では、循環グルココルチコイド(ストレスのマーカー)のレベルが、社会的地位の低下とともに徐々に増加することが示されています。しかし、これは、下位の個体が優位の個体からの攻撃を受け、近くに支援的な社会的つながりや親族がいない場合にのみ発生します(Abbott et al., 2003)。ストレスと抑うつ的な行動は、非ヒト霊長類でも観察されており、その原因と可能な軽減要因を理解するための示唆を与えています(Shively and Willard, 2012)。このような系統発生学的研究は、人間にとって潜在的に有害で有益な社会的配置を示す可能性があります。
BPSモデルは、例えば、強力な社会ネットワークが特定の健康問題を軽減できることを研究者に認識させるなど、バイオメディカルモデルを改善しましたが、その説明の浅さから、情報に基づかない実りのない対応につながる可能性があります。例えば、うつ病がしばしば失業後に起こる場合、現代的な観点からの合理的な仮定は、うつ病は経済的損失によって引き起こされるということかもしれません。つまり、個人の経済状況を改善すること(例えば、失業手当によって)がうつ病の改善に役立つはずだということです。また、うつ病で失業している人は、認知行動療法などの通常のうつ病の介入に反応すると想定されるかもしれません。しかし、うつ病が主に地位と社会ネットワークの喪失に関連している場合(進化論的観点からは、財政よりも心理的に重要であると認識されます)、財政改善やセラピーだけではうつ病にほとんど効果がないと予想されます。失われた地位と社会的なつながりを取り戻すライフスタイルの変化、おそらく別の仕事を見つけることが伴う必要があります。これは実際にそのようであるようです(Moore et al., 2017)。
具体的な例を超えて、進化論は、心理社会的要因と健康転帰との関係に関する一般的なメカニズムを理解するために使用できます。例えば、発達可塑性(第1章を参照)は選択によって形成されており、社会的身分、自尊心、アイデンティティ、離婚、死別、失業、逮捕、退職など、病因の観点からはまだ十分に理解されていない健康に影響を与えるさまざまな要因を説明するのに役立つ可能性があります(Cohen et al., 2019)。これらの負の健康影響につながる根本的なメカニズムを理解するためには、究極の進化論的な問いを適用する必要があります(Troisi, 2020)。人間が発達的に可塑的である(Lea et al., 2017)または感受性のレベルが異なる(Albott et al., 2018)場合、この適応プロセスは誤作動する可能性があります。可塑性が進化するためには、祖先的に平均して生殖成功を提供する必要がありましたが、必ずしも完璧である必要はありませんでした。障害は、ストレス要因が寿命または重症度の閾値を超えた場合など、偶発的な不幸な副産物として生じる可能性があります。可塑性(特に神経可塑性)は必然的に誤りやすい可能性があり、この誤りやすさは現代のミスマッチ環境で増幅される可能性があります。この分析は、可塑性と障害の背後にある近位の神経生物学的システムに関する研究を補完することに注意してください。次に、進化論的視点は、なぜそれらのシステムとその脆弱性が存在するのかを説明し、量的遺伝学と環境暴露を結びつける広範な行動遺伝学の研究を裏付けます(Assary et al., 2018)。BPSモデルが公衆衛生の考慮事項を近位の心理社会的要因を含むように拡大したのに対し、進化医学と進化精神医学はこれらのシステムの理解を深めます。
2.2.3 プラセボ効果と防御
プラセボ効果の研究は、臨床的改善の期待と条件付けを含み、肯定的な健康転帰をもたらす特定の試験状況に特有の心理社会的文脈の研究です(Colloca and Benedetti, 2005; Finniss and Benedetti, 2005)。プラセボ効果は、体細胞、環境、心理的、社会的な境界の多孔性の明確な証拠です。環境的および心理社会的な出来事は、患者の脳と体に生化学的変化を引き起こし、それが今度は病気の経過と治療結果に影響を与えます(Benedetti, 2008)。プラセボ効果では、対人関係と社会環境が、癒しとケアがどのように機能するかを説明する上で不可欠な要素になります(McQueen et al., 2013)。
プラセボ反応により、哺乳類は内部プロセスと行動を修正することができます。そして、外部環境、社会的相互作用、将来の環境の評価に関する正確な肯定的評価に照らして、内部環境を修正するこれらの能力から、複数の適応上の利点が生じる可能性があります(McQueen et al., 2013)。Nesse(2019)は、プラセボ反応は主に病気のプロセスを変更するのではなく、体の防御の修正を伴うことを強調しています(ボックス2.1を参照)。進化は、傷害、感染、または中毒から防御する生物学的メカニズムを選択しており、これらの防御の調節は環境の評価によって影響を受けます。しかし、多くの防御は過剰に発現しているように見えます。これは、「信号検出分析」の枠組みの中で理解できます。生物がわずかなコストで保護的な方法で反応できるが、潜在的に莫大な利益(例えば、死を回避する)がある場合、最適なシステムは多くの誤警報を発します。嘔吐は数百カロリーと数分しか消費しないかもしれませんが、嘔吐しないことは中毒による死のリスクを伴います。これは「煙探知機原理」と呼ばれています(Nesse, 2001; この巻の第7章を参照)。防御の調節はまた、そうでなければ「保護的」な防御を、脱出を容易にするために極度の危険な状況下で、そしてもはや必要のない回復に適した状況下でオフにすることも可能にします。進化論的視点からこの複雑で遍在的な人間のヘルスケアの要素を理解することは、現代の医学的議論に欠けている有用な洞察を提供します。
ボックス2.1 体の防御の例
- 体性感覚:痛み、倦怠感、吐き気、不安、抑うつ、発熱、かゆみ
- 骨格/運動反応:逃走、凍結
- 内臓:くしゃみ、咳、嘔吐、下痢
- 体液性反応:免疫調節、視床下部-下垂体-副腎皮質軸の調節
2.3 非還元主義の正当化
現代の精神科医の懸念は、エンゲルが提唱したBPSモデルが精神医学研究や実践に十分な影響を与えていないことです(Deacon, 2013)。ある程度、これはその哲学的および科学的欠陥の必然的な結果でした。その影響は、その理論的な欠点によって制限されてきました。ここでは、進化医学が、より理論的に満足のいく方法で、BPSモデルの中核的な原則、つまり体細胞、心理的、社会的な相互作用するレベルをどのように正当化し、拡張できるかを検討します。
2.3.1 双方向の因果関係
進化論は、体細胞、心理的、社会的なレベル間の双方向の因果関係を認識しています。心理的および社会的なレベルは、選択圧を通じて世代を超えて体細胞を変化させますが、常に体細胞のプロセス(神経生物学的、遺伝的など)によって媒介されています。例えば、セロトニン作動性システムは、気分と心理的および社会的相互作用に影響を与える遺伝子によって作成され、それらの心理的および社会的影響が適合性を高めるかどうかは、セロトニン作動性システムの進化に影響を与えます。したがって、体細胞システムは、その心理的および社会的影響に関連する正当な進化上の理由のために存在します(Andrews et al., 2015)。医学において進化論的視点を取り入れることは、遺伝学を逆転させることに等しいです。うつ病の非進化論的な生物医学的見解は、特定のDNA配列をその状態の主要な生物学的原因と見なすかもしれませんが、進化論的アプローチは、進化の歴史を通じてこれらの遺伝子の存続につながった選択圧を理解しようともします(例えば、Raison and Miller, 2013)。
2.3.2 多重実現可能性
進化論的アプローチの主な強みの1つは、生物の包括的適応度(自然環境において)に貢献した特性を特定することで、選択の最終産物を特定できることです。選択された表現型がより低いレベルの組織化で多重に実現される可能性があるという洞察は、非還元的な分析をさらに正当化します(Borsboom et al., 2019; Krakauer et al., 2017)。多重実現可能性の例は豊富にあります。そのような例の1つは、マラリア感染を予防または軽減するために進化した表現型であり、これは異なる遺伝的継承を持つヘモグロビンの複数の分子配置を通じて達成されます(Weatherall and Clegg, 2001)。純粋な還元主義はこれらを異なる現象と誤解するでしょう。マラリアが豊富な環境における個体レベルの特性の進化論的分析は、それらの共有された究極の原因を認識するために必要です。現象が進化的視点と完全な社会的/環境的文脈の中で探求されない限り、重要な機能は認識されない可能性があります。したがって、農業が蚊、ひいてはマラリアを助長するという認識(Janko et al., 2018)も、マラリアの世界的な死亡率を完全に理解するために必要であり、プラスモジウム寄生虫の還元主義的な分析だけでは不十分です。
同じ適応機能に対する複数の体細胞実現の他の顕著な例が存在します。成人期までラクターゼ酵素を持続させるラクターゼ持続性は、いくつかの集団で、そしてそれらの集団内の異なるメカニズムを通じて、繰り返し独立して進化しました(Campbell and Ranciaro, 2021)。薄い肌は、北東アジアと北西ヨーロッパの低紫外線地域で、異なる遺伝的要因によって独立して進化しました(Hider et al., 2013; Norton et al., 2007)。高地の低酸素条件下で生きるための適応は、アンデス(ボリビアとチリ)、チベット、エチオピアの高地の人々において異なる経路を通じて進化しました(Beall, 2007; Bigham et al., 2010; Scheinfeldt et al., 2012; Xu et al., 2011)。これらの例はすべて機能的な適応ですが、鎌状赤血球症などのヘモグロビン関連疾患、乳糖不耐症、葉酸およびビタミンD欠乏症、多血症などの健康問題を引き起こす可能性もあることに注意してください。これらの例は、還元主義的な説明の限界を示しており、健康問題を体細胞、心理的、社会的なレベルで同時に捉える必要性を正当化し、そのような健康問題は進化論のレンズを通して見た場合に、よりよく理解されることを強調しています。
2.3.3 多層的なミスマッチ
第1章で議論したように、私たちの遺伝子、体、脳は主に狩猟採集民の生活の中で進化しましたが、今日では現代社会の新しい環境に生まれており、この「ミスマッチ」は健康に悪影響を与える可能性があります。表2.2は、人間の進化の歴史の中で存在しなかったか、まれであった新しい生活条件が、複数の領域にわたる影響を与える例を示しています。社会的または心理的要因が体細胞の障害を引き起こす可能性があり、その逆もまた同様です。このミスマッチを認識することで、還元主義的な分析の必要性を正当化しながら、さまざまな有害な健康転帰について、生物医学またはBPSモデルのいずれの範囲を超える説明の深さが可能になります。
表2.2 BPS領域におけるミスマッチとその結果の例
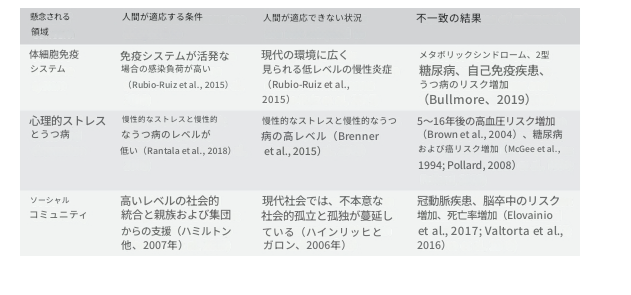
2.4 現在の障害カテゴリーの問題点
現在の精神医学のパラダイム全体で、単純化しすぎることが問題となっています。『精神障害の診断と統計マニュアル』(DSM)は、臨床精神医学の実践において診断システムを作成、検証、研究、および使用するための記述主義的なシステムです。無理論的であろうと試みられていますが、精神障害の性質に関する重要な前提を抱えています。それは、区別しラベル付けする精神障害が明確な症候群であると規定しています。これらの症候群を、定期的に同時に発生する症状のクラスターとして定式化することで、統一された病理学的プロセスを持つ同質な患者グループが研究され、特定の精神療法と薬物療法が開発されることが期待されました。しかし、精神疾患は単純なカテゴリー的または症候群的な構造に従いません(Zachar and Kendler, 2017)。症状は複数の障害間で共有され、健康な集団では亜臨床的な形で現れ、同じ診断を受けた個人の間だけでなく、時間経過に伴う個人の内部でも、その性質と重症度が異なります。脳はDSMシステムの明確なカテゴリー内で機能したり、機能不全になったりしません(Schwartz, 2013)。しかし、DSMは精神科医に対し、一般医学と同様に、患者のカテゴリー診断に基づいて治療することを推奨しています。このアプローチは、各統合失調症患者または各うつ病患者が症状、心理、社会的つながり、および基礎となる生物学的脳機能において同一である場合のように、同質な集団では理にかなっています。精神疾患はこのように同質ではありません。
エンゲルのBPSモデルは、生物医学的アプローチよりも個別化された治療を奨励しました。なぜなら、心理社会的レベルでは個人の違いが明らかだからです。同じ障害カテゴリー内の個人の体細胞の違いは間違いなく存在しますが、通常の精神医学的評価では特定できません。BPSモデルの進化的な進歩は、BPSのすべてのレベルにわたる進化したシステムの固有の複雑さ、相互作用、および文脈依存性の認識を正当化することにより、ここで実際的な意味を持ちます。精神障害は、DSM型の記述主義的分類に適した単純な病気ではありません。将来的には、進化的に知識のある精神科医は、遺伝子型、内表現型、および心理社会的な所見を利用して、初期の精神薬理学的治療計画と管理を調整し、適切な神経経路をより直接的に標的とし、内表現型を正常化し、ライフスタイルの変化を提案し、症状の解消を促進することができるはずです。「症状を駆動する回路」という理論的アプローチは、DSMで検証された症候群的な診断アプローチよりも、実際には脳機能により近いかもしれません。
2.5 機能、機能不全、そして有害な機能不全の概念
最後に、進化医学と進化精神医学の独自の理論的強みに注目します。これは、医療の指導的パラダイムとしての適合性を正当化するものです。病気と障害の概念は、想定される生物学的機能不全と不可分に結びついています。この意味で、健康のBPSモデルは、機能不全を回復させるか、機能不全の症状を緩和することによって、生物学的機能不全を特定し治療するためのモデルです。進化論的視点は、機能と機能不全の概念を進化論に根拠づけることによって、BPSモデルを進歩させることができます。
一般的に、「機能的」と「機能不全」という用語は、あいまいな定義で使用されています。「機能的」は、正常、作動中、または良好な状態を広く指し、「機能不全」は、異常、混乱した、または悪い状態を指します。これらは、真に客観的な基準ではなく、規範によって定義されます。この定義の曖昧さは、医学と精神医学にも存在します(Boyd, 2000)。「病気」、「疾患」、「不健康」、「状態」、「障害」、「機能不全」、「病理」、「病」という用語は、明確な定義や区別なしに、私たちが悪いと見なし、医学的に関連性がある生物学的状態に対して、本質的に交換可能に使用されています(ただし、間違いなく異なる含意を持っています。ボックス2.2を参照)。しかし、生物学的機能は進化的に定義することができ、医学用語に大きな明確さを与えます。
----
ボックス2.2 医学と精神医学における現在の用語の含意
疾患:客観的な生物学的異常、機能不全、または生物の状態。医学では、体細胞的および病態生理学的に理解される。
障害:規範的な機能不全。精神医学では、異常な思考、感情、行動、および他者との関係を伴う心理社会的問題の組み合わせによって特徴づけられると理解される。
病気:主観的な経験、特に苦痛。疾患と障害の症候性兆候を含み、現象学的に理解される。
状態:特定の存在状態。医学において名詞として使用される場合、疾患、診断、障害、病気などを含む可能性のある否定的な状況または苦境として理解される。
障害:人の機能を制限する永続的な身体的または精神的状態。
健康:生物の全体的な幸福の状態。規範的な機能不全、役に立たない主観的な苦痛、または体細胞の不調がなく、最適なパフォーマンスを可能にする、現在の環境における適切な機能として理解される。
----
厳密には機能しているとも機能不全であるとも言えない物理学や化学の対象やプロセスとは異なり(Bolton and Gillett, 2019)、生物学的システムは自然選択による進化の産物であり、したがってそれらの「機能」と「機能不全」はその進化プロセスに関連付けることができます。進化的な機能の概念は、遺伝子が伝播される原因となった表現型によってもたらされる生殖成功に関連しています。目は見るために進化しました。目の機能は見ることです。したがって、機能不全の目は見ることができない目です。この機能の感覚は、視力の進化の歴史が生殖成功をもたらしたことによって定義され、健康と障害の一般的な概念がしばしば依存する正常と異常に関する現在の文化的意見によって定義されるのではありません(Fabrega and Brüne, 2017)。重要な事実は、現代の評価ではなく、進化の歴史です。これにより、進化論は機能不全、そしてひいては医学用語を客観的に定義する上で、他に類を見ないほど理想的になります。
ジェローム・ウェイクフィールドの、疾患と障害に関する影響力のある「有害な機能不全」モデル(Wakefield, 1992, 1997, 2015)は、この進化的な枠組みを利用し、真の障害を2つの構成要素を持つハイブリッド概念として定義しています。最初の構成要素は機能不全の存在であり、システムの進化した機能を実行できないという生物学的基準に基づいています。2番目の非生物学的基準は、機能不全が、一般的な社会文化的基準によって判断されるように、個人に害を及ぼすことです。したがって、私たちの祖先にとって適応的な目的を果たさず、生殖成功を減少させたうつ病の形態は真の障害ですが、(通常の気分の落ち込みに近い)抑うつ状態は、適応的な目的(例えば、社会的敗北後の危険な敵対的行動を防ぐこと。Gilbert and Allan, 1998)を果たしましたが、現代環境では有害であると見なされますが、機能不全ではないため、真の障害として分類されません。
有害な機能不全モデルの理論的な一貫性と、文化的先行詞ではなく明示的な生物学的先行詞への依存にもかかわらず、それ自身の実際的な困難を抱えています。最も重要な科学的困難は、精神障害に関与する神経生物学的システムの大部分の機能が依然として十分に理解されておらず、私たちの進化の歴史が過去によって曖昧にされているため、認知機能の科学的評価が困難であることです。2番目は、医学的治療の適切な適用に関連するより実際的な困難です。苦痛、衰弱、精神的な痛みは、機能的なシステムの特性として、または見慣れないミスマッチ環境で不適応な結果を生み出す機能的なシステムから生じる可能性があります(Del Giudice, 2018; Nesse, 2019)。真の生物学的機能不全がない場合でも、苦痛を伴うさまざまな状況で医学的および精神医学的介入が正当に提供されます。そのような状況は、障害ではなく「精神医学的関心の状態」(Fabrega and Brüne, 2017)とラベル付けされています。精神医学と医学で使用される用語が進化論によって十分に明確化される前に、これらの2つの問題にはかなりの理論的研究が必要です。
この懸念は単なる用語上の問題ではありません。システムの機能状態を正しく特定することは、必要な介入の種類と状態の予後に重要な影響を与える可能性があります。特定の形態のうつ病が、認識可能な適応的進化の歴史を持つ最近の生活上の出来事に対する正常な反応である場合(例えば、地位の喪失後。Sloman et al., 2003)、異なる心理社会的介入が推奨されるかもしれません。臨床および研究の現場で、進化論的な線に沿った用語を適用すること(例えば、真に機能不全のうつ病を「病気」、機能的だが有害なうつ病を「障害」と呼ぶこと)は有用かもしれません。究極の原因を説明する進化論的視点は、「うつ病」のような広範で異質なラベルのより意味のあるサブタイピングを可能にします(Rantala et al., 2018; この巻の第8章も参照)。この客観的な「機能不全」の基準を導入することは、文化的な態度が変化するまで同性愛を精神障害としてラベル付けし、その後投票によってDSMから削除するなど(Drescher, 2015)、最近の歴史の明白に非科学的な慣行から精神医学を遠ざけることにもなります。
2.6 結論
2002年から2015年まで国立精神衛生研究所を率いたアメリカの神経科学者であり生物学的精神科医であるトーマス・インセルは、精神保健研究の現状について、「過去50年間やってきたことは、うまくいっていない…自殺者数、障害者数、死亡率のデータを見ると、それはひどく、改善されていない。おそらく、このアプローチ全体を再考する必要があるだろう…1980年以来、検証されたバイオマーカーがなく、新しい医療処置もほとんどない…精神障害を再考する時が来た」と述べた(Greenberg, 2013)。私たちは、精神障害を再考すること、そして実際には医学全般をより生産的な道(さらにはパラダイム)に乗せるためには、すべての生物学の基本理論である進化論を統合する必要があると信じています。
BPSモデルは、医学と精神医学が還元主義的な生物医学モデルを超えることを奨励する重要な一歩でしたが、十分ではありませんでした。進化精神医学は、主流の生物学的精神医学の、精神障害を単に脳障害と見なす狭い「脱文脈化された」見解(Andreasen, 1984)とは対照的に、生物学的概念を心理的、社会的、文化的領域を包含するように拡大するための科学的根拠を提供します(Abed and St John-Smith, 2016)。進化論的に知識のある研究者は、あらゆる特定の状態に関連する複数のレベルを認識し、そのような要因の影響を改善するための変化を予測、理解、提案し始めることができます。ヘルスケアを進化論に根拠づけることは、BPSモデルの重要な考慮事項に、強固な科学的および哲学的にも首尾一貫した基盤を与えます。BPSモデルの進化的な進歩は、単に規範的ではない機能不全の定義を可能にし、病気の原因を理解する上で新しいミスマッチ環境を認識するために医学研究を開き、人間がうまく適応できていない現代環境の特定の側面を理解し軽減することを奨励するなど、いくつかの方法で治療と予防の努力を方向付けます。理論的および実際的な潜在的な利点のより多くの例は、この巻のさまざまな章で見つかるでしょう。
エンゲルは、実践者のための行動計画としてBPSモデルを提案しました。理論的な一貫性は彼の目的ではありませんでした。彼の業績は賞賛に値しますが、この盲点は、生物医学的アプローチが不均衡な支配を維持している理由である可能性が高いです。心理社会的な視点からなされた誤った主張は、BPSモデルの科学的な魅力を低下させ、哲学的な再評価も役立ちませんでした。私たちは、進化論的な再概念化は非常に科学的に魅力的であり、医学と精神医学を生物学理論に組み込むと主張します。生物医学の実践者は脳のプロセスを優先し、患者の独自の心理的または社会的状態をほとんど重視しないかもしれませんが、BPSの実践者は心理社会的な側面を認識しますが、深い説明的な知識はありません。進化論はそのより深い理解を提供します。一つの直接的な結果は、治療関係を強化し、認知行動療法において役割を果たすことかもしれません。最近の出版物では、「ICT」(情報に基づいた認知療法)というラベルの下で、この応用のための初期のガイドラインが提供されています(Abrams, 2020)。
ダーウィンが『種の起源』初版の結論で書いたように、「遠い未来には、はるかに重要な研究のための開かれた分野が見える」と(Darwin, 1859)。進化科学は、複雑な健康問題を理解し、特に遺伝学、エピジェネティクス、発達可塑性の分野における継続的な研究、そしてこの巻に見られる進化医学と進化精神医学からの基礎的な理論的研究に関連して、新しい解決策を刺激する上で非常に重要です。これは、医学の分野における深刻な科学的進歩であり、その歴史のほとんどは理論よりも実践に関心を持っていました。フレイジャー(2020)が述べているように、BPSモデルは正確にはクーン的なパラダイムシフトというよりも、医学のより包括的な見方への移行でした。しかし、進化医学と進化精神医学への移行は、より妥当にパラダイムシフトと呼ばれる可能性があり、優れた理論的枠組みの中で健康問題を説明し、医学の実践を生物学の科学と結びつけます。
References
Abbott, D. H. et al. (2003) ‘Are subordinates always stressed? A comparative analysis of rank differences in
cortisol levels among primates’, Hormones and Behavior, 43, pp. 67-82.
Abed, R. and St John-Smith, P. (2016) ‘Evolutionary psychiatry: a new College special interest group’,
BJPsych Bulletin, 40, pp. 233-236.
Abrams, M. (2020) The New CBT: Evolutionary Clinical Psychology. San Diego, CA: Cognella Press.
Adler, R. H. (2009) ‘Engel’s biopsychosocial model is still relevant today’,Journal of Psychosomatic Research,
67, pp. 607-611.
Alami, S. et al. (2020) ‘Mother’s social status is associated with child health in a horticulturalist popula-
tion’, Proceedings of the Royal Society B: Biological Sciences, 287, p. 20192783.
Albott, C. S., Forbes, M. K. and Anker, J. J. (2018) ‘Association of childhood adversity with differential sus-
ceptibility of transdiagnostic psychopathology to environmental stress in adulthood’,JAMA Network Open,
1, p. e185354.
Alonso, Y. (2004) ‘The biopsychosocial model in medical research: the evolution of the health concept over
the last two decades’, Patient Education and Counseling, 53, pp. 239-244.
Álvarez, A. S., Pagani, M. and Meucci, P. (2012) ‘The clinical application of the biopsychosocial model in
mental health: a research critique’, American Journal of Physical Medicine and Rehabilitation, 91, pp. S173-
S180.
Andreasen, N. C. (1984) The Broken Brain: The Biological Revolution in Psychiatry. New York: Harper & Row.
Available at: https://cmc.marmot.org/Record/.b11138543 (accessed 10 May 2019).
Andrews, P. W. et al. (2015) ‘Is serotonin an upper or a downer? The evolution of the serotonergic system
and its role in depression and the antidepressant response’, Neuroscience and Biobehavioral Reviews, 51, pp.
164-188.
Assary, E. et al. (2018) ‘Gene-environment interaction and psychiatric disorders: review and future direc-
tions’, Seminars in Cell and Developmental Biology, 77, pp. 133-143.
Barabási, A. L. and Oltvai, Z. N. (2004) ‘Network biology: understanding the cell’s functional organization’,
Nature Reviews Genetics, 5, pp. 101-113.
Beall, C. M. (2007) ‘Two routes to functional adaptation: Tibetan and Andean high-altitude natives’,
Proceedings of the National Academy of Sciences of the United States of America, 104, pp. 8655-8660.
Benedetti, F. (2008) ‘Mechanisms of placebo and placebo-related effects across diseases and treatments’,
Annual Review of Pharmacology and Toxicology, 48, pp. 33-60.
Benning, T. (2015) ‘Limitations of the biopsychosocial model in psychiatry’, Advances in Medical Education
and Practice, 6, p. 347.
Bigham, A. et al. (2010) ‘Identifying signatures of natural selection in Tibetan and Andean populations us-
ing dense genome scan data’, PLoS Genetics, 6, p. e1001116.
Boehm, C. (2012) ‘Costs and benefits in hunter-gatherer punishment’, Behavioral and Brain Sciences, 35,
pp. 19-20.
Bolton, D. and Gillett, G. (2019) The Biopsychosocial Model of Health and Disease: New Philosophical and
Scientific Developments. London: Palgrave Macmillian.
Borell-Carrió, F., Suchman, A. L. and Epstein, R. M. (2004) ‘The biopsychosocial model 25 years later: prin-
ciples, practice, and scientific inquiry’, Annals of Family Medicine, 2, pp. 576-582.
Borsboom, D., Cramer, A. O. J. and Kalis, A. (2019) ‘Brain disorders? Not really: why network structures
block reductionism in psychopathology research’, Behavioral and Brain Sciences, 42, pp. 1-54.
Boyd, K. M. (2000) ‘Disease, illness, sickness, health, healing and wholeness: exploring some elusive con-
cepts’, Medical Humanities, 26, pp. 9-17.
Bracken, P. et al. (2012) ‘Psychiatry beyond the current paradigm’, British Journal of Psychiatry, 201, pp.
430-434.
Braveman, P. and Gottlieb, L. (2014) ‘The social determinants of health: it’s time to consider the causes of
the causes’, Public Health Reports, 129, pp. 19-31.
Brenner, S. L. et al. (2015) ‘Evolutionary mismatch and chronic psychological stress’,Journal of
Evolutionary Medicine, 3, p. 11.
Brown, E. S., Varghese, F. P. and McEwen, B. S. (2004) ‘Association of depression with medical illness: does
cortisol play a role?’, Biological Psychiatry, 55, pp. 1-9.
Bullmore, E. T. (2019) The Inflamed Mind: A Radical New Approach to Depression. London: Short Books Ltd.
Bynum, W. F. et al. (2006) The Western Medical Tradition: 1800 to 2000. Cambridge: Cambridge University
Press.
Campbell, M. C. and Ranciaro, A. (2021) ‘Human adaptation, demography and cattle domestication: an
overview of the complexity of lactase persistence in Africa’, Human Molecular Genetics, 30, pp. 98-109.
Chagnon, N. A. (2013) Noble Savages: My Life among Two Dangerous Tribes – The Yanamamo and the
Anthropologists. New York: Simon & Schuster.
Cohen, S., Murphy, M. L. M. and Prather, A. A. (2019) ‘Ten surprising facts about stressful life events and
disease risk’, Annual Review of Psychology, 70, pp. 577-597.
Colloca, L. and Benedetti, F. (2005) ‘Placebos and painkillers: is mind as real as matter?’, Nature Reviews
Neuroscience, 6, pp. 545-552.
Conrad, P. and Kern, R. (1981) The Sociology of Health and Illness: Critical Perspectives. New York: St.
Martin’s Press.
Darwin, C. (1859) On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured
Races in the Struggle for Life. Available at: http://darwin-online.org.uk/Variorum/1866/1866-576-c-
1859.html (accessed 10 August 2019).
Deacon, B. J. (2013) ‘The biomedical model of mental disorder: a critical analysis of its validity, utility, and
effects on psychotherapy research’, Clinical Psychology Review, 33, pp. 846-861.
Del Giudice, M. (2018) Evolutionary Psychopathology: A Unified Approach. New York: Oxford University
Press.
Doyal, L. (1979) The Political Economy of Health. London: Pluto Press.
Drescher, J. (2015) ‘Out of DSM: depathologizing homosexuality’, Behavioral Sciences, 5, pp. 565-575.
Elovainio, M. et al. (2017) ‘Contribution of risk factors to excess mortality in isolated and lonely individu-
als: an analysis of data from the UK Biobank cohort study’, Lancet Public Health, 2, pp. e260-e266.
Engel, G. L. (1977) ‘The need for a new medical model: a challenge for biomedicine’, Science, 196, pp. 129-
136.
Engel, G. L. (1980) ‘The clinical application of the biopsychosocial model’, American Journal of Psychiatry,
137, pp. 535-544.
Evers, A. W. M. et al. (2014) ‘Incorporating biopsychosocial characteristics into personalized healthcare: a
clinical approach’, Psychotherapy and Psychosomatics, 83, pp. 148-157.
Fabrega, H. and Brüne, M. (2017) ‘Evolutionary foundations of psychiatric compared to nonpsychiatric
disorders’, in T. K. Shackelford and V. Zeigler-Hill (eds.), The Evolution of Psychopathology. Cham: Springer,
pp. 1-35.
Finniss, D. G. and Benedetti, F. (2005) ‘Mechanisms of the placebo response and their impact on clinical tri-
als and clinical practice’, Pain, 114, pp. 3-6.
Frazier, L. D. (2020) ‘The past, present, and future of the biopsychosocial model: a review of The
Biopsychosocial Model of Health and Disease: New Philosophical and Scientific Developments by Derek Bolton
and Grant Gillett’, New Ideas in Psychology, 57, p. 100755.
Ghaemi, S. N. (2009) ‘The rise and fall of the biopsychosocial model’, British Journal of Psychiatry, 195, pp.
3-4.
Ghaemi, S. N. (2010) The Rise and Fall of the Biopsychosocial Model: Reconciling Art and Science in Psychiatry.
Baltimore, MD: Johns Hopkins University Press.
Gilbert, P. and Allan, S. (1998) ‘The role of defeat and entrapment (arrested flight) in depression: an explo-
ration of an evolutionary view’, Psychological Medicine, 28, pp. 585-598.
Greenberg, G. (2013) ‘The Rats of N.I.M.H.’, The New Yorker. Available at:
www.newyorker.com/tech/annals-of-technology/the-rats-of-n-i-m-h (accessed 15 June 2020).
Hamilton, M. J. et al. (2007) ‘The complex structure of hunter-gatherer social networks’, Proceedings of the
Royal Society B: Biological Sciences, 27, pp. 2195-2203.
Harrell, C. J. P. et al. (2011) ‘Multiple pathways linking racism to health outcomes’, Du Bois Review, 8, pp.
143-157.
Heinrich, L. M. and Gullone, E. (2006) ‘The clinical significance of loneliness: A literature review’, Clinical
Psychology Review, 26, pp. 695-718.
Hider, J. L. et al. (2013) ‘Exploring signatures of positive selection in pigmentation candidate genes in pop-
ulations of East Asian ancestry’, BMC Evolutionary Biology, 13, p. 150.
Hoke, M. K. and McDade, T. (2014) ‘Biosocial inheritance: a framework for the study of the intergenera-
tional transmission of health disparities’, Annals of Anthropological Practice, 38, pp. 187-213.
Janko, M. M. et al. (2018) ‘The links between agriculture, Anopheles mosquitoes, and malaria risk in chil-
dren younger than 5 years in the Democratic Republic of the Congo: a population-based, cross-sectional,
spatial study’, Lancet Planetary Health, 2, pp. e74-e82.
Kelly, M. P. et al. (2009) ‘A conceptual framework for public health: NICE’s emerging approach’, Public
Health, 123, pp. e14-e20.
Krakauer, J. W. et al. (2017) ‘Neuroscience needs behavior: correcting a reductionist bias’, Neuron, 93, pp.
480-490.
Krieger, N. (1994) ‘Epidemiology and the web of causation: has anyone seen the spider?’, Social Science and
Medicine, 39, pp. 887-903.
Krieger, N. (2001) ‘A glossary for social epidemiology’,Journal of Epidemiology and Community Health, 55,
pp. 693-700.
Lea, A. J. et al. (2017) ‘Developmental plasticity: bridging research in evolution and human health’,
Evolution, Medicine, and Public Health, 2017, pp. 162-175.
Linden, D. E. J. (2012) ‘The challenges and promise of neuroimaging in psychiatry’, Neuron, 73, pp. 8-22.
Ludwig, J. et al. (2011) ‘Neighborhoods, obesity, and diabetes – a randomized social experiment’, New
England Journal of Medicine, 365, pp. 1509-1519.
Ludwig, J. et al. (2012) ‘Neighborhood effects on the long-term well-being of low-income adults’, Science,
337, pp. 1505-1510.
Mai, F. M. (1995) ‘Clinical and basic science aspects of the biopsychosocial model’,Journal of Psychiatry &
Neuroscience, 20, pp. 335-336.
Marmot, M. (2005) ‘Social determinants of health inequalities’, Lancet, 365, pp. 1099-1104.
Marmot, M. (2006) ‘Status syndrome: a challenge to medicine’,Journal of the American Medical Association,
295, pp. 1304-1307.
Marmot, M. and Wilkinson, R. (eds.) (1999) Social Determinants of Health. Oxford: Oxford University Press.
Mayr, E. (1961) ‘Cause and effect in biology’, Science, 134, pp. 1501-1506.
McElroy, A. (1990) ‘Biocultural models in studies of human health and adaptation’, Medical Anthropology
Quarterly, 4, pp. 243-265.
McGee, R., Williams, S. and Elwood, M. (1994) ‘Depression and the development of cancer: a meta-analy-
sis’, Social Science and Medicine, 38, pp. 187-192.
McLaren, N. (1998) ‘A critical review of the biopsychosocial model’, Australian & New Zealand Journal of
Psychiatry, 32, pp. 86-92.
McQueen, D. et al. (2013) ‘Rethinking placebo in psychiatry: how and why placebo effects occur’, Advances
in Psychiatric Treatment, 19, pp. 171-180.
