
第一章の要約。
第1章の要約:進化心理学、遺伝学と精神病理
- 進化心理学の原理:
- 人間の心は生物の形態的特徴と同様に、自然選択と性選択によって進化した。
- 新生児の心は「タブラ・ラサ(白紙)」ではなく、生得的な心理メカニズムを持つ。
- これらのメカニズムは、愛着形成、特定の形状への好み、特定の恐怖、模倣能力など、生存と繁殖に有利な適応である。
- 心理的メカニズムは生得的な基盤を持つが、経験による修正(「開かれたプログラム」)も重要であり、「生得性」と「育ち」の相互作用が不可欠。
- 進化した心理的メカニズムと生物社会的目標:
- 心理的メカニズムは、適応上の重要な問題を解決するために進化によって設計された。
- これらは領域特異的に機能することが多いが、複数の情報源を統合する領域一般メカニズムも存在する可能性。
- 人間の脳機能は、抽象的な問題よりも社会的な問題の解決に偏っていることが多い。
- 進化した心理的メカニズムの実際の標的は、生物社会的目标の追求(他者からのケア、配偶者の発見、協力、社会的地位の獲得など)。
- これらの目標達成は繁殖の成功の可能性を高めるが、意識的な適応度最大化を目指すわけではない。
- 男性と女性では、祖先時代の異なる役割と繁殖上の制約から、目標の優先順位が異なる。
- 進化した心理的メカニズムは最適設計ではなく、設計上の妥協を含むことが多い。
- 進化的適応環境(EEA):
- 人間の主要な心理的メカニズムは、更新世の狩猟採集生活という遠い過去の環境条件下で進化した(ボウルビーの提唱)。
- 祖先の人類は、小さく、血縁に基づいた、比較的平等主義的な部族集団で生活し、協力が不可欠だった。
- 単一のEEAシナリオは狭すぎるとの批判もあり、更新世以前の適応や、人類がアフリカを出てからの地域的な適応も考慮する必要がある。
- EEAは、進化する人間に選択圧を生み出すのに十分な長さ続いたさまざまな側面の統計的な複合体として捉えるのが適切。
- 現代人と初期の解剖学的現代人は完全に同一ではなく、最近の遺伝子変異も進化に関与している可能性。
- 人間の認知、感情、行動の進化的仮説の検証:
- 進化心理学の枠組みは、検証可能な予測を生み出す理論を提供する。
- 検証は、トップダウン(理論から仮説へ)とボトムアップ(観察から仮説へ)のアプローチがある。
- 証拠源として、実験、質問票、アーカイブデータ、異文化比較、種間比較、精神病理学的兆候などが用いられる。
- 行動の病理は、正常な機能を理解する上で重要な情報源となる可能性がある(ローレンツの主張)。
- 精神病理学におけるこの視点は、これまで十分に活用されてこなかった。
- 遺伝学:
- 生物の成長に必要な情報はゲノムにDNAとして保存され、4種類の塩基の配列(遺伝コード)によってコード化される。
- 遺伝コードはすべての生物で共通であり、高度に保存されている。
- ヒトゲノムは約35億塩基対で構成され、コーディングDNAとノンコーディングDNAを含む。
- 遺伝子はタンパク質の設計図であり、DNAからmRNAへの転写と翻訳を経てタンパク質が合成される。
- ヒトは23対46本の染色体を持つ。遺伝子の変異(対立遺伝子)が進化の源となる。
- 染色体異常(異数性、構造異常)は、精神遅滞や行動異常と関連する可能性がある。
- ヒトのDNAは99%以上が同一だが、SNPやCNVなどの遺伝子多型が存在し、表現型の多様性に寄与する。
- 遺伝子発現はエピジェネティックなメカニズム(DNAメチル化など)によって調節される。
- 遺伝子重複や遺伝子流動、遺伝的浮動も進化に影響を与える。
- 精神病理への遺伝的寄与の決定:
- SNP、遺伝子相互作用、遺伝子サイレンシング、インプリンティングなどが精神病理にどのように寄与するかは、まだ初期段階の理解。
- 遺伝的要因は精神病理への脆弱性と関連するが、遺伝と環境の影響を分離するのは困難。
- 多くの精神障害はメンデル遺伝に従わず、遺伝子発現も可変的で浸透率も不完全。
- 精神病理学的症候群は量的形質であり、多因子脆弱性閾値モデルによる理解が必要。
- 遺伝率(集団レベルでの遺伝的変動による表現型分散の割合)の概念。
- 家族研究、双生児研究、養子研究による遺伝的寄与の推定。
- 遺伝子-環境相互作用と相関(受動的、能動的)の複雑さ。
- 連鎖解析と関連解析による疾患関連遺伝子の探索。
- 複雑な精神障害の定量的遺伝学の課題(多遺伝子性、民族差、診断基準の曖昧さ)。
- 内表現型(遺伝的に単純な生物学的・行動学的マーカー)の重要性。
- 動物モデルを用いた研究の可能性。
- 精神障害の高い有病率を進化論的に説明する試み(バランス選択、頻度依存性選択など)。
- 単一の「病気の遺伝子」の存在は否定的であり、遺伝子と環境の相互作用が重要。
- 精神病理の理解には、進化論的な視点(近接要因と究極要因)が不可欠。
第1章 進化論、進化心理学、遺伝学の原理
1. はじめに
従来、精神障害や精神病理学的な兆候や症状の原因は、胎児期の発達における障害、乳幼児期や青年期の逆境的な出来事、そして神経生物学的なレベルでは、神経回路の機能不全、神経伝達の不均衡、または遺伝的な脆弱性といった観点から概念化されてきました。例えば、うつ病は、愛着のある人物との分離などの乳幼児期の逆境的な出来事の結果、セロトニン欠乏症候群、あるいはセロトニントランスポーター遺伝子の対立遺伝子変異と関連付けられることがあります。心理的機能不全の行動学的および神経生物学的根源の研究が、精神病理学の理解を大きく進歩させたことは疑いありません。しかし、より広範な生物学的視点から見ると、これは物語の半分に過ぎません。
精神医学における一般的な誤解は、個人の初期の発達状態と、脳機能の神経化学的または分子レベルの理解が、認知-感情-行動システムの病理を理解するのに十分であると想定することです。これは、個体発生と生理学だけでは動物の行動の性質を理解するのに十分であると考えるのと同じくらい誤っています。昨世紀の偉大な動物行動学者の一人であるニコラス・ティンバーゲンは、これらのレベルを行動の近接的または直接的な原因と呼びました。行動の近接的な原因は、個体発生的な発達と生理学的メカニズムを含みます。行動(および病理)の近接的な原因は、個人の生涯にわたって変化する可能性があります。
しかし、ティンバーゲンが指摘したように、行動を完全に理解するためには、行動の究極的なレベルも認識することが不可欠です。究極的なレベルは、行動の系統発生的な発達の再構築、つまり、近縁種を注意深く研究することによって行動の進化的ルーツをたどり、特定の形質の選択的有利性または適応的価値を分析する試みを含みます。これら4つの「なぜ」の問い、すなわち個体発生、生理学的メカニズム、系統発生、そして適応的価値は、行動の完全な理解の主要な構成要素を構成します。近接的なレベルと究極的なレベルは決して相互に排他的ではありません。それどころか、それらは本質的に相補的です。
この視点から、人間の認知、感情、行動は、これら4つの問いすべてに取り組むことによってのみ理解でき、不適応な形質も同様の方法で分析する必要があるという明確な理解が、この教科書の主要なテーマを構成します。しかし、これはあらゆる精神病理学的な兆候や症状が適応を表しているという意味ではありません。逆に、兆候や症状は、この用語の一般的な理解と進化的意味の両方において、真に不適応です。
精神病理学的な兆候や症状は、現在の状況における異常な頻度、強度、または不適切さのために機能不全になった、変動の極端な状態を反映しています。例えば、恐怖は、個体に脅威を知らせ、環境の危険を回避するのに役立つ古代からの適応的な形質です。対照的に、病的な不安は、影響を受けた個人が特定の状況を脅威として主観的に認識する可能性があるにもかかわらず、「現実の」脅威をもたらさない状況で発生するため、不適応であると考えられます。恐怖反応は通常容易に引き起こされます。これは、放出閾値を低く保つことが進化的に理にかなっているためです。閾値が高すぎると、恐れを知らない個体はおそらく非常に早く命を落とし、したがって生存する子孫を残さなかったでしょう。したがって、本当に危険なときに一度恐れを知らないよりも、時々不必要に怯える方が「安上がり」なのです。これが、恐怖と不安が非常に多くの精神病理学的状態の一部である理由かもしれません。適応性と不適応性の区別は確かに曖昧な境界を持っています。しかし、うつ病は悲しみではなく、パラノイアは疑い深さではありません。前者は明らかに後者の不適応な極端な状態です。
これらの簡単な例は、私たちの種がどのような状況に適応してきたのか?人間の心を形作った環境条件はどのようなものだったのか?人間の状態の出現の進化的コストは何だったのか?といった問いに答えることがなぜ重要なのかを示唆しているかもしれません。
私たちの種の進化的設計を理解することは、他の医学分野と比較して精神医学において特に必要かもしれません(糖尿病、高血圧、肥満などの「文明病」のより深い理解も、進化論的アプローチから確実に恩恵を受けるでしょう)。言い換えれば、複雑なシステムの機能不全を理解したいのであれば、その機能をあらゆるレベルで理解する必要があります。次の段落では、心理的メカニズムと病的な変異の究極的な原因を理解するために不可欠な基本的な進化論の概念を要約します。
解説補足
- 胎児期の発達(たいじきのはったつ、foetal development): 受精から出生までの期間における、胎児の身体や器官の形成過程。この時期の障害は、後の精神発達に影響を与える可能性があります。
- 乳幼児期(にゅうようじき、infancy): 出生から歩行ができるようになるまでの時期。
- 幼少期(ようしょうき、early childhood): 歩行ができるようになってから小学校入学までの時期。
- 青年期(せいねんき、adolescence): 子供から大人への移行期。
- 神経回路(しんけいかいろ、neural circuits): 脳内の神経細胞が形成するネットワーク。情報の伝達や処理に関わります。
- 神経伝達(しんけいでんたつ、neurotransmission): 神経細胞間で情報が伝達される仕組み。神経伝達物質のバランスの乱れは、精神障害の原因となることがあります。
- 遺伝的脆弱性(いでんてきぜいじゃくせい、genetic vulnerability): 特定の遺伝子変異を持つことで、精神障害を発症しやすくなること。
- 愛着のある人物(あいちゃくのあるじんぶつ、attachment figure): 乳幼児にとって最も重要な養育者であり、安心感や安全感の源となる人物。
- セロトニン欠乏症候群(セロトニンけつぼうしょうこうぐん、serotonin deficiency syndrome): 神経伝達物質であるセロトニンの不足が、うつ病などの精神障害の原因となるという考え方。
- 対立遺伝子変異(たいりついでんしへんい、allelic variation): 同じ遺伝子座に存在する異なる遺伝子の型。
- セロトニントランスポーター遺伝子(セロトニントランスポーターいでんし、serotonin transporter gene): 神経細胞間のセロトニンを再吸収するタンパク質をコードする遺伝子。この遺伝子の変異が、うつ病のリスクに関わることが示唆されています。
- 行動学的(こうどうがくてき、behavioural): 行動に関する。
- 神経生物学的(しんけいせいぶつがくてき、neurobiological): 神経系の生物学的な側面に関する。
- 心理的機能不全(しんりてききのうふぜん、psychological dysfunction): 認知、感情、行動などの心理的な機能が正常に働かない状態。
- より広範な生物学的視点(よりこうはんせいぶつがくてきしってん、broader biological perspective): 生物の進化や適応といった、より包括的な視点。
- 個体発生(こたいはっせい、ontogeny): 個々の生物が受精卵から成体になるまでの発達の過程。
- 生理学(せいりがく、physiology): 生物の体や器官の機能に関する学問。
- 動物行動学(どうぶつこうどうがく、ethology): 動物の自然な環境における行動を研究する学問。ニコラス・ティンバーゲンは、動物行動学の創始者の一人です。
- 近接的な原因(きんせつてきなげんいん、proximate causation): 行動が起こる直接的なメカニズムや、個体の一生における発達的要因。
- 生理学的メカニズム(せいりがくてきメカニズム、physiological mechanisms): ホルモン、神経系など、体の内部で働く仕組み。
- 究極的なレベル(きゅうきょくてきなレベル、ultimate level): 行動が進化の過程でどのように形成され、どのような適応的な意味を持つかという視点。
- 系統発生的な発達(けいとうはっせいてきなはったつ、phylogenetic development): 生物が進化の過程でどのように変化してきたかの歴史。
- 近縁種(きんえんしゅ、closely related species): 進化的に近い関係にある生物種。
- 選択的有利性(せんたくてきゆうりせい、selective advantage): 特定の形質を持つ個体が、持たない個体よりも生存や繁殖において有利であること。
- 適応的価値(てきおうてきかち、adaptive value): 特定の形質が、個体の生存や繁殖に貢献する度合い。
- 相補的(そうほてき、complementary): 互いに補い合い、全体として完全なものにする関係。
- 不適応な形質(ふてきおうなけいしつ、maladaptive traits): 個体の生存や繁殖を妨げる可能性のある形質。
- 変動の極端な状態(へんどうのきょくたんなじょうたい、extremes of variation): 生物集団における形質のばらつきの中で、非常に稀な状態。
- 機能不全(きのうふぜん、dysfunctional): 本来の機能を果たせなくなった状態。
- 異常な頻度、強度、または不適切さ(いじょうなひんど、きょうど、またはふてきせつさ、abnormal frequency, intensity or inappropriateness): 行動や感情などが、通常とは異なる頻度で起こったり、極端に強かったり、状況にそぐわなかったりすること。
- 古代からの適応的な形質(こだいからのてきおうてきなけいしつ、ancient adaptive trait): 進化の初期から存在し、生存に有利に働いてきた形質。
- 病的な不安(びょうてきなふあん、pathological anxiety): 現実の危険がないにもかかわらず、過剰で持続的な不安を感じる状態。
- 放出閾値(ほうしゅついきち、releasing threshold): 特定の行動を引き起こすための刺激の強さの限界値。
- 進化的に理にかなう(しんかてきにりにかなう、makes evolutionary sense): 進化の過程において、生存や繁殖に有利であったと考えられること。
- 安上がり(やすあがり、cheaper): 生存や繁殖におけるコストが低いこと。
- うつ病(うつびょう、depression): 気分の落ち込みや意欲の低下などが持続する精神障害。
- 悲しみ(かなしみ、sadness): 一時的な感情の一つ。
- パラノイア(パラノイア、paranoia): 根拠のない疑心暗鬼や被害妄想を抱く状態。
- 疑い深さ(うたがいぶかさ、suspiciousness): 他者を疑う傾向。
- 進化的コスト(しんかてきコスト、evolutionary costs): 特定の形質が進化する上で、他の形質に及ぼす負の影響や、エネルギー消費などの負担。
- 文明病(ぶんめいびょう、diseases of civilization): 現代の生活習慣や環境が原因で増加している病気。
- 糖尿病(とうにょうびょう、diabetes): 血糖値を調節するホルモンの異常によって起こる病気。
- 高血圧(こうけつあつ、hypertension): 血管内の圧力が必要以上に高い状態。
- 肥満(ひまん、obesity): 体内に脂肪が過剰に蓄積した状態。
- 複雑なシステム(ふくざつなシステム、complex systems): 多くの要素が相互に作用し合って機能する仕組み。脳や精神機能は複雑なシステムとして捉えられます。
- 究極的な原因(きゅうきょくてきなげんいん、ultimate causes): 行動や形質が進化してきた歴史的背景や適応的な意義。
- 心理的メカニズム(しんりてきメカニズム、psychological mechanisms): 認知、感情、行動などを生み出す心の働き。
- 病的な変異(びょうてきなへんい、pathological variants): 正常な範囲を超えた、病的な状態の形質。
2. 進化の定義
進化とは、直接観察することができない歴史的な過程です。観察から推測する必要がありますが、進化の過程が解剖学や生理学と同様に、人間の認知、感情、行動を形作ってきたことに疑いの余地はありません。共通祖先からの進化は、非常に保守的で、むしろゆっくりとした過程です。例えば、チンパンジーと人間の分子的な類似性は、両者の系統が約500万年から700万年前(mya)に共通の祖先から分岐したことを示唆していますが、酵母と人間でさえ共通の遺伝子をいくつか持っています。
進化は、種内の交配可能な個体群レベルで起こり、個体の遺伝的交代を反映します。進化は非常にゆっくりとしています。なぜなら、何百世代、何千世代にもわたる安定化選択は、集団内の個体間の遺伝的分散を減少させ、それによって集団を最適な遺伝子型に近づける傾向があるからです。しかし、集団内には個体間の遺伝的および表現型的変異が存在し、これは選択が作用する「原材料」を生み出すために絶対に必要です。定義上、繁殖に成功した個体は、現在の環境条件に最もよく適応していると見なされます(誤解を招くように「適者生存」と要約されます)。デオキシリボ核酸(DNA)のコピーエラーである突然変異から生じるほとんどの新しい変異は不利であるため、選択は主に適応度の低い変異を排除する過程(負の選択)として作用します。しかし、新しく発達した形質が個体の繁殖適応度を高め、集団の遺伝子プールに固定される場合、正の選択が起こる可能性があります。
これは、選択が個体の表現型レベルで作用することを示唆しており、より急進的な見解では、選択は個々の遺伝子に作用し、遺伝子同士が競合する可能性があるとされています。選択がどのレベルで作用するかについての議論は、完全には決着していません。しかし、以前の提案とは対照的に、選択が種レベルで起こるという仮定は否定されています。
自然選択による進化は「倹約的な」過程です。これは、適応は設計によって最適なのではなく、適応形質の経済性と信頼性の観点から、利益とコストの間の妥協を表していることを意味します。病理(精神病理を含む)が存在し、選択圧によって除去されなかった理由という問いに取り組む際には、この点を念頭に置くことが重要です(第4章を参照)。
個体の表現型は、その構造と生理機能だけでなく、「拡張された」表現型と呼ばれる認知、感情、行動の構成要素も含むことに注意することが重要です。拡張された表現型は選択の対象となり得ますが、これは人間の特性の研究にとって特に重要です。
群選択は、高度に協力的な種において、群の純粋な適応度の利点がすべての個体の適応度の算術平均を超える場合に起こりうるメカニズムです。人間においては、個体選択と群選択は相補的です。遺伝的に無関係な個体間の協力は、生存と繁殖の観点から群に利益をもたらしました。それはまた、群の規範の違反者である「ただ乗りする人」を検出する能力や、道徳的な処罰といった個人の認知および感情的な適応を形作ってきました。これは、社会環境からの情報を処理することに特化した人間の脳の理解(「社会脳仮説」、第2章の考察を参照)、および欺瞞に対する過度の警戒心(その極端なものは精神医学用語で「迫害妄想」と呼ばれる)などの特定の病理の理解にとって特に重要です。
解説補足
- 進化(しんか、evolution): 生物の形質が世代を超えて変化していく現象。
- 共通祖先(きょうつうそせん、common descent): 現在の複数の種が、過去に共有していた祖先。
- 系統(けいとう、lineage): 進化の過程における、祖先から子孫へのつながり。
- 遺伝子(いでんし、gene): 生物の遺伝情報を担うDNAの特定の領域。
- 種(しゅ、species): 生物分類の基本的な単位で、自然条件下で交配し、繁殖力のある子孫を残せる個体群。
- 個体群(こたいぐん、population): 特定の地域に生息する、同じ種の個体の集まり。
- 遺伝的交代(いでんてきこうたい、genetic turnover): 集団内で、世代交代に伴って遺伝子の構成が変化していくこと。
- 安定化選択(あんていかせんたく、stabilizing selection): 集団内の平均的な形質を持つ個体が、極端な形質を持つ個体よりも生存や繁殖に有利になる選択。
- 個体間分散(こたいかんぶんさん、interindividual variance): 集団内の個体間で、形質にばらつきがあること。
- 最適な遺伝子型(さいてきないでんしがた、optimal genotype): 特定の環境において、最も高い適応度をもたらす遺伝子の組み合わせ。
- 遺伝的変異(いでんてきへんい、genetic variation): 集団内の個体間で、遺伝子構成が異なること。
- 表現型的変異(ひょうげんがたきへんい、phenotypic variation): 集団内の個体間で、外見や生理機能などの表現形が異なること。遺伝的変異と環境要因の相互作用によって生じます。
- 選択(せんたく、selection): 環境に適した形質を持つ個体が、そうでない個体よりも生存や繁殖において有利になり、その形質が次世代に伝わりやすくなる過程。
- 適者生存(てきしゃせいぞん、survival of the fittest): ダーウィンの進化論を要約した言葉としてよく知られていますが、実際には「最も適応したものが生き残る」という意味合いがより正確です。繁殖の成功も重要な要素です。
- 突然変異(とつぜんへんい、mutation): DNAの塩基配列が変化すること。新たな遺伝的変異の主な原因となります。
- デオキシリボ核酸(DNA、ディーエヌエー、deoxyribonucleic acid): 生物の遺伝情報を担う分子。
- 不利な変異(ふりなへんい、disadvantageous mutation): 生存や繁殖を妨げる可能性のある突然変異。
- 負の選択(ふのせんたく、negative selection): 適応度の低い変異を持つ個体が淘汰される選択。
- 正の選択(せいのせんたく、positive selection): 適応度の高い変異を持つ個体が生き残りやすく、その変異が集団内に広まる選択。
- 繁殖適応度(はんしょくてきおうど、reproductive fitness): 個体が次世代に遺伝子を残す能力。生存だけでなく、繁殖の成功も含まれます。
- 遺伝子プール(いでんしプール、genepool): 集団内のすべての個体が持つ遺伝子の総体。
- 表現型(ひょうげんがた、phenotype): 生物の外見、生理機能、行動など、観察可能な形質の総体。遺伝子型と環境要因の相互作用によって決まります。
- 遺伝子レベルの選択(いでんしレベルのせんたく、selection at the level of individual genes): リチャード・ドーキンスが提唱した「利己的な遺伝子」の考え方で、選択の基本的な単位は個体ではなく遺伝子であるとする見解。
- 種レベルの選択(しゅレベルのせんたく、selection at the species level): かつて提唱された、種の存続のために選択が働くと考える説。現在では否定されています。
- 倹約的な過程(けんやくかてきなかっせい、thrifty process): 進化は、過剰なコストをかけずに、必要最低限の適応を生み出す傾向があるということ。
- 適応(てきおう、adaptation): 生物が環境に適応するために獲得した形質や能力。
- 拡張された表現型(かくちょうされたひょうげんがた、extended phenotype): 個体の遺伝子が、その個体の体だけでなく、環境中の構造物や他の生物の行動にも影響を与えるという概念。
- 群選択(ぐんせんたく、group selection): 個体の適応度だけでなく、群全体の適応度が選択に影響を与えるメカニズム。協力的な行動の進化を説明する上で重要となることがあります。
- 純粋な適応度の利点(じゅんすいなてきおうどのりてん、net fitness advantage): 群全体として得られる適応度の利益。
- 算術平均(さんじゅつへいきん、arithmetic mean): 数値の合計を個数で割った平均値。
- 個体選択(こたいせんたく、individual selection): 個々の個体の適応度の違いに基づく選択。
- 協力(きょうりょく、cooperation): 複数の個体が互いに利益をもたらすように行動すること。
- 遺伝的に無関係な個体(いでんてきにむかんけいなこたい、genetically unrelated individuals): 血縁関係のない個体。
- ただ乗りする人(ただのりするひと、free-riders): 群の利益を享受しながら、協力的な行動をとらない個体。
- 道徳的な処罰(どうとくてきなしょばつ、moral punishment): 群の規範に違反した者に対して行われる制裁。
- 社会脳仮説(しゃかいのうかせつ、social brain hypothesis): 霊長類において、集団の規模と脳の大きさ(特に大脳新皮質)が相関しているという仮説。複雑な社会関係に対処するために脳が発達したと考えられています。
- 過度の警戒心(かどのかいけいしん、hypervigilance): 常に周囲の脅威に注意を払い、過敏に反応する状態。
- 欺瞞(ぎまん、deception): 他者を欺くこと。
- 迫害妄想(はくがいもうそう、persecutory delusions): 他者が自分を害しようとしているという根拠のない強い確信。
2.1. 自然選択と性選択
進化の過程は、理論的には自然選択によって生じる形質と、性選択に由来する形質に分けられます。一般的に言って、自然選択は個体の生存に役立つ特性を有利にします。例えば、捕食者から逃れるためには、速く走ることが有利になる(あるいは、保護色を持つことも同様です)。その結果、足の速い個体は、遅い個体よりも平均して生存する子孫を残す可能性が高くなります。捕食する種も同様に選択にさらされます。なぜなら、最も熟練した狩猟者は、最も繁殖する可能性が高いからです。自然界におけるこの競争原理は、おそらくアナゲネシス、つまり進化における複雑さと体サイズの増大傾向が観察される理由の一つでしょう。進化論の研究者たちはこれを、共進化的な「軍拡競争」または「赤の女王の原理」(ルイス・キャロルの『不思議の国のアリス』にちなむ)と呼び、複雑さの増大は少なくとも同じ場所に留まる(そして絶滅しない)ための必要条件であるという比喩としています。
進化的な「軍拡競争」は、種間だけでなく種内でも起こります。例えば、人間においては、遺伝的に無関係な個体間の協力の増加に向けた選択が、「心の理論」、つまり自分自身と他者の心的状態を認知的に表象する能力のような多くの社会認知能力の進化にかなりの圧力をかけた可能性があります(詳細は第2章の考察を参照)。自然選択された形質は、基本的に両性で同じです。対照的に、同じ種の雄と雌の個体間で異なる特性は、性選択の産物です。
チャールズ・ダーウィンが、性選択を第二の(そして少なくとも同等に重要な)進化の力として発見する前は、特に雄において、明らかに生存に不利な形質が存在するという事実に困惑していました。例えば、クジャクの尾は非常に目立ち、重いため、特に最も大きな尾を持つ個体が捕食者から逃れるのを妨げます。しかし、雌が配偶者として優先的に選ぶのは、これらの個体なのです。雄におけるこのような誇張された形態的特徴の進化を説明する可能性のある仮説が二つあります。一つは「遺伝的浮動」と呼ばれ、小さな繁殖集団における対立遺伝子頻度のランダムな変化が、いわゆる暴走選択を通じて特定の特性の頻度を増加させるメカニズムです。暴走選択は、偶然に雌に好まれた形質が、世代を経るごとに雄によるこの形質の過剰発現を引き起こし、それが今度は雌におけるさらに強い好みを引き起こすメカニズムとして理解されています。もう一つは「良い遺伝子仮説」または「ハンディキャップ原理」と呼ばれます。例えばクジャクにおいて、大きな尾は良い遺伝子を示す可能性があります。なぜなら、それを「維持する余裕」のある個体は、病原体に対して抵抗力がある可能性が高いからです。さらに、雄の雌に対する魅力は、対称性も関係しています。「良い遺伝子」は、変動する非対称性の減少と関連していると提唱されています。なぜなら、特に初期の個体発生における疾病の過程は、より大きな非対称性をもたらし、それは魅力的でないと認識されるからです(これは最近、統合失調症に関して提唱されている仮説です。第10章を参照)。
ほとんどの種において、雄は雌へのアクセスを巡って互いに競争します(同性内競争)。したがって、少なくとも一夫多妻の種においては、選択は雌と比較して雄のより大きな体サイズと強さを有利にします。これは性的二形と呼ばれます。縄張り争いにおける同性内競争は、体サイズや強さだけでなく、雄が優位性を確立するための角や大きな犬歯のような武器によっても解決されることがあります。雄が雌に対してほぼ平等なアクセス権を持つ場合、特殊なタイプの同性内競争が起こります。そのような場合、精子の量と質が、受精した雌の子供の父親となる雄を決定する可能性があり、これは同性内競争が起こりうるもう一つのレベルを示しています。一部の個体は、非常に異形に見えるものの、他の精子に対して有毒である可能性のある、いわゆる「カミカゼ精子」さえ生産します(詳細は「親投資と性選択」のセクションを参照)。
異性間の競争もあり、これは異性間競争と呼ばれます。雌の選択の原理はすでに上で述べました。雌が雄よりも配偶者選択において慎重である理由は、ほとんどの種において、雌は雄よりも潜在的な子孫に多くの投資をするからです(「親投資と性選択」のセクションを参照)。言い換えれば、雌は利用可能な最良の配偶者を徹底的に選ばなければ、雄と比較して繁殖適応度の点で失うものがはるかに多いのです。雌の選択の背後にある論理は、繁殖適応度は単なる子孫の数だけでなく、次世代の遺伝的質によってより大きく決定されるということです。したがって、雌にとって遺伝的に質の高い雄を選ぶことは利益になります。多くの鳥類のような一夫一妻の種では、雌はつがい外の交尾に従事することさえあります。一方、一部の種では、雄は強制交尾や配偶者強制によって雌の選択を回避する戦略を進化させてきました。これらのメカニズムは、社会構造や他者との遺伝的関連性に応じて、雄と雌の行動に深刻な影響を与えます。
解説補足
- 自然選択(しぜんせんたく、natural selection): 環境に適した形質を持つ個体が生き残りやすく、子孫を残しやすいという進化のメカニズム。
- 性選択(せいせんたく、sexual selection): 生殖において有利な形質を持つ個体が、異性を獲得しやすく、子孫を残しやすいという進化のメカニズム。
- アナゲネシス(anagenesis): 進化の系統において、祖先の種が絶滅し、子孫の種が祖先の種とは異なる特徴を持つようになる進化の様式。一般的に、複雑化や大型化の傾向が見られます。
- 共進化(きょうしんか、co-evolution): 異なる種の間で、互いに影響を与え合いながら進化する現象。
- 軍拡競争(ぐんかくきょうそう、arms race): 共進化の一種で、捕食者と被食者の間などで、互いに対抗する形質を強化し合う進化。
- 赤の女王の原理(あかのじょおうのげんり、Red Queen Principle): 進化において、現状を維持するためには常に変化し続けなければならないという考え方。ルイス・キャロルの『鏡の国のアリス』に登場する赤の女王のセリフに由来します。
- 心の理論(こころのりろん、theory of mind): 自分自身や他者の心的状態(信念、意図、感情など)を理解し、推測する能力。
- 性的二形(せいてきにけい、sexual dimorphism): 同じ種の雄と雌の間で、体サイズや形態に違いが見られる現象。
- 遺伝的浮動(いでんてきふどう、genetic drift): 小さな集団において、偶然によって対立遺伝子の頻度が変動する現象。
- 暴走選択(ぼうそうせんたく、runaway selection): 特定の形質に対する異性の好みと、その形質を持つことの遺伝的傾向が連鎖し、世代を超えて極端な形質が発達する現象。
- 良い遺伝子仮説(よいいでんしかせつ、good genes hypothesis): 異性が特定の誇張された形質を好むのは、それが遺伝的な質の高さを示す指標であるためという考え方。
- ハンディキャップ原理(ハンディキャップげんり、handicap principle): 生存に不利なはずの誇張された形質が、実際には遺伝的な質の高さを示す正直な信号であるという考え方。維持にコストのかかる形質ほど、質の高い個体しか持つことができないため。
- 病原体(びょうげんたい、pathogen): 病気を引き起こす微生物やウイルスなど。
- 対称性(たいしょうせい、symmetry): 左右の形が均等であること。一般的に、対称性の高い個体は遺伝的に質が高いと考えられています。
- 変動する非対称性(へんどうするひたいしょうせい、fluctuating asymmetry): 本来対称であるべき左右の形が、個体によってわずかに異なること。発達期のストレスなどによって増大すると考えられています。
- 初期の個体発生(しょきのこたいはっせい、early ontogeny): 個体の初期の発達段階。
- 同性内競争(どうせいないきょうそう、intrasexual competition): 同じ性別(通常は雄)の個体間で、異性へのアクセスを巡って起こる競争。
- 縄張り争い(なわばりあらそい、competition over territories): 繁殖や資源を確保するために、縄張りを巡って起こる競争。
- 武器(ぶき、weaponry): 角、牙、爪など、競争において使用される身体的な構造。
- 優位性階層(ゆういせいかいそう、dominance hierarchies): 集団内における個体間の優劣関係。
- 精子の競争(せいしのきょうそう、sperm competition): 一匹の雌が複数の雄と交尾した場合に、受精を巡って精子同士が競争する現象。
- カミカゼ精子(カミカゼせいし、kamikaze sperm): 他の精子を妨害したり、殺したりする機能を持つと考えられる特殊な形態の精子。
- 異性間競争(いせいかんきょうそう、intersexual competition): 異なる性別間(通常は雌による雄の選択)で起こる競争。
- 雌の選択(めすのせんたく、female choice): 繁殖相手の雄を雌が選択すること。
- 親投資(おやとうし、parental investment): 親が子孫の生存と繁殖のために費やす時間、エネルギー、資源など。一般的に、雌の方が親投資の量が多いです。
- 繁殖適応度(はんしょくてきおうど、reproductive fitness): 個体が次世代に遺伝子を残す能力。
- 遺伝的質(いでんてきしつ、genetic quality): 個体が持つ遺伝子の良さ。生存能力や病気への抵抗力などに影響します。
- 一夫一妻制(いちぶいっさいせい、monogamous species): 一組の雄と雌がペアを形成し、共同で子育てを行う繁殖システム。
- つがい外の交尾(つがいそとのこうび、extra-pair copulation): つがいを形成している個体が、つがい相手以外の個体と交尾すること。
- 強制交尾(きょうせいこうび、forced copulation): 雌の意思に反して雄が交尾を強行すること。
- 配偶者強制(はいぐうしゃきょうせい、mate coercion): 雄が雌を脅迫したり、傷つけたりすることで、交尾を強いる行動。
- 社会構造(しゃかいこうぞう、social structure): 集団内における個体間の関係や組織のパターン。
- 遺伝的関連性(いでんてきかんれんせい、genetic relatedness): 共有する遺伝子の割合。血縁関係の近さを示します。
2.2. 包括適応度理論
進化論の現代的統合(メンデルの遺伝学とダーウィンの進化論を統合した理論)は、個体の繁殖成功度が、差のある進化的発達の中心的なテーマであることを明らかにしました。包括適応度理論は、繁殖成功度は個体の産んだ子供の数(古典的適応度)だけに依存するのではなく、遺伝的に関連のある個体の繁殖成功度にも依存すると提唱します。したがって、適応度の観点から見ると、近親者の繁殖努力を支援することが有利になる可能性があります。言い換えれば、包括適応度とは、個体の繁殖成功度と、血縁度によって重み付けされた親族の成功度の合計です(例えば、兄弟姉妹は従兄弟よりも血縁関係が近い)。
選択は、繁殖成功度を最大化する行動を有利にするはずです。なぜなら、遺伝的な観点から見ると、生物が自身の遺伝子を次世代に広めることは利益になるはずだからです。したがって、利他的な行動は血縁関係に限定されるはずであり、実際、利他的な行為は、社会性昆虫や他の社会生活を送る種を含む動物界で広く見られます。しかし、私たちのような社会生活を送る種では、遺伝的に遠い、または無関係な個体間でも相互扶助が見られます。これは、包括適応度理論と一見矛盾するため、説明が必要です。
対立は、遺伝的に無関係な個体間の相互依存の状態だけでなく、異性の個体間でも生じます。なぜなら、彼らの繁殖成功度を最大化するための戦略は根本的に異なる可能性があるからです。遺伝的な非類似性によるこのような対立は、親と子の世代間でも起こります。なぜなら、親は平均して、個々の子供と遺伝物質のわずか50パーセントしか共有していないからです。これらの対立の領域は、ロバート・トリヴァースによって3つの画期的な論文で指摘されています。
解説補足
- 現代的統合(げんだいてきとうごう、modern synthesis): 20世紀半ばに確立した、ダーウィンの自然選択説とメンデルの遺伝学を統合した進化論の包括的な理論。
- 差のある進化的発達(さのあるしんかてきはったつ、differential evolutionary development): 個体間で繁殖成功度に差があることが、進化の方向性を決定する主要な要因であるという考え方。
- 包括適応度理論(ほうかつてきおうどりろん、inclusive fitness theory): 個体の遺伝子が次世代にどれだけ広まるかを評価する際に、その個体自身の繁殖成功度だけでなく、遺伝的に関連のある個体の繁殖成功度も考慮に入れる理論。W.D.ハミルトンによって提唱されました。
- 古典的適応度(こてんてきおうど、classical fitness): 個体が直接生み出した子供の数。
- 血縁度(けつえんど、degree of relatedness): 遺伝的なつながりの強さ。例えば、親子や兄弟姉妹では0.5、祖父母と孫では0.25、従兄弟では0.125となります。
- 利他的な行動(りたてきなこうどう、altruistic behaviour): 自身の利益を損なってでも、他者の利益になるような行動。
- 血縁選択(けつえんせんたく、kin selection): 血縁者への利他的な行動が、自身の遺伝子を間接的に次世代に伝えるのに役立つため、進化的に有利になるという考え方。
- 相互扶助(そうごふじょ、mutual help): 遺伝的に関係のない個体間で行われる協力的な行動。
- 互恵的利他主義(ごけいてきりたしゅぎ、reciprocal altruism): 将来の見返りを期待して、遺伝的に関係のない他者に利益を与える行動。ロバート・トリヴァースによって提唱されました。
- 繁殖戦略(はんしょくせんりゃく、reproductive strategies): 個体が自身の繁殖成功度を最大化するためにとる行動や特性。雄と雌では、卵や精子の生産コストの違いなどから、異なる戦略をとることがあります。
- 遺伝的な非類似性(いでんてきなひるいじせい、genetic dissimilarity): 遺伝子構成が異なること。
- 親子の対立(おやこのたいりつ、parent-offspring conflict): 親が子のために投資する資源の量について、親の最適な戦略と子の最適な戦略が異なるために生じる対立。
2.2.1. 互恵的利他主義
互恵的利他主義は、遺伝的に無関係な個体間の利他主義の一形態として記述できます。それは、ある生物が別の生物の犠牲のもとに利益を提供し、その利他的行為に対する将来の互恵が期待されるものです。食物の共有は、多くの種で見られるそのような行動の一つです。
互恵的利他主義が適切に機能するためには、利他的に行動する個体が、その状況と利益を受けた個体を記憶するための心理的メカニズムを持っている必要があります。さらに、その個体が互恵を拒否した場合に、それを認識できることが不可欠です。欺瞞の検出は、欺瞞的な行動に対抗するために選択されたに違いない認知メカニズムの一つです。なぜなら、そうでなければ、欺瞞の傾向を有利にする遺伝子が集団に広まり、相互扶助のシステムは崩壊していたでしょう。
互恵的利他主義に関連する問題は、祖先(そして間違いなく現代の)人類社会において喫緊の課題でした。なぜなら、交換の機会は、分散率が低く、相互依存度が高く、長寿の種において最も大きいからです。これには、長期間にわたる親による育児も含まれます。したがって、危険な時の相互援助、食物の共有、病気の人、非常に高齢または非常に若い個体の世話、道具や知識の共有は、あらゆる人間の文化で定期的に観察される利他的な行動です。
トリヴァースは、互恵的利他主義の問題に対する解決策として、人間に進化したいくつかの認知的および感情的特性を特定しました。友情、同情、感謝のような肯定的な感情は、利他的な協力関係における感情的な報酬として進化した可能性があります。対照的に、道徳的攻撃性を含む否定的な感情は、欺瞞の傾向を減らし、互恵がない場合の継続的な利他的行動を防ぐ可能性があります。欺瞞の試みが検出された場合、罪悪感や恥のような感情は、互恵的なパートナーシップの回復に役立つかもしれません。認知レベルでは、他者に利他的な行動を誘発するために、道徳的攻撃性、同情、または罪悪感の感情を装うこともありえます。そのような巧妙な欺瞞メカニズムは、正直な行動と偽善的な行動を区別する能力の選択につながった可能性があります。信頼と疑念の感情は、互恵的利他主義の問題に関連して直接進化した可能性があります。さらに、互恵関係を維持することは、数年、あるいは数十年後の個人的に重要な社会的相互作用の想起を可能にする自伝的記憶の進化にも貢献したと考えられます。
群選択はさらに、欺瞞者に対する集団的処罰を有利にした可能性があります。初期の人類集団における個人の並外れて広範な相互依存を考慮すると、群の結束と社会的規則や規範への服従が選択圧の標的となった理由が明らかになります。人間が、グループメンバーの非協力的な行動を罰するために、追加のコストをいとわないという豊富な証拠があります。
精神病理学の観点から見ると、互恵的利他主義の問題に関連して、二つの側面が重要です。一つは、疑い深さの極端な変動、つまり妄想性観念であり、最終的には迫害妄想や関係妄想につながる可能性があります。興味深いことに、迫害妄想の内容はほとんど常に、影響を受けた個人が誰か他の人が悪意を持っている(つまり、自分を欺こうとしている)と確信しているというものです。迫害妄想の内容として、好意的な意図は例外です(詳細は第10章を参照)。もう一つは、巧妙な欺瞞の極端な変動、つまり精神病質です。精神病質は、低有病率レベルの集団で優勢になる可能性のある欺瞞者の形態として解釈されています。現在の概念化では、反社会性パーソナリティ障害との重複があるにもかかわらず、精神病質はそれ自体が病理を表すものではないと考えられています(詳細は第14章を参照)。
解説補足
- 互恵的利他主義(ごけいてきりたしゅぎ、reciprocal altruism): 遺伝的に関係のない個体間で、将来的な見返りを期待して行われる利他的な行動。
- 食物の共有(しょくもつのきょうゆう、food sharing): 獲得した食物を他の個体と分け合う行動。
- 欺瞞(ぎまん、deception): 他者を欺くこと。
- 欺瞞の検出(ぎまんのけんしゅつ、detection of cheating): 他者が利己的な目的で欺こうとしていることを見抜く能力。
- 祖先人類社会(そせんじんるいしゃかい、ancestral human societies): 狩猟採集民など、現代文明以前の人類社会。
- 分散率(ぶんさんりつ、dispersal rates): 個体が生まれた場所から移動する割合。低い分散率は、集団内の個体が長く一緒にいることを意味します。
- 相互依存度(そうごいぞんど、degree of mutual dependence): 個体同士が互いに頼り合って生きている度合い。
- 親による育児(おやによるいくじ、parental care): 親が子どもの世話をすること。
- 相互援助(そうごえんじょ、mutual assistance): 困っている個体を助ける行動。
- 道具や知識の共有(どうぐやちしきのかくゆう、sharing implements and knowledge): 生存に必要な道具や情報を他の個体と分け合う行動。
- 認知的および感情的特性(にんちてきおよびかんじょうてきとくせい、cognitive and emotional characteristics): 思考や感情に関わる特性。
- 友情(ゆうじょう、friendship): 親しい間柄の感情。
- 同情(どうじょう、sympathy): 他者の苦しみや悲しみに共感する感情。
- 感謝(かんしゃ、gratitude): 他者からの恩恵に対して抱く感謝の気持ち。
- 道徳的攻撃性(どうとくてきこうげきせい、moralistic aggression): 規範を破った者や欺瞞的な行為をした者に対して抱く怒りや、それを表明する行動。
- 罪悪感(ざいあくかん、guilt): 自分の過ちや不正な行為に対して抱く後悔の気持ち。
- 恥(はじ、shame): 他者からの評価を意識し、自分の行動や状態を不名誉に感じる気持ち。
- 自伝的記憶(じでんてききおく、autobiographical memory): 個人の経験や出来事に関する記憶。
- 集団的処罰(しゅうだんてきしょばつ、collective punishment): 集団の規範を破った者に対して、集団全体が行う処罰。
- 初期の人類集団(しょきじんるいしゅうだん、early hominid groups): 人類の祖先の初期の集団。
- 群の結束(むれのけっそく、group cohesion): 集団のまとまり。
- 社会的規則や規範への服従(しゃかいてききそくやきはんへのふくじゅう、obedience to social rules and norms): 集団で定められたルールや行動基準に従うこと。
- 選択圧の標的(せんたくあつのひょうてき、targets of selective forces): 進化において、自然選択や性選択の影響を受けやすい特性。
- 非協力的な行動(ひきょうりょくてきなこうどう、non-cooperative behaviour): 集団の利益に反する、または協力しない行動。
- 妄想性観念(もうそうせいかんねん、paranoid ideation): 根拠のない疑念や被害的な考え。
- 迫害妄想(はくがいもうそう、delusions of persecution): 他者が自分を害しようとしているという妄想。
- 関係妄想(かんけいもうそう、delusions of reference): 周囲の出来事や他者の行動が、自分に特別な意味を持っていると誤って信じる妄想。
- 悪意(あくい、malicious intents): 他者を害しようとする意図。
- 好意的な意図(こういてきないと、benevolent intentions): 他者の幸福を願う意図。
- 精神病質(せいしんびょうしつ、psychopathy): 他者への共感性の欠如、衝動性、反社会的な行動などを特徴とするパーソナリティ特性。
- 欺瞞者の形態(ぎまんしゃのけいたい、cheater morph): 集団内で、欺瞞的な行動をとる個体のタイプ。
- 低有病率レベル(ていゆうびょうりつレベル、low prevalence levels): 集団内でその特性を持つ個体の割合が低い状態。
- 反社会性パーソナリティ障害(はんしゃかいせいパーソナリティしょうがい、antisocial personality disorder): 他者の権利を侵害したり、社会のルールを無視したりする行動パターンを特徴とするパーソナリティ障害。
2.2.2. 親投資と性選択
有性生殖は、適切な配偶者を見つけるための時間とエネルギーという点でコストがかかるにもかかわらず存在します。有性生殖はまた、個体の遺伝子のわずか50パーセントしか子孫に伝わらないという事実を考えると、遺伝物質の浪費とも関連しています。それにもかかわらず有性生殖が存在する理由は、遺伝子の組み換えによって有害な突然変異の蓄積の影響を打ち消し、急速に進化する病原菌や寄生虫に対する生物の防御システムを向上させるメカニズムだからです。有性生殖は、遺伝物質の交代が遅い長寿命の種にとって特に重要です。
進化は2種類の配偶子を生み出しました。一つはかなりの代謝資源を必要とする大きな配偶子(卵)、そしてエネルギー的にずっと安価で、はるかに多数生産される配偶子(精子)です。慣例的に、これらの異なる種類の配偶子は「雌」と「雄」と呼ばれます。雄と雌の配偶子の違いは、子孫への親投資の量と繁殖戦略に重要な影響を与えます。
トリヴァースは、親投資を「親による個体の子孫へのあらゆる投資であり、その投資は、親が他の子孫に投資する能力を犠牲にして、その子孫の生存(ひいては繁殖)の可能性を高めるもの」と定義しました。哺乳類では、妊娠、授乳、養育、保護は雌の生物の負担ですが、ほとんどの種では雄は受精以上の投資をほとんど行いません。この予測から、潜在的な子孫に多くの投資をする性(通常は雌)は配偶者選択においてより識別力が強く、投資の少ない性(通常は雄)はより多くの投資をする性へのアクセスを巡って同性内で競争し、配偶者を選ぶ際に識別力が低いと推測できます。その理由は、繁殖はエネルギー、資源、時間という点でコストがかかり、これらの制約は雌にとってより重要であるからです。なぜなら、彼女たちの繁殖成功度は、受精可能な卵の数によって制限されるからです。対照的に、雄の繁殖成功度は(理論的には)アクセス可能な受精可能な雌の数によって制限されます。言い換えれば、子孫の生産に最も投資する性が、もう一方の性が競争する制限された資源となるのです。
霊長類、特に人間においては、繁殖成功度は、非常に長い幼年期、繁殖における高齢、単胎出産、長い出産間隔を含む追加の多くの要因によって制約されます。人間においては、選択は子孫に多大な投資をいとわない雄を有利にしてきました。多くの人間社会で見られる核家族(つまり一夫一妻制)またはせいぜい穏やかな一夫多妻制は、ほとんどの他の霊長類と比較して、人間の雄のより大きな父親としての投資の結果かもしれません。それでも、人間の母親は平均して父親よりも子供に多くの投資をしており、女性が子供を育てる上で配偶者からの支援を得るために、長期的なパートナーシップをいとわず、そのための資源を持つ配偶者を選ぶことは彼女たちの利益になったに違いありません。逆に、親投資理論からすると、人間の男性が複数の女性と交尾することは利益になるはずです。これらの予測と一致して、選択は配偶者防衛、嫉妬の感情、隠蔽排卵、そしておそらく性的強制(「雌の選択」の原則を回避するため)を含む多くの感情的および認知的適応を有利にしてきました。これらの適応は、行動レベルでは霊長類や人間に特有のものではありませんが、主観的な感情の側面はおそらくそうです。嫉妬は男性と女性の両方にとって重要な感情ですが、おそらく異なる理由からです。女性は、配偶者に他の女性のために見捨てられることを恐れる可能性があるため、より感情的な嫉妬をするかもしれません。対照的に、男性は、父親であることの不確実性という問題に直面するため、より性的な嫉妬をすると考えられています。女性は自分が子供の母親であると常に100パーセント確信できますが、父親は同じように確信することはできません。実際、托卵は一夫一妻制の鳥類を含む多くの種で広範囲に見られます。人間では、最大10パーセントの子供が生物学的な父親と遺伝的なつながりを持っていません。実験的に、男性はパートナーとの分離後に、父親であることの不確実性のリスクを補償するために、より多くの精子を生産するように選択されたことが示されています。さらに、陰茎亀頭の解剖学的形状は、自身の精子を注入すると同時に、膣から他の精子を取り除くための適応として解釈されています。これらのメカニズムは、いわゆる「精子競争」、つまり雄間の同性内競争の交尾後の遅延に関連している可能性があります(下記参照)。
しかし、一夫一妻制は、おそらく人間に進化した唯一の可能な配偶戦略ではありません。実験条件下では、排卵期に近い若い女性は(もちろん無意識的に)、対称的な男性の体臭と男らしい男性の顔の特徴を好みますが、実際の配偶者選択は、男性が長期的な関係を追求する意思によって影響を受ける可能性があります。最も近縁な親戚を詳しく見ると、選択が配偶システムにどのように異なる影響を与えてきたかがわかります。例えば、ゴリラは雄と雌の間の体サイズの大きな違いが特徴です。しかし、ゴリラの精巣の重さは、体サイズと比較して小さいです(約30グラム)。これは、単雄複雌と最もよく記述できるゴリラの社会構造に関係しています。対照的に、チンパンジーでは性的二形は比較的小さいですが、雄のチンパンジーは、成体のシルバーバックゴリラの3分の1のサイズしかないにもかかわらず、約120グラムの巨大な精巣を持っています。チンパンジーは多雄多雌のグループで生活し、雌は発情期に複数の雄と交尾するという意味で「乱交的」に行動します。雄間の同性内競争は、雄が雌の膣管に注入できる精子の量の変動(精子競争)によって、交尾後に移行します。雌のチンパンジーは、卵に到達する精子が競争力を持つ(したがって、その生産者の「良い遺伝子」を示す)ことを保証するために、複数の雄からの精子を受け入れることによって、精子競争の過程で積極的な役割を果たす可能性があります。
人間の身体的特徴は異なる物語を語っています。性的二形は比較的小さく(男性は約10〜15パーセント大きい)、精巣は小さく(約40グラム)、そして女性はチンパンジーとは対照的に、排卵を示す目に見える兆候がありません。隠蔽排卵は、雄からの資源の確保(「セックス・フォー・フード」仮説)、雄による子殺しからの子孫の保護、つがい関係の強化など、さまざまな適応上の問題を解決するために進化した可能性があります。雄による子殺しは、霊長類を含む多くの哺乳類で一般的であり、新しい成体の雄がアルファの地位を占めると発生する可能性があります。雄の視点からすると、自分が父親でないすべての子孫を殺すことは理にかなっています。なぜなら、雌ははるかに早く再び受容的になり、雄がアルファの地位を享受できる時間は短い可能性が高いからです。これが初期の人類において問題であったかどうかは依然として議論の余地がありますが、現代社会と狩猟採集民文化の研究では、乳児は生物学的な父親よりも義父によって虐待されたり、殺されたりするリスクがはるかに高いことが示されています。
雄と雌の親投資の量の違いは、両性の生存率にも反映されています。霊長類では、生涯を通じてほとんどの場合、雌が有利ですが、雄が子を運ぶ種では、生存における有利性の差は消滅するか、あるいは雄に有利に逆転することさえあります。類人猿では、雄は子育てにほとんど貢献せず、雌には大きな生存の利点があります。雌の雄に対する生存の比率は、チンパンジーでは約1.4、ゴリラでは約1.2です。人間では1.05から1.08に減少し、これは他の類人猿と比較して、子供に対する父親のケアの相対的な大きさを示しています。
すでに示したように、これらの生物学的要因の多くは、人間の行動、特に配偶者選択を形作ってきました。広範な研究により、平均して女性はわずかに年上の社会的地位の高い男性を配偶者として好み、一方男性は腰と尻の比率と胸のサイズによって魅力が示される若い女性を好むことが示されています。男性も女性も、左右対称性の低いパートナーを好みます。なぜなら、高い左右非対称性は、より大きな突然変異負荷、寄生虫感染、または他の環境ストレス要因と関連していることが示されているからです。したがって、潜在的な配偶者の繁殖の「質」は、主に無意識的に評価されます。さらに、配偶者選択はパートナーの類似性の程度によっても影響を受け、完全にランダムではないという証拠があります(「同類婚」と呼ばれます)。
性的選択された人間の特性として出現したメカニズムの変動の極端なものとしての精神病理学的相関は豊富です。潜在的なパートナーとしての自分の資質を誇示することは、おそらく躁病エピソードや演技性パーソナリティ障害の重要な側面の一つです。配偶者防衛は、典型的には男性に見られる嫉妬妄想の症例において病的に誇張されます。「雌の選択」の病的な変動は、おそらく女性に最も多く見られる、他人から愛されているという妄想的な確信である色情妄想に反映されています(詳細は第10章と第14章を参照)。
解説補足
- 有性生殖(ゆうせいせいしょく、sexual reproduction): 異なる個体からの遺伝子を持つ配偶子が合体して新しい個体を形成する生殖様式。
- 無性生殖(むせいせいしょく、asexual reproduction): 単一の個体から遺伝的に同一の子孫が生まれる生殖様式。
- 配偶子(はいぐうし、gamete): 生殖細胞。動物の卵や精子、植物の花粉や卵細胞など。
- 遺伝物質(いでんぶっしつ、genetic material): DNAのこと。
- 組み換え(くみかえ、recombination): 減数分裂の際に、相同染色体間で遺伝子の一部が交換される現象。遺伝的多様性を生み出す要因の一つ。
- 有害な突然変異(ゆうがいなとつぜんへんい、deleterious mutations): 生物の生存や繁殖に不利な影響を与える突然変異。
- 病原菌(びょうげんきん、pathogenic germs): 病気を引き起こす細菌などの微生物。
- 寄生虫(きせいちゅう、parasite): 他の生物に寄生して栄養を摂取し、宿主に害を与える生物。
- 代謝資源(たいしゃしげん、metabolic resources): 生物が生命活動を維持するために必要なエネルギーや物質。
- 妊娠(にんしん、pregnancy): 雌が胎児を体内で育てる期間。
- 授乳(じゅにゅう、lactation): 雌が乳腺から乳を分泌し、子に与えること。
- 養育(よういく、nurturing): 子の世話をすること。
- 保護(ほご、protection): 子を危険から守ること。
- 受精(じゅせい、fertilization): 雄の配偶子と雌の配偶子が合体すること。
- 識別力が強い(しきべつりょくがつよい、more discriminating): 相手を注意深く選び、安易に受け入れないこと。
- 同性内競争(どうせいないきょうそう、intrasexual competition): 同じ性別(通常は雄)の個体間で、異性へのアクセスを巡って起こる競争。
- 繁殖成功度(はんしょくせいこうど、reproductive success): 個体が次世代に遺伝子を残す能力。
- 受精可能な卵(じゅせいかのうなたまご、fertile eggs): 受精する能力のある卵。
- アクセス可能な受精可能な雌(アクセスかのうなじゅせいかのうなめす、accessible fertile females): 雄が交尾できる状態の雌。
- 制限された資源(せいげんされたしげん、limiting resource): 他の個体が競争する対象となる、希少な資源。この場合は、繁殖においてより多くの投資をする性(通常は雌)が該当します。
- 霊長類(れいちょうるい、primates): サル、キツネザル、ヒトなどを含む哺乳類のグループ。
- 幼年期(ようねんき、juvenile period): 出生から性的に成熟するまでの期間。
- 単胎出産(たんたいしゅっさん、singleton births): 一度の出産で一人の子を産むこと。
- 長い出産間隔(ながいしゅっさんかんかく、long birth intervals): 次の出産までの期間が長いこと。
- 核家族(かくかぞく、nuclear family): 親と未婚の子供からなる家族。
- 一夫多妻制(いちぶたさいせい、polygyny): 一人の雄が複数の雌と配偶関係を持つこと。
- 父親としての投資(ちちおやとしてのかとうし、paternal investment): 父親が子育てにかける時間、エネルギー、資源など。
- 長期的なパートナーシップ(ちょうき的なパートナーシップ、long-term partnerships): 長い期間にわたる配偶関係。
- 配偶者防衛(はいぐうしゃぼうえい、mate guarding): 自分の配偶者が他の異性と関係を持たないように監視したり、干渉したりする行動。
- 嫉妬(しっと、jealousy): 自分の配偶者が他の異性に興味を持つことに対して抱く不安や怒りの感情。
- 隠蔽排卵(いんぺいはいらん、concealed ovulation): 雌が排卵期であることを示す明らかな身体的兆候がないこと。人間や一部の霊長類に見られます。
- 性的強制(せいてききょうせい、sexual coercion): 雌の意思に反して雄が性的関係を強いる行動。
- 感情の側面(かんじょうのそくめん、emotional dimension): 感情的な経験や反応。
- 父親であることの不確実性(ちちおやであることのふかくじつせい、uncertain paternity): 子供の生物学的な父親が誰であるか確信できないこと。
- 托卵(たくらん、cuckoldry): 配偶者以外の異性と性的関係を持ち、その結果生まれた子供を自分の子供として育てさせること。
- 陰茎亀頭(いんけいきとう、glans penis): 陰茎の先端部分。
- 精子競争(せいしきょうそう、sperm competition): 一匹の雌が複数の雄と交尾した場合に、受精を巡って精子同士が競争する現象。
- 交尾後の遅延(こうびごのちえん、post-copulatory deferral): 交尾後に行われる同性内競争。
- 一夫一妻制(いちぶいっさいせい、monogamy): 一組の雄と雌がペアを形成し、共同で子育てを行う繁殖システム。
- 排卵期(はいらんき、ovulation): 卵巣から卵子が放出される時期。
- 体臭(たいしゅう、body odours): 体から発する匂い。
- 顔の特徴(かおのとくちょう、facial features): 顔の形やパーツの配置。
- 男らしい(おとこらしい、masculine): 男性的な特徴を持つこと。
- 長期的な関係(ちょうきてきなかんけい、long-term relationship): 長い期間にわたる男女の関係。
- ゴリラ(gorillas): 大型類人猿の一種。
- 精巣の重さ(せいそうのおもさ、testes weight): 精巣の質量。精子生産量と関連することがあります。
- 単雄複雌(たんゆうふくし、unimale polygyny): 一匹の雄が複数の雌と配偶関係を持つ社会構造。
- チンパンジー(chimpanzees): 大型類人猿の一種。
- シルバーバックゴリラ(silverback gorilla): 成熟した雄のゴリラ。背中に銀色の毛が生えるのが特徴。
- 多雄多雌(たゆうたし、multimale-multifemale groups): 複数の雄と複数の雌が一緒に生活する社会構造。
- 発情期(はつじょうき、oestrus): 雌が交尾を受け入れる期間。
- 乱交的(らんこうてき、promiscuously): 複数の異性と性的関係を持つこと。
- 膣管(ちつかん、vaginal tract): 雌の生殖器官の一部。
- 卵(たまご、egg): 雌の配偶子。
- 左右対称性(さゆうたいしょうせい、symmetry between bilateral characters): 左右の体の部分が鏡像のように対応していること。
- 変異負荷(へんいふか、mutation load): 個体が持つ有害な突然変異の数。
- 寄生虫感染(きせいちゅうかんせん、parasitic infections): 寄生虫が体内に侵入し、増殖すること。
- 環境ストレス要因(かんきょうストレスよういん、environmental stressors): 生物に悪影響を与える環境の要因。
- 繁殖の質(はんしょくのしつ、reproductive ‘quality’): 遺伝的な健康さや、子孫を残す能力の高さ。
- 無意識的に評価される(むいしきてきにひょうかされる、largely unconsciously evaluated): 意識しないレベルで判断されること。
2.2.3. 親子間の対立
親と子の間には、個々の子への親の投資の量と期間に関して、利害の対立が存在します。親と子の遺伝的利益の間には重複があるのは当然ですが、利益は同一ではありません。子の個体発生の発達のある時点で、親が他の子に投資することは親の利益になります。なぜなら、遺伝子中心の観点から見ると、親は自分の子孫とわずか50パーセントしか共有していないため、繁殖成功度を最大化するためにより多くの子を産むことが理にかなっているからです。しかし、子の利益は、親が与えようとする以上の投資を得ることですが、それは、親がその特定の子を養育することが2倍以上の費用がかかる時点までです。なぜなら、完全な兄弟姉妹も遺伝子の50パーセントを共有しているからです。したがって、この時点で、選択は親(ここでは母親)と子の両方に作用して、投資を終わらせるはずです。これは、すべての有性生殖を行う種に当てはまります。
哺乳類では、親子間の対立は妊娠中にもすでに問題になる可能性があります。例えば、胎児の遺伝子は、母親の利益になるよりも多くの資源を母親の生物から抽出するように選択される可能性があります。例えば、母親のインスリンに対する感受性は、胎盤ホルモンによって下方制御され、それによって母親の血糖値が上昇し、胎児へのエネルギー供給が増加します。EPH(浮腫、蛋白尿、高血圧)妊娠中毒症は、この対立の極端な(そして病的な)変異と見なすことができます。EPH妊娠中毒症は、胎児と母親の最適な血糖値の間の調節不全のために、母親に深刻な健康問題を引き起こす可能性があります。
哺乳類に典型的な別の親子間の対立は、離乳です。通常、乳児は、母親が提供するように選択されているよりも、より大きく、より長い投資(ここでは、ミルクの供給の観点から)を要求するように選択されます。この理論はまた、乳児の将来の兄弟姉妹が、完全な兄弟姉妹よりも遺伝的に遠い関係にある場合、離乳の対立が悪化すると予測しています。言い換えれば、将来の兄弟姉妹が異父兄弟である場合、つまり異なる男性によって父親である場合、離乳の対立は激化します。したがって、人間では、異なる父親であることはおそらく例外であるため、離乳の対立は、他の哺乳類と比較して、実際には弱い可能性があります。乳児と母親の間の同様の対立は、乗ることや運ぶことについても生じる可能性があります。人間の子供は、自分で歩くことができるにもかかわらず、しばしば運ぶことを要求します。
乳児(人間と人間以外)は、親の投資を誘発するために、さまざまな行動を持っています。乳児は親よりもはるかに小さく、経験が浅く、親は利用可能な資源を管理しているため、乳児は親の投資の量を増やすために、物理的な手段ではなく、心理的なメカニズムを発達させるように選択されます。空腹時や危険を感じたときに乳児が泣くことは、そのような例かもしれません。子が特に無力で脆弱な種では、親は子のニーズにより迅速に対応するように選択されています。これは、人間の進化全体を通して確かに重要でした。なぜなら、人間の赤ちゃんは、生理学的に早産であり、したがって、他の霊長類の赤ちゃんと比較して非常に未熟だからです(第3章を参照)。子が成長するにつれての「戦術」は、親の投資を増やすために、初期の個体発生段階の行動に戻ることかもしれません。この現象は、人間心理学でよく知られており、「退行」と呼ばれています。かんしゃくは、親に投資を増やすことを強制する別の方法です。自然な状況下では、かんしゃくの存在は直感的ではありません。なぜなら、それは捕食者を引き付ける可能性があり、エネルギー的に費用がかかるからです。ある意味で、かんしゃくは親の投資を増やすための危険な戦略ですが、しばしば非常に成功します。捕食者に襲われるリスクを冒す個々の子はまた、遺伝的な関係とすでに投資されたすべての資源の潜在的な損失のために、親の適応度を潜在的に低下させます。
兄弟姉妹間の相互作用も、親子間の対立の理論の観点から解釈できます。親はすべての個々の子と等しく関係があるため、理論的には、資源が子間で平等に分配されることが親の利益になります。しかし、遺伝的な関係からすると、個々の子は、兄弟への利益が提供者へのコストの2倍の大きさである場合にのみ、完全な兄弟に対して利他的な行為を行うことになります。言い換えれば、親は兄弟間の利他主義を奨励し、利己的な行動を阻止するように選択されます。
他の形の親子間の対立は、子の成体(生殖)の役割についても生じる可能性があります。トリヴァースは、人間の親は自分の子供の配偶者選択に特に関心を持つべきだと主張しました。なぜなら、親の利益のために、近親者への子の利他主義を高めるために、近親者との結婚を奨励することがあるからです(この傾向は、近親相姦の回避によって打ち消されます)。一方、子の方が親よりも早く親の投資を終了させたい状況もあるかもしれません。これは、例えば、親が個々の子の繁殖を妨げ、その子を「巣のヘルパー」として保持したい場合です。そのような対立は、成人した子供にとって特に解決が難しいかもしれません。
親子間の対立の特別な変種は、子の性別を指します。通常、自然選択は、息子と娘に等しく投資する親を有利にし、約50:50の性比につながります。しかし、良好な条件では、親がより多くの息子を産むことは有利になる可能性があります。なぜなら、息子の繁殖成功度は娘のそれよりも大きい可能性があるからです。しかし、悪い状況では、選択は娘により多くの投資をする親を有利にする可能性があります。なぜなら、環境条件は男性よりも女性に不均衡に小さい影響を与えるからです(トリヴァース-ウィラード仮説)。これらの生物学的に決定された偏りは、人間や他の哺乳類における(認識された?)環境条件の関数として、子宮内での男性の死亡率に深刻な影響を与えることが示されています。一部の哺乳類は、環境条件に応じて、妊娠中に子の性別を決定し、適切な性別でない場合は胎児を中絶することさえできます。人間や他の霊長類のように、親の投資期間が長くなると、息子と娘に対する親の行動の差の偏りが大きくなります。しかし、父親の投資の役割は、これらの偏りをさまざまな方法で修正する可能性があります。
人間における息子と娘への異なる投資の存在は、特に道徳的な問題が生物学的な原則と混同されている場合、大きな議論の的です。しかし、乳児殺しは、霊長類と人間が親の投資の調整を出生後まで延期する方法である可能性があるという証拠があり、これは祖先または現代の狩猟採集民社会でのみ起こったことではないようです。そのような場合、母親が新生児への愛着を発達させる前(第3章と第16章も参照)、ほぼ常に新生児殺し(出生後24時間以内)が行われます。しかし、子供が母親に受け入れられるかどうかは、母親の年齢や生活状況にも依存します。これは、現代社会における母親の中絶の決定にも深刻な影響を与えます。例えば、シングルマザーは、既婚女性よりも妊娠中絶する可能性が高くなります。これは、若い女性の方が結婚し、他の子供を産む将来の見込みがはるかに高く、年齢とともに低下することに関係しています。その結果、生殖期の終わりに近い高齢の女性は、たとえ一人暮らしであっても、妊娠中絶する可能性は低くなります。対照的に、高齢の既婚女性、特にすでに1人または2人の子供がいる女性では、中絶率が増加します。先天性欠損症の恐怖は、意思決定において主導的な役割を果たしているようには見えません。なぜなら、中絶率の急増は、独身の高齢女性では並行して起こらないからです。
親投資理論と親子間の対立に関連する問題の組み合わせは、ゲノムインプリンティングに反映されています。父親由来および母親由来の遺伝子は、母親の生物によって提供される資源を巡って競合する可能性があります。この問題は、「遺伝学」のセクションで扱います。
進化生物学の専門用語で表現されていますが、個体間の遺伝的構成の違いによる対立の可能性は、人間の心理学と精神病理学の理解に深刻な影響を与えます。妄想的な信念やパーソナリティ障害の例はすでに述べました。生物学的に引き起こされる対立が現れるもう一つの重要な分野は、心理療法です。生物学的な対立の領域は、個人にとってほとんど無意識であることは、いくら強調してもしすぎることはありません。それらは原則として意識的な認識にアクセス可能ですが、おそらく制御することは困難です。心理療法的な関係は、おそらく主に親子間の対立の投影によって特徴付けられ、特に境界性パーソナリティ障害などの重度のパーソナリティ障害は、治療において、親の投資を誘発する行動と拒絶のすべての配列を示す可能性があります(詳細については、第14章と第15章を参照)。同様に、うつ病や不安障害などの防御メカニズムによって特徴付けられる精神病理学的状態は、しばしば「退行」に類似した行動を利用し、「脅威なし」を強く示し、ケアを最大化することを目指します(第11章と第12章を参照)。
解説補足
- 親子間の対立(おやこかんのたいりつ、Parent-offspring conflict): 親と子では、最適な繁殖戦略が異なるため、互いの利益が対立する現象。ロバート・トリヴァースによって提唱されました。
- 個体発生(こたいはっせい、ontogenetic development): 受精卵から成体になるまでの個体の発達の過程。
- 遺伝子中心の観点(いでんしちゅうしんのかんてん、gene-centered point of view): 進化を遺伝子の視点から捉える考え方。
- 繁殖成功度(はんしょくせいこうど、reproductive success): 個体が次世代に遺伝子を残す能力。
- 完全な兄弟姉妹(かんぜんなきょうだいしまい、full siblings): 両親を同じくする兄弟姉妹。遺伝子の50%を共有します。
- 有性生殖(ゆうせいせいしょく、sexually reproducing species): 卵と精子などの配偶子を介して子孫を作る生物の繁殖様式。
- 胎児(たいじ、foetus): 妊娠中の哺乳類の子供。
- 胎盤ホルモン(たいばんホルモン、placental hormones): 胎盤から分泌されるホルモン。
- EPH妊娠中毒症(EPHにんしんちゅうどくしょう、EPH gestosis): 妊娠中に起こる高血圧、蛋白尿、浮腫を特徴とする病態。
- 離乳(りにゅう、weaning): 乳児が母親の乳を飲むのをやめる時期。
- 異父兄弟(いふきょうだい、half-siblings): 父親が異なる兄弟姉妹。遺伝子の25%を共有します。
- 退行(たいこう、regression): 心理的なストレスなどによって、より幼い発達段階の行動パターンを示すようになること。
- かんしゃく(tantrums): 欲求不満などから激しく泣いたり、わめいたりする行動。
- 捕食者(ほしょくしゃ、predators): 他の動物を捕食する動物。
- 兄弟姉妹間の相互作用(きょうだいしまいかんのそうごさよう、Interaction between siblings): 兄弟姉妹間の関係や行動。
- 利他的な行為(りたてきなこうい、altruistic act): 自分の利益を損なってでも、他者の利益になるような行動。
- 利己的な行動(りこてきなこうどう、selfish behaviours): 自分の利益のみを追求する行動。
- 近親相姦の回避(きんしんそうかんのかいひ、incest avoidance): 近親者との性的関係を避ける傾向。
- 巣のヘルパー(すのヘルパー、helper at the nest): 親の繁殖を助けるために、自分の繁殖を遅らせる子供。
- 性比(せいひ、sex ratio): 集団における男女の比率。
- トリヴァース-ウィラード仮説(Trivers-Willard hypothesis): 親の体の状態が良いときには息子を、悪いときには娘を多く産む方が繁殖上有利になるという仮説。
- 子宮内での男性の死亡率(しきゅうないでのだんせいのしぼうりつ、intrauterine male mortality): 妊娠中に死亡する男性胎児の割合。
- ゲノムインプリンティング(genomic imprinting): 遺伝子の由来(父親由来か母親由来か)によって、遺伝子の発現が異なる現象。
- 遺伝的構成(いでんてきこうせい、genetic makeup): 個体が持つ遺伝子の組み合わせ。
- 妄想的な信念(もうそうてきなしんねん、delusional beliefs): 事実と異なることを確信してしまうこと。
- パーソナリティ障害(パーソナリティしょうがい、personality disorders): 特徴的な行動パターンが極端に偏り、社会生活に支障をきたす状態。
- 心理療法(しんりりょうほう、psychotherapy): 心理的な問題の解決を支援する治療法。
- 境界性パーソナリティ障害(きょうかいせいパーソナリティしょうがい、borderline personality disorder): 気分の変動が激しく、対人関係が不安定になりやすいパーソナリティ障害。
- 防御メカニズム(ぼうぎょメカニズム、defence mechanisms): 無意識のうちに、不快な感情や状況から自分を守ろうとする心理的な働き。
- うつ病(うつびょう、depression): 気分の落ち込みや意欲の低下が続く精神疾患。
- 不安障害(ふあんしょうがい、anxiety disorders): 過剰な不安や恐れを感じる精神疾患。
はい、承知いたしました。第1章の「3. 進化心理学の原理」の部分を、高校の理科の教科書のような文体で平易に理解しやすく翻訳し、必要に応じて解説を補足します。
3. 進化心理学の原理
進化心理学の前提は、人間の心は、すべての生物の形態的特徴と同じ生物学的法則に従って進化したということです。したがって、自然選択と性選択は、私たちが考え、感じ、行動し、愛着を抱き、生き残る方法を形作ってきました。
進化心理学と、動物行動学、社会生物学、行動生態学などの関連分野は、人間の心の働き方に関するいくつかの誤った前提を反駁することに貢献してきました。そのような前提の一つは、生まれたばかりの人間の赤ちゃんの心は「タブラ・ラサ」(白紙)であるというものでした。私たちは現在、注意深い観察から、これが事実ではないことを知っています。例えば、新生児は、機能的にはもはや有用ではないものの、モロー反射や把握反射など、私たちの霊長類の遺産に深く根ざした原始的な反射をいくつか保持しています。さらに、生まれたばかりの赤ちゃんは、愛着と絆を強めるために、通常は母親である主要な養育者と非常に洗練された方法で容易に相互作用することができます。赤ちゃんは、幾何学的な形よりも顔のような形を好み、一人にされること、高い場所、大きな音などに対する生得的な恐怖を持ち、顔の表情を模倣する能力を持っています。これらは、他の多くのものと同様に、私たちの種特有の環境条件への適応として出現した高度に適応的な心理的メカニズムです。
しかし、ほとんどの心理的メカニズムは、経験による修正に対して非常に開かれています。つまり、それらは(はるかに本能主導である「閉じたプログラム」とは対照的に)人間の生涯を通じて学習経験に対応する「開かれたプログラム」を表しており、一部の期間は「刷り込み」のようなプロセスにとって重要です。
言い換えれば、心理的メカニズムが生理的に機能するためには、生得的な素質(「生得性」)が、刺激的な環境からの適切な入力(「育ち」)と出会うことが不可欠です。そうでなければ、機能不全に苦しむ可能性があります。例えば、幼い子供の軽い斜視が見過ごされた場合、「非優位眼」は決して見ることを学習せず、弱視のままになる可能性があります。弱視では、患部に器質的な異常はありません。むしろ、その障害は、患部が適切に刺激されないという発達上の問題から生じます。同様に、言語、共感など、他の多くの進化した心理的メカニズムも、個体が適切な刺激、つまり環境からの入力を奪われた場合、本質的に同じように起こる可能性があります。
解説補足
- 進化心理学(しんかしんりがく、evolutionary psychology): 人間の心理的特性や行動は、進化の過程で自然選択や性選択によって形成された適応であると考える心理学の分野。
- 形態的特徴(けいたいてきとくちょう、morphological characteristics): 生物の体の形や構造の特徴。
- 動物行動学(どうぶつこうどうがく、ethology): 動物の自然な環境における行動を研究する学問。
- 社会生物学(しゃかいせいぶつがく、sociobiology): 動物の社会行動を進化論的な視点から研究する学問。
- 行動生態学(こうどうせいたいがく、behavioural ecology): 生物の行動を、その生態学的環境への適応として研究する学問。
- タブラ・ラサ(tabula rasa): ラテン語で「白紙」を意味し、生まれたばかりの赤ちゃんの心は何も書き込まれていない白紙の状態であるという哲学的な考え方。
- 原始的な反射(げんしてきなはんしゃ、primitive reflexes): 新生児に見られる、生後数ヶ月で消失する反射的な行動。
- モロー反射(モローはんしゃ、Moro reflex): 大きな音や急な体位の変化に反応して、赤ちゃんが両腕を広げ、抱きつくような動作をする反射。
- 把握反射(はあくはんしゃ、grasp reflexes): 赤ちゃんの手のひらに何かが触れると、強く握る反射。
- 霊長類の遺産(れいちょうるいのいさん、primate heritage): 人類が進化の過程で受け継いできた、霊長類に共通する特徴。
- 主要な養育者(しゅようなよういくしゃ、primary care figure): 赤ちゃんの世話をする最も重要な人物(通常は母親)。
- 愛着と絆(あいちゃくときずな、attachment and bonding): 赤ちゃんと養育者の間に形成される、親密な感情的なつながり。
- 生得的な恐怖(せいとくてきなきょうふ、inborn fear): 生まれつき持っている恐怖。
- 顔の表情を模倣する能力(かおのひょうじょうをもほうするのうりょく、ability to imitate facial expressions): 生まれたばかりの赤ちゃんが、養育者の顔の表情を真似る能力。
- 高度に適応的な心理的メカニズム(こうどにてきおうてきなしんりてきメカニズム、highly adapted psychological mechanisms): 特定の環境に適応するために進化した、心の働き。
- 種特有の環境条件(しゅとくゆうのかんきょうじょうけん、species-specific environmental conditions): その種が進化してきた特有の環境。
- 経験による修正(けいけんによるしゅうせい、experiential modification): 経験や学習によって変化すること。
- 開かれたプログラム(ひらかれたプログラム、open programmes): 生得的な基盤を持ちながらも、経験によって柔軟に変化する心理的メカニズム。
- 閉じたプログラム(とじたプログラム、closed programmes): 生得的に固定されており、学習による影響を受けにくい、本能的な行動パターン。
- 刷り込み(すりこみ、imprinting): 特定の感受性期に起こる、強固で不可逆的な学習。
- 生得的な素質(せいとくてきなそしつ、innate predispositions): 生まれつき持っている傾向や能力(「生得性」)。
- 刺激的な環境(しげきてきなかんきょう、stimulating environment): 発達に必要な適切な刺激を提供する環境(「育ち」)。
- 機能不全(きのうふぜん、dysfunction): 本来の機能を果たせなくなること。
- 斜視(しゃし、strabismus): 両眼の視線が一致しない状態。
- 非優位眼(ひゆういがん、non-dominant eye): 左右の目のうち、あまり使わない方の目。
- 弱視(じゃくし、amblyopia): 器質的な異常がないにもかかわらず、視力が発達しない状態。
- 器質的な異常(きしつてきないじょう、organic failure): 臓器や組織に構造的な異常があること。
- 発達上の問題(はったつじょうのもんだい、developmental problem): 発達の過程で生じる問題。
- 適切な刺激(てきせつなしげき、appropriate stimulation): 発達に必要な適切な環境からの入力。
- 共感(きょうかん、empathy): 他者の感情を理解し、共有する能力。
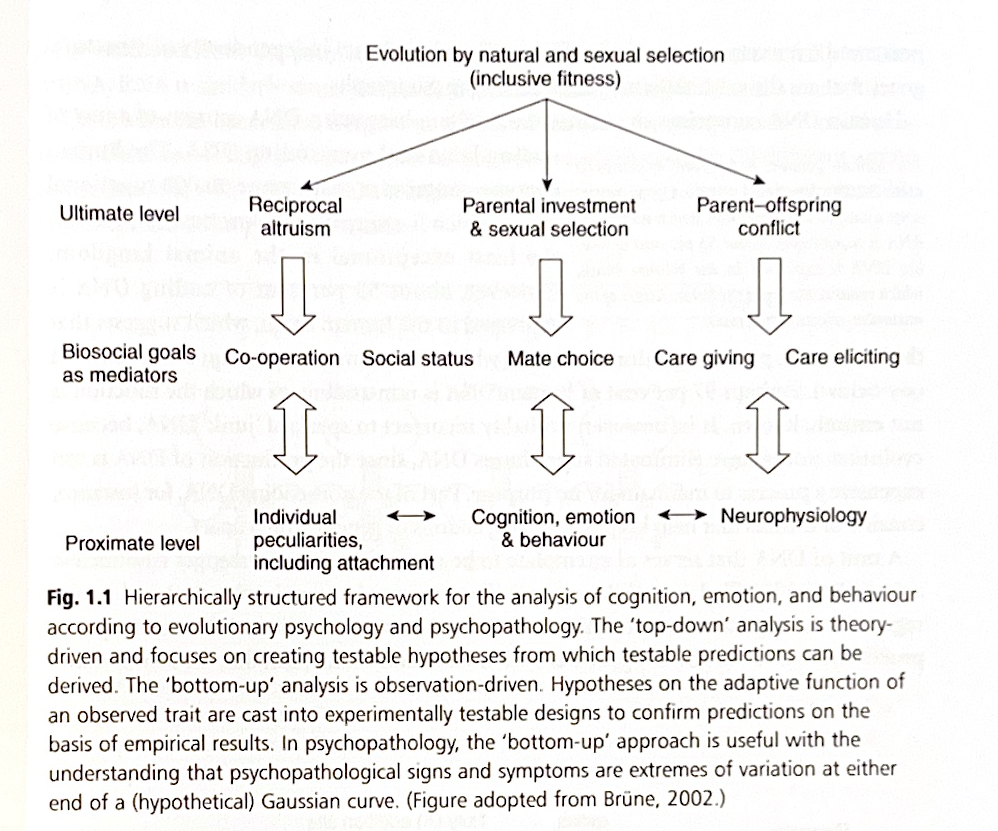
3.1. 進化した心理的メカニズムと生物社会的目標
人間の生物-社会-心理的な機能は、私たちの進化の過去における選択圧に対応して進化した心理的メカニズムに満ちています(「心理的メカニズム」という用語は、硬直性や学習による修正に対する不浸透性を示唆するため、洗練されていないことを強調する必要があります。しかし、これは進化心理学者が実際に意図している意味ではありません)。ある意味で、心理的メカニズムは、適応上の重要な特定の問題を解決するために進化によって設計されています。そうするために、それらは経済的で、信頼性が高く、効率的で、正確である必要があります。これらの特性は、複数の情報源からの情報を統合するいくつかの領域一般メカニズムが存在する可能性とともに、領域特異的な機能によって最もよく維持されます。領域特異性は、適応上の問題が進化の過程でかなりの期間にわたって繰り返し存在し、個体の繁殖成功に影響を与えてきた場合に想定されます。多くの心理的メカニズムは、他のモジュールとはほぼ独立して、特定の種類の情報のみを処理する精神的なモジュールとして概念化できます。モジュールは、注意、感情的な評価、および迅速な反応のための行動反応を集中させるのに役立つ決定ルールまたはトリガーアルゴリズムによって活性化されます。モジュールは、効率を高めるために階層的に組織されていると考えられています。例えば、暗闇の中で一人でいるなどの警戒心が高まっている状況下で活性化される脅威検出のための基本的なモジュールは、動く形状の検出に焦点を当てるアルゴリズムによって活性化される可能性があります。その動く形状が(空気の循環によって生じるランダムな動きとは対照的に)自走していると識別された場合、動きの方向を調べるアルゴリズムによって、より高次のモジュールが活性化される可能性があります。動きの方向が個人に向いている場合、その動く物体が動物か人間かを判断するアルゴリズムによって、別のモジュールがトリガーされる可能性があります。以下同様です。
進化した心理的メカニズムは、単一の万能ツールではなく、さまざまなタスクを非常に効率的な方法で実行するように設計された個別のデバイスのコレクションを含むスイスアーミーナイフの多様な機能に例えられてきました。ほとんどの比喩的な類似性と同様に、人間の脳とスイスアーミーナイフの比較は理想的ではありません(例えば、スイスアーミーナイフを意図的に扱う人がいなければなりませんが、複数のタスクの協調的な実行は人間の脳自体の別の機能です。したがって、進化論的な観点からは、人間の心の二元論的な概念化の余地はありません)。しかし、この類似性は、意思決定と推論のプロセスが抽象的でも論理的でもなく、人間の進化における繰り返しの問題に対する選択された答えであることを示しています。進化心理学の最も重要な発見の1つは、おそらくほとんどの人間の脳機能が、抽象的な問題よりも社会的な問題の解決に偏っており、進化した心理的メカニズムは、特に社会的な推論に関しては、ばかばかしいほど非論理的であることが多いということです(社会脳仮説に関する詳細については、第2章を参照)。
人間の種にとって社会生活が非常に重要であり、人間の心に作用してきた多岐にわたる選択圧を考慮すると、生物社会的目标の追求は、進化した心理的メカニズムの実際の標的です。他の生物と同様に、人間は、自身の個々の適応度を最大化するように意識的に行動が導かれるという意味で、「適応度最大化者」ではないことを強調する必要があります。包括適応度は事後的にしか測定できません。将来の繁殖成功度は計算不可能です。したがって、進化した「戦略」とも呼ばれる進化した心理的メカニズムは、近接レベルで作用します。なぜなら、生物社会的目标の達成における成功は、繁殖の成功の可能性を高めるからです(それは適応度の指標となる可能性もありますが、そうである必要はありません)。注目すべきは、「戦略」という言葉は、特定の生物社会的目标を達成する方法の意識的な認識を伴わないということです。
人間の生物社会的目标には、他者からのケアを引き出す動機と行動、主に親族や親しい同盟者へのケアの提供、適切な配偶者の発見、協力的な同盟の形成、そして可能な限り最高の社会的地位の獲得が含まれる可能性があります。生物社会的目标はまた、劣等感や優越感の観点から自分の社会的地位を他者の地位と比較し、より高い地位やより低い地位の個人との関係を交渉することを目指します。厳密に言えば、生物社会的目标は近接レベルの一部です。なぜなら、それらは個人の生涯にわたって変化する可能性があるからです。例えば、個人は社会的階層のはしごを上り下りしたり、他者への社会的支援の努力を変えたりすることができます。したがって、生物社会的目标は、生理学的プロセス、遺伝子の作用、または個人の養育条件によって媒介される認知、感情、行動の基本的な近接原因と、個体間の遺伝的類似性および非類似性によって引き起こされる問題との間のある程度中間的な位置を占めています。男性と女性は目標の優先順位が著しく異なり、これは祖先時代の男性と女性が担っていた異なる役割(下記参照)と、異なる繁殖上の制約に関連しています。したがって、男性にとっては地位と資源へのアクセスがより重要であった可能性があり、一方女性は強い親和的な絆と感情的なつながりに、より重点を置いていた可能性があります。
注目すべきことに、進化した心理的メカニズムは、最適に設計されていることはめったになく、もしそうであっても、設計の最適性は進化するにはあまりにも高価であるため、しばしば設計上の妥協を表しており、本質的に誤るリスクを抱えています。同様に、相互依存と社会関係を円滑にする必要性は、心理的な障害につながる可能性のある問題を生み出します。これらの問題と精神病理の他の可能な原因は、第4章およびその後の臨床の章でさらに探求されます。
解説補足
- 生物社会-心理的な機能(せいぶつしゃかい-しんりてきなきのう、bio-social-psychological functioning): 人間の生物学的、社会的、心理的な側面が相互に関連し合って働くこと。
- 心理的メカニズム(しんりてきメカニズム、psychological mechanisms): 特定の適応上の問題を解決するために進化した、心の働き。
- 選択圧(せんたくあつ、selection pressures): 生物の生存や繁殖に影響を与える環境要因。
- 硬直性(こうちょくせい、rigidity): 柔軟性がないこと。
- 不浸透性(ふしんとうせい、imperviousness): 影響を受けにくいこと。
- 適応上の重要性(てきおうじょうのじゅうようせい、adaptive significance): 生物の生存や繁殖にとっての重要性。
- 経済的(けいざいてき、economical): 無駄がなく、効率的であること。
- 信頼性(しんらいせい、reliable): 常に安定して機能すること。
- 効率的(こうりつてき、efficient): エネルギーや資源の消費を抑えながら、目的を達成すること。
- 正確(せいかく、precise): 誤りが少ないこと。
- 領域特異的な機能(りょういきていなきのう、domain-specific functioning): 特定の種類の情報や問題に対して特化した心の働き。
- 領域一般メカニズム(りょういきいっぱんメカニズム、domain-general mechanisms): さまざまな種類の情報や問題に対応できる、汎用的な心の働き。
- 適応上の問題(てきおうじょうのもんだい、adaptive problem): 生存や繁殖を脅かす可能性のある課題。
- 精神的なモジュール(せいしんてきなモジュール、mental modules): 特定の情報を処理する、独立した心の機能単位。
- 決定ルール(けっていルール、decision rules): 行動を選択するための規則。
- トリガーアルゴリズム(トリガーアルゴリズム、triggering algorithms): 特定の刺激や状況に応じて、特定の反応を引き起こす仕組み。
- 階層的に組織化される(かいそうてきにそしきかされる、hierarchically organized): 機能が段階的に組み合わさって、より複雑な処理を行う構造。
- 脅威検出(きょういけんしゅつ、threat detection): 危険なものを認識する能力。
- 警戒心が高まっている状態(けいかいしんがたかまっているじょうたい、conditions of heightened vigilance): 周囲の状況に注意深く意識を向けている状態。
- 自走(じそう、self-propelled): 外部からの力によらず、自ら動くこと。
- 高次のモジュール(こうじのモジュール、higher-order module): より複雑な情報を処理する、上位の機能単位。
- スイスアーミーナイフ(Swiss army knife): 多機能な折りたたみナイフのたとえ。人間の心が、さまざまな特定の機能を持つ道具の集まりに似ていることを示す。
- 比喩的な類似性(ひゆてきなるいじせい、metaphorical analogies): あるものを理解するために、似た別のものを引き合いに出して説明すること。
- 二元論的な概念化(にげんろんてきながいねんか、dualistic conceptualizations): 心と体を別々の実体として捉える考え方。進化心理学では否定されます。
- 意思決定と推論のプロセス(いしけっていとすいろんのプロセス、decision-making and reasoning processes): 行動を選択したり、結論を導き出したりする心の働き。
- 抽象的(ちゅうしょうてき、abstract): 具体的な事物や現象から離れた、一般的な概念に関する。
- 論理的(ろんりてき、logical): 筋道が通っていること。
- 社会的な問題(しゃかいてきなもんだい、social problems): 他者との関係や集団生活における課題。
- 社会脳仮説(しゃかいのうかせつ、social brain hypothesis): 霊長類において、集団の規模と脳の大きさ(特に大脳新皮質)が相関しているという仮説。複雑な社会関係に対処するために脳が発達したと考えられています。
- 生物社会的目标(せいぶつしゃかいてきもくひょう、biosocial goals): 生存と繁殖の成功を高めるための、社会的な相互作用に関わる目標。
- 適応度最大化者(てきおうどさいだいかしゃ、fitness maximizers): 自身の適応度を意識的に最大化するように行動する主体。進化心理学では、人間の行動は直接的な適応度最大化を目指すのではなく、生物社会的目标の達成を目指すとされます。
- 包括適応度(ほうかつてきおうど、inclusive fitness): 個体の遺伝子が次世代にどれだけ広まるかを評価する際に、その個体自身の繁殖成功度だけでなく、遺伝的に関連のある個体の繁殖成功度も考慮に入れる概念。
- 事後的にしか測定できない(じごてきにしかそくていできない、can only be measured in retrospect): 結果が出てからでないと評価できないこと。
- 将来の繁殖成功度(しょうらいのはんしょくせいこうど、future reproductive success): 将来、子孫を残せる可能性。
- 進化した「戦略」(しんかした「せんりゃく」、evolved ‘strategies’): 生物社会的目标を達成するために進化した心理的メカニズム。意識的な計画とは異なります。
- 近接レベル(きんせつレベル、proximate level): 行動の直接的な原因やメカニズムのレベル。
- 繁殖の成功(はんしょくのせいこう、successful reproduction): 子孫を残すこと。
- 指標(しひょう、indicator): 何らかの状態や性質を示すもの。繁殖の成功は適応度の一つの指標となりえます。
- 意識的な認識(いしきてきなにんしき、conscious awareness): 自分が何をしているか、なぜそうしているかを自覚していること。
- 他者からのケアを引き出す(たしゃからのケアをひきだす、elicit care from others): 他者に世話をしてもらうように働きかけること。
- 他者へのケアを提供する(たしゃへのケアをていきょうする、provide care for others): 他者を世話すること。
- 親族(しんぞく、kin): 血縁関係のある人。
- 親しい同盟者(したしいどうめいしゃ、close allies): 協力関係にある親しい人。
- 適切な配偶者を見つける(てきせつなはいぐうしゃをみつける、find a suitable mate): 繁殖に適したパートナーを見つけること。
- 協力的な同盟を形成する(きょうりょくてきなどうめいをけいせいする、form cooperative alliances): 他者と協力関係を築くこと。
- 可能な限り最高の社会的地位を獲得する(かのうなかぎりさいこうのしゃかいてきちいをかくとくする、attain the highest social rank possible): 集団の中で高い地位を得ること。
- 劣等感(れっとうかん、inferiority): 自分は他人より劣っていると感じる気持ち。
- 優越感(ゆうえつかん、superiority): 自分は他人より優れていると感じる気持ち。
- 関係を交渉する(かんけいをこうしょうする、negotiate relationships): 他者との関係を調整すること。
- 可塑的(かそてき、malleable): 変化しやすいこと。
- 社会的階層(しゃかいてきかいそう、social hierarchy): 集団内における地位の順序。
- 社会的支援(しゃかいてきしえん、social support): 他者からの助けや支え。
- 基本的な近接原因(きほんてきなきんせつげんいん、basic proximate causes): 行動の直接的な生物学的・心理的な原因。
- 生理学的プロセス(せいりがくてきプロセス、physiological processes): 体内で起こるさまざまな機能。
- 遺伝子の作用(いでんしのさよう、the action of genes): 遺伝子が生物の特性に与える影響。
- 個人の養育条件(こじんのよういくじょうけん、individual rearing conditions): 生育環境や親からの育てられ方。
- 遺伝的類似性および非類似性(いでんてきるいじせいおよびひるいじせい、genetic similarity and dissimilarity): 個体間の遺伝子の類似度や違い。
- 目標の優先順位(もくひょうのゆうせんじゅんい、goal priorities): 達成したい目標の重要度の順序。
- 祖先時代の条件(そせんじだいのじょうけん、ancestral condition): 人類が進化してきた過去の環境や生活様式。
- 繁殖上の制約(はんしょくじょうのせいやく、reproductive constraints): 繁殖の可能性を制限する要因。
- 地位(ちい、status): 集団内における個人の社会的評価や位置。
- 資源へのアクセス(しげんへのアクセス、access to resources): 生存に必要な食料、水、住居などを利用できること。
- 強い親和的な絆(つよいしんわてきなきずな、strong affiliative bonds): 他者との親密で友好的なつながり。
- 感情的なつながり(かんじょうてきなつながり、emotional ties): 他者との感情的な結びつき。
- 設計上の妥協(せっけいじょうのだきょう、design compromises): 進化の過程で、複数の異なる要求を満たすために、完全な最適化を避けて調整された形質やメカニズム。
- 誤るリスク(あやまるリスク、risk to go wrong): 機能がうまくいかない可能性。
- 設計の最適性(せっけいのさいてきせい、design optimality): 特定の目的を最も効率的に達成する設計。
- 相互依存(そうごいぞん、mutual dependence): 個体同士が互いに頼り合って生きていること。
- 社会関係を円滑にする必要性(しゃかいかんけいをえんかつにするひつようせい、need for accommodating social relationships): 他者との良好な関係を築き、維持する必要性。
- 心理的な障害(しんりてきなしょうがい、psychological disturbances): 精神的な健康上の問題。
- 精神病理(せいしんびょうり、psychopathology): 精神疾患や精神的な異常。
- 臨床の章(りんしょうのしょう、clinical chapters): 精神疾患や心理的な問題に関する具体的な記述や議論を行う章。
3.2. 進化的適応環境
私たちを人間たらしめるほとんどの心理的メカニズムは、遠い過去の環境条件下で進化したという考えは、ジョン・ボウルビーによって提唱されました。ボウルビーは、人間の精神性の大部分は、私たちの祖先が更新世(約260万年前から1万年前)と呼ばれる地質時代に狩猟採集民として生活していたときに起源を持つと主張しました。狩猟採集民の生活に関する最近の研究は、祖先の人類が、血縁に基づいた小さく、主に父系居住の部族集団で生活しており、社会構造は比較的平等主義的で、物質的な富の蓄積はほとんどなかったことを示唆しています。このモデルは、少なくとも約15万年前に解剖学的現代人が出現して以来、私たちの種の歴史の大部分で一般的だったと考えられます。狩猟や採集から食料を確保するためには、グループメンバー間の協力が不可欠でした。これには、女性間の協力的な子育て(それによっておそらく出産間隔が5〜6年から3〜4年に短縮された)、および男性間の協力による大型捕食者や競合する人間グループからのグループの保護が含まれていました。さらに、これらの緊密なコミュニティは、おそらく30〜40人以下の個人で構成されており、近隣グループや拡大家族と合わせて、互いに個人的に知り合う最大150人程度の集団を形成していました。交易だけでなく、隣接するグループとの戦争も、おそらく人類の歴史を通じて一般的でした。
希少な資源を巡って激しい競争にさらされている現代の園芸社会からのデータは、男性の約4人に1人が暴力的な死を遂げたことを示唆しており、その主な原因はグループ間の戦闘でした。その結果として、そして今日まで続く人間の悲劇の主な原因として、人間はグループ内での協力への強い傾向があるにもかかわらず、本質的に外国人嫌悪、つまり見知らぬ人を強く不信にする傾向があります。幼児でさえ、生後1年以内に「本能的に」外国人嫌悪を発達させ、特に見知らぬ男性に向けられます。
しかし、単一の進化的適応環境(EEA)のシナリオは、狭すぎると批判されてきました。第一に、更新世以前の適応を無視してはなりません。例えば、人間とチンパンジーの系統は約500万〜600万年前に分岐し、それ以来、直立二足歩行、脳の拡大と再編成、言語など、寒冷化する気候への多くの適応が人間に現れました。これらが起こる以前でさえ、霊長類は主に夜行性の単独生活を送る昆虫食動物から、昼行性の群れをなす採食動物へと進化しました。そして、さらなる社会的適応のために、両眼視、色覚、手と目の協調の進化の重要性を想像することができます。例えば、眼球が頭蓋骨の側面から正面に移動したことは、狩猟スキルと腕渡り(ぶら下がり移動)を向上させた可能性がありますが、以前のほぼパノラマのような視野を狭め、後方から近づく捕食者を検出する能力を低下させるという代償を伴いました。これは、今度は社会集団の形成、ひいては社会環境からの適応上の問題に対処するための脳システムを選択的に有利にした可能性があります(より遠い適応は、私たちの階層的に組織化された「三位一体脳」に反映されています。詳細は第2章を参照)。
第二に、初期の人類がアフリカを離れて事実上地球全体(南極大陸を除く)を歩き回った後に、いくつかの重要な適応が出現しました。すべての人間の密接な遺伝的関連性(「進化的ボトルネック」仮説と呼ばれる)が示すように、おそらく少数の人間グループが出発しただけでしたが、地域集団が適応した多くのEEAがありました(例えば、ランダムに選ばれた2人の人間は、同じ社会グループの2匹のチンパンジーよりも遺伝的にはるかに密接に関連しています)。したがって、EEAは、進化する人間に選択圧を生み出すのに十分な長さ続いたさまざまな側面の統計的な複合体として概念化するのがおそらく最善です。最後に、約15万年前に進化した最初の解剖学的現代人と現代人が異ならないという暗黙の概念は、おそらく完全には正しくありません。例えば、研究者たちは、脳の拡大と人間の言語の進化に関連する可能性のある人間のASPM遺伝子とFoxP2遺伝子ファミリーにおける最近の対立遺伝子変異を発見しました。ASPM遺伝子のそのような変異の1つは、わずか5800年前のものであることが発見されており、最も初期の文書記録との偶然の一致は偶然ではないと推測されています。一方、過去5万年間で人間の脳サイズと体サイズは減少しており、これは攻撃性の低下に向けた選択の結果としての「家畜化」効果に関連している可能性があるという推測を引き起こしました。さらに最近では、その最初の発生が中世に遡ることができ、人々がますます過密な都市に集まるようになったテイ・サックス病の出現は、結核に対するより大きな抵抗力に関連する遺伝子の選択に関連している可能性があります。これらの後者の例は、人間における最も最近の適応でさえ神経系を免れなかったこと(第2章も参照)、そして人間が進化し続ける可能性があることを示しているかもしれません。しかし、現代の環境において、人間の認知、感情、行動に選択がどのように作用するかは、十分に理解されていません(しかし、科学的に根拠のない多くの推測を引き起こしています)。
解説補足
- 進化的適応環境(しんかてきおうかんきょう、Environment of evolutionary adaptedness – EEA): ジョン・ボウルビーが提唱した概念で、人間の主要な心理的メカニズムが適応したと考えられる、遠い祖先の環境。更新世の狩猟採集生活がその典型とされます。1
- 更新世(こうしんせい、Pleistocene): 約260万年前から1万年前までの地質時代。氷河期と間氷期が繰り返された時代で、人類の進化にとって重要な時期とされます。
- 狩猟採集民(しゅりょうさいしゅうみん、hunter-gatherers): 野生の動植物を狩猟や採集によって食料とする人々。
- 父系居住(ふけいきょじゅう、patrilocal): 結婚後、夫婦が夫の親族の近くまたは同じ場所に住む居住形態。
- 平等主義的(びょうどうしゅぎてき、egalitarian): 社会的な階層や地位の差が小さいこと。
- 物質的な富の蓄積(ぶっしつてきなとみのちくせき、accumulation of material goods): 多くの財産を蓄えること。
- 解剖学的現代人(かいぼうがくてきげんだいじん、anatomically modern humans): 現代人と同様の身体的特徴を持つ人類。約30万年前以降にアフリカで出現したと考えられています。
- 協力(きょうりょく、cooperation): 複数の個体が互いに利益をもたらすように行動すること。
- 共同育児(きょうどういくじ、cooperatively raise children): 複数の人が協力して子供を育てること。
- 出産間隔(しゅっさんかんかく、between-birth intervals): 一人の女性が出産してから次の出産までの期間。
- 緊密なコミュニティ(きんみつなコミュニティ、close-knit communities): 強い絆で結ばれた集団。
- 隣接するグループ(りんせつするグループ、neighbouring groups): 近隣に住む他の集団。
- 拡大家族(かくだいかぞく、extended kin): 核家族だけでなく、祖父母、叔父叔母、従兄弟などを含む親族。
- 交易(こうえき、trade): 物資を交換すること。
- 戦争(せんそう、warfare): 集団間の武力衝突。
- 園芸社会(えんげいしゃかい、horticultural societies): 鍬などを用いた簡単な農耕を行う社会。
- 希少な資源(きしょうなしげん、scarce resources): 十分な量が存在しない資源。
- 暴力的な死(ぼうりょくてきなし、violent death): 戦闘、殺人、事故などによる死。
- グループ間の戦闘(グループかんのせんとう、between-group battle): 異なる集団間の戦い。
- 人間の悲劇(にんげんのひげき、human tragedy): 人間の苦しみや不幸。
- グループ内での協力(グループないでのきょうりょく、within-group cooperation): 同じ集団のメンバー間での協力。
- 外国人嫌悪(がいこくじんけんお、xenophobic): 見知らぬ人や異質なものに対して抱く嫌悪感や不信感。
- 本能的に(ほんのうてきに、’instinctively’): 生まれつき持っている性質として。
- 単一のシナリオ(たんいつのシナリオ、a single scenario): 一つの特定の状況設定。
- 更新世以前(こうしんせいぜん、prior to the Pleistocene period): 更新世よりも前の時代。
- 直立二足歩行(ちょくりつにそくほこう、bipedalism): 二本の足でまっすぐに立って歩くこと。
- 脳の拡大と再編成(のうのかくだいとさいへんせい、brain enlargement and reorganization): 脳のサイズが大きくなり、構造や機能が変化すること。
- 言語(げんご、language): 思考や感情を伝達するための、音声や文字などを用いた記号システム。
- 夜行性(やこうせい、nocturnal): 夜に活動すること。
- 単独生活(たんどくせいかつ、solitary living): 一匹で生活すること。
- 昆虫食動物(こんちゅうしょくどうぶつ、insectivores): 昆虫を主食とする動物。
- 昼行性(ちゅうこうせい、diurnal): 昼に活動すること。
- 群れをなす採食動物(むれをなすさいしょくどうぶつ、gregarious foragers): 群れで生活し、食物を探し求める動物。
- 両眼視(りょうがんし、binocular view): 左右の目で見た像を統合して、立体的に捉える視覚。
- 色覚(しきかく、colour vision): 色を識別する能力。
- 手と目の協調(てとめのきょうちょう、hand-eye coordination): 見ているものに合わせて手を正確に動かす能力。
- 社会的適応(しゃかいてきおう、social adaptations): 集団生活に適応するための進化的な変化。
- 眼球の移動(がんきゅうのいどう、movement of the eyeballs): 眼球の位置が側面から正面に移動すること。
- 腕渡り(うでわたり、brachiation): 腕を使って枝から枝へとぶら下がりながら移動する方法。
- パノラマのような視野(パノラマのようなしや、panoramic visual field): 広い範囲を見渡せる視野。
- 後方から近づく捕食者(こうほうからちかづくほしょくしゃ、predators approaching from the rear): 後ろから忍び寄る捕食動物。
- 社会集団の形成(しゃかいしゅうだんのけいせい、the formation of social groups): 複数の個体が集まって生活する集団を作ること。
- 社会環境からの適応上の問題(しゃかいかんきょうからのてきおうじょうのもんだい、adaptive problems from the social environment): 集団生活におけるさまざまな課題。
- 三位一体脳(さんみいったいのう、’triune’ brain): ポール・マクリーンが提唱した、爬虫類脳、旧哺乳類脳、新哺乳類脳の3つの層からなる脳のモデル。進化の異なる段階に対応するとされる。
- アフリカを出て(アフリカをでて、had left Africa): 人類がアフリカ大陸から他の地域へ拡散したこと。
- 地球全体を歩き回る(ちきゅうぜんたいをあるきまわる、roam virtually the entire planet): ほぼ全世界に分布すること。
- 南極大陸(なんきょくたいりく、Antarctica): 最も南に位置する大陸。
- 少数の人間グループ(しょうすうのにんげんグループ、a small group of humans): 人類の初期の移住者の数が少なかったという説。
- 密接な遺伝的関連性(みっせつないでんてきかんれんせい、close genetic relatedness): 遺伝子的な類似性が高いこと。
- 進化的ボトルネック(しんかてきボトルネック、’evolutionary bottleneck’ hypothesis): 過去のある時期に、集団のサイズが一時的に非常に小さくなり、その後の集団の遺伝的多様性が失われたという仮説。
- 地域集団(ちいきしゅうだん、local populations): 特定の地域に住む集団。
- 統計的な複合体(とうけいてきなふくごうたい、statistical composite): さまざまな要素を統計的にまとめたもの。2
- 選択圧を生み出す(せんたくあつをうみだす、create selection pressures): 生物の生存や繁殖に影響を与え、進化の方向性を決める力となること。
- 暗黙の概念(あんもくのがいねん、implicit notion): 明示的には述べられていないが、前提として含まれている考え方。
- 現代人(げんだいじん、contemporary humans): 現在生きている人々。
- 最初の解剖学的現代人(さいしょのかいぼうがくてきげんだいじん、the first anatomically modern humans): 約30万年前に出現した、現代人と同様の身体的特徴を持つ最初の人類。
- 最近の対立遺伝子変異(さいきんのたいりついでんしへんい、recent allelic variation): 遺伝子における、比較的最近生じた異なる遺伝子型の存在。
- ASPM遺伝子(ASPM gene): 脳の発達に関わる遺伝子。3
- FoxP2遺伝子ファミリー(FoxP2 gene family): 言語能力の発達に関わる遺伝子群。4
- 人間の音声の進化(にんげんのおんせいのしんか、the evolution of human speech): 人間が言葉を話す能力が進化してきた過程。
- 文書記録(ぶんしょきろく、written documents): 文字で書かれた記録。
- 偶然の一致(ぐうぜんのいっち、coincidence): たまたま同時に起こること。
- 脳サイズと体サイズの減少(のうサイズとたいサイズのげんしょう、brain size and body size have declined): 人間の脳と体の大きさが、過去数万年で小さくなってきたこと。
- 家畜化効果(かちくかこうか、’domestication’ effects): 動物が家畜化される過程で、攻撃性の低下など特定の形質が選択されること。人間にも同様の選択が働いた可能性を示唆する説。
- 攻撃性の低下(こうげきせいのていか、reduced aggressiveness): 争いや攻撃的な行動が少なくなること。
- テイ・サックス病(Tay-Sachs disease): 遺伝性の神経疾患。5
- 中世(ちゅうせい、medieval times): 西暦5世紀から15世紀頃までのヨーロッパの時代。
- 過密な都市(かみつなとし、overcrowded cities): 人口が密集した都市。
- 結核に対するより大きな抵抗力(けっかくにたいするよりおおきなていこうりょく、greater resistance against tuberculosis): 結核菌に対する免疫力が高いこと。
- 神経系(しんけいけい、nervous system): 脳、脊髄、末梢神経などからなる、情報伝達のネットワーク。
- 現代の環境(げんだいのかんきょう、our modern environments): 現代社会の生活環境。
- 科学的に根拠のない推測(かがくてきにこんきょのないすいそく、scientifically untenable speculation): 科学的な証拠に基づかない憶測。
3.3. 人間の認知、感情、行動の進化的仮説の検証
人間の心理的機能の進化的枠組みの大きな利点の1つは、これらの理論から予測を立て、その予測を検証できる一連の理論が存在することです。進化心理学の仮説を検証するには、トップダウン方式、つまり、親投資理論などの生物の遺伝的関連性に関する主要な理論の1つから導き出された検証可能な仮説に到達する方法を取ることができます。例えば、女性の潜在的な子孫へのより大きな親投資は、男性よりも社会的に高く評価されている配偶者とのパートナーシップを求める可能性が高いという仮説を立てることができます。対照的に、ボトムアップ方式では、実際の行動の観察から始めます。例えば、パートナーの(認識された)社会的地位が低下した場合、女性はより頻繁に離婚する、または女性パートナーの繁殖力が低下した場合(例えば、年齢の関数として)、男性は女性よりも若いパートナーのために女性パートナーをより頻繁に捨てる、といった観察です。このような観察は、その観察の正しい解釈を確認するための検証可能な仮説にすることができます。
認知、感情、行動における性差を調べるための実験的テスト、質問票(ただし、社会的期待に沿った回答という問題が伴います)、進化論とは無関係に収集されたアーカイブデータ、人間の普遍性を探すための異文化比較、そして進化したメカニズムの文化特異的な修正を評価するための異文化比較、特に私たちの最も近縁な親戚である大型類人猿との種間比較など、いくつかの証拠源を使用できます。最後に、精神病理学的兆候や症状でさえ、正常な心理を理解するための有効な情報源として使用できます。コンラート・ローレンツは、行動の病理は、関与する特定のメカニズムについてより多くを語ることができるため、生理学的機能の分析よりも正常な機能に対して優れた説明力を持つ可能性があると主張しました。精神病理が人間の心理を理解するための証拠源として考慮される場合、このようなアプローチは特に有益かもしれません。しかし、今日まで、この見通しは進化心理学において大きく無視されてきました。
解説補足
- 進化的枠組み(しんかてきわくぐみ、evolutionary framework): 生物の特性を進化の視点から理解するための理論的な枠組み。
- 予測(よそく、predictions): 理論に基づいて将来の状態や現象を予言すること。
- 検証可能(けんしょうかのう、testable): 科学的な方法で真偽を確かめることができること。
- トップダウン方式(トップダウンほうしき、top-down way): 一般的な理論から具体的な仮説を導き出し、検証する方法。
- 遺伝的関連性(いでんてきかんれんせい、genetic relatedness): 遺伝的なつながりの強さ。
- 親投資理論(おやとうしりろん、parental investment theory): 親が子孫の生存と繁殖のために費やす資源の量が、配偶者選択や親子間の対立に影響を与えるという理論。
- パートナーシップ(パートナーシップ、partnerships): 配偶者としての関係。
- 社会的地位(しゃかいてきちい、socially highly regarded mates): 社会的に高く評価されている配偶者。
- ボトムアップ方式(ボトムアップほうしき、bottom-up approach): 実際の観察から出発し、その観察を説明する仮説を構築する方法。
- (認識された)社会的地位((にんしきされた)しゃかいてきちい、(perceived) social rank): 他者からどのように評価されているかという認識。
- 繁殖力(はんしょくりょく、fecundity): 子を産む能力。
- 年齢の関数として(ねんれいのかんすうとして、as a function of age): 年齢によって変化すること。
- 証拠源(しょうこげん、sources of evidence): 仮説を検証するためのデータや情報。
- 実験的テスト(じっけんてきテスト、experimental tests): 特定の変数を操作し、その影響を観察する検証方法。
- 性差(せいさ、sex differences): 男性と女性の間の違い。
- 認知(にんち、cognition): 思考、記憶、学習などの精神活動。
- 感情(かんじょう、emotion): 喜び、悲しみ、怒りなどの心の動き。
- 行動(こうどう、behaviour): 生物の外部から観察できる活動。
- 質問票(しつもんひょう、questionnaires): 特定の質問に回答してもらうことでデータを収集する方法。
- 社会的期待(しゃかいてききたい、social expectations): 社会的に望ましいとされる行動や考え方。質問票の回答がこれに影響される可能性があります。
- アーカイブデータ(アーカイブデータ、archival data): 過去に収集され、保管されているデータ。
- 進化論(しんかろん、evolutionary theory): 生物が世代を経て変化していく過程を説明する理論。
- 異文化比較(いぶんかひかく、cross-cultural comparison): 異なる文化を比較することで、人間の普遍性や文化特異性を明らかにする方法。
- 人間の普遍性(にんげんのふへんせい、human universals): すべての文化に共通して見られる特性や行動。
- 文化特異的な修正(ぶんかとくいてきなしゅうせい、culture-specific modifications): 進化したメカニズムが、特定の文化の中で変化した形。
- 種間比較(しゅかんひかく、across-species comparison): 異なる種の間で特性や行動を比較すること。
- 大型類人猿(おおがたるいじんえん、the great apes): チンパンジー、ゴリラ、オランウータン、ボノボなど、人間に最も近縁な霊長類のグループ。
- 精神病理学的兆候や症状(せいしんびょうりてきちょうこうやしょうじょう、psychopathological signs and symptoms): 精神疾患や精神的な異常を示す兆候や症状。
- 正常な心理(せいじょうなしんり、normal psychology): 精神的な健康状態にある人の心の働き。
- コンラート・ローレンツ(Konrad Lorenz): 動物行動学の創始者の一人。
- 行動の病理(こうどうのびょうり、pathologies of behaviour): 異常な行動や病的な行動。
- 生理学的機能(せいりがくてききのう、physiological function): 生物の体内で起こる正常な機能。
- 特定のメカニズム(とくていのメカニズム、specific mechanism involved): 問題となっている心の働きや生物学的過程。
- 有益(ゆうえき、fruitful): 役に立つ、価値がある。
- 見通し(みとおし、prospect): 将来の可能性。
- 無視されてきた(むしされてきた、has greatly been neglected): ほとんど注意が払われてこなかった。
はい、承知いたしました。第1章の「4. 遺伝学」の残りの部分を、高校の生物の教科書のような文体で平易に理解しやすく翻訳し、必要に応じて解説を補足します。
4. 遺伝学
生物を成長させるために必要な情報は、個々のゲノムに保存されています。すべての遺伝情報は、デオキシリボ核酸(DNA)と呼ばれる巨大分子にコード化されています。DNAは、4種類の塩基のうちの1つを運ぶモノマーであるデオキシヌクレオチドからできています。その4種類の塩基とは、アデニン(A)、グアニン(G)、シトシン(C)、チミン(T)です。Aは常にTと、Gは常にCと対になっています。この「アルファベット」または遺伝コードは、すべての生物で同一であり、したがって高度に保存されています。例えば、動物の基本設計、いわゆるバウプラン(ドイツ語の「Plan」は「地図」と訳す方が適切ですが、「体型」と誤訳されています)は、非常に長い期間にわたって保存されてきた調節遺伝子によって維持されています。これらの調節遺伝子は、以下の段落の中心的なテーマである構造遺伝子とは、主に独立して作用します。
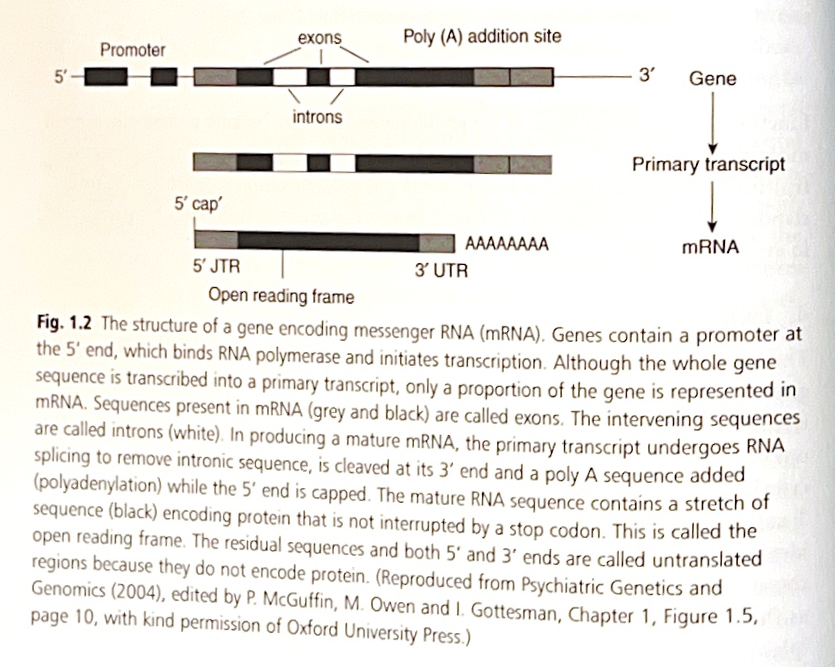
ヒトのDNAは約35億塩基対で構成されています。DNAは、タンパク質の設計図となるコーディングDNAと、それ以外のノンコーディングDNAの混合物です。ヒトゲノムにはおそらく約3万個の機能的な遺伝子が含まれており、これは驚くほど少ない数ですが、動物界ではそれほど例外的ではありません。しかし、コーディングDNAの約55パーセントはヒトの脳で発現しており、これは脳が突然変異の主要な標的であり、それが脳機能を妨げる可能性もそうでない可能性もあることを示唆しています(下記参照)。ヒトDNAの約97パーセントはノンコーディングであり、その機能は完全にはわかっていません。しかし、「ジャンクDNA」と呼ぶのはおそらく不適切です。なぜなら、DNAの生産は目的のない維持には高すぎるコストがかかるため、進化は余分なDNAを排除してきたはずだからです。例えば、ノンコーディングDNAの一部はイントロンで構成されており、遺伝子の開始コドン(エクソン)を隔てるのに役立っています。
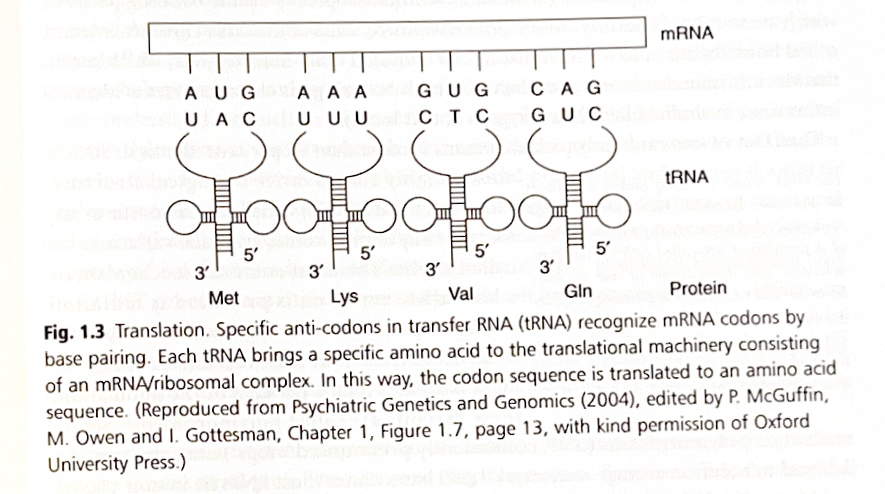
メッセンジャーリボ核酸(mRNA)に転写される鋳型として機能するDNAの単位は、遺伝子と呼ばれます。遺伝子は通常、開始(プロモーター)領域と停止領域で構成されています。これらの領域は、DNAからRNAへの転写プロセスの開始または終了に重要です。遺伝子にはイントロンとエクソンがあり、完全に成熟したmRNAを生成するためには、RNAスプライシングによって一次転写物からイントロンを除去する必要があります。RNAはDNAと同様の構造をしていますが、Tの代わりにウラシル(U)が含まれています。隣接する3つの塩基はトリプレットまたはコドンを形成し、タンパク質を構築するためのアミノ酸をコード化します。4の3乗(64)の可能なトリプレットの組み合わせがあり、20種類の異なるアミノ酸をコード化します。したがって、DNA(RNA)コードは縮重しており、1つのアミノ酸に対して複数の可能なトリプレットの組み合わせが存在します。
通常、DNAは二重らせん構造を取り、ヒトでは23対の染色体の基礎を形成しており、そのうち2対は性染色体XとYです。染色体はサイズ順に番号が付けられており、1番染色体が最も大きいです。48本の染色体を持つ大型類人猿と比較して、ヒトの46本の染色体は数が減少していますが、遺伝情報の量は減少していません。ヒトとチンパンジーの系統が分岐した後のある時点で、ゴリラとチンパンジーに存在する共通祖先の染色体12と13が融合して、ヒトの染色体2を形成しました。哺乳類では、雌の生物は2本のX染色体を持ちますが、雄は1本のX染色体と1本のY染色体を持ちます。Y染色体は、X染色体よりもかなり小さいですが、性を決定します。Y染色体を持つと雄になり、持たないと雌になります(対照的に、鳥類ではXXが雄でXYが雌です)。生物の核型は、常染色体と性染色体の数を別々に示します。したがって、健康なヒトは44 XXまたは44 XYを持ちます。正常な染色体数の変化は、異数性と呼ばれます。最も頻繁な例は、ダウン症候群を引き起こす21トリソミーです。性染色体の異数性は、クラインフェルター症候群(47 XXY)、ターナー症候群(45 X0)、または他の症候群を引き起こす可能性があります。異数性は、しばしば精神遅滞、精神病のリスク増加、または他の行動異常と関連しています。異数性の可能性に加えて、染色体の一部は欠失によって失われたり、逆向きに再挿入されたり(逆位)、重複したり、別の染色体に転座したりする可能性があります。これらの再編成は、精神的な健康を含む発達に深刻な影響を与える可能性があります。
染色体は、セントロメア領域で偏心的に「絞られて」おり、長い腕(q腕)と短い腕(p腕)が生じています。これらの腕には、顕微鏡で見える縞模様があり、セントロメアに近い部分から番号が付けられています。同じ染色体のペアは相同染色体と呼ばれます。「ホモ接合体」と混同してはならない用語です。「ホモ接合体」とは、相同染色体の特定の遺伝子座(ローカス)にある情報が両方とも同一であることを意味します(そうでない場合、その個体はその遺伝子座でヘテロ接合体です)。
ランダムに選ばれた2人のヒトのDNAは、99パーセント以上が同一です。したがって、DNAは高度に保存された生物学的構造です。しかし、理論的には、どの染色体のどの遺伝子座でも変異が発生する可能性があり、そのような変異は対立遺伝子と呼ばれます。対立遺伝子変異が生じる最も一般的なメカニズムは、単一の塩基またはヌクレオチドの置換による突然変異です。特定の遺伝子座における少なくとも2つの対立遺伝子の頻度が集団の1パーセントを超える場合、これらの変異体は多型と呼ばれます。一塩基多型(SNP、都合よく「スニップス」と発音されます)は一般的であり、平均して1,500塩基ごとに1回発生すると考えられています。ほとんどのSNPはノンコーディング領域に存在するため、表現型には影響を与えません。しかし、SNPがコーディング領域、プロモーター領域、エンハンサーまたはサイレンサー領域など、遺伝子発現に影響を与える可能性のあるゲノム領域に位置する場合、劇的な影響を与える可能性があります。同様に、1つ以上の塩基の挿入または欠失(消失)は、対応するゲノム領域の機能に深刻な影響を与える可能性があります。挿入、欠失、重複、および複雑な多部位変異体は、最近「コピー数変異」(CNV)または「コピー数多型」(CNP)という用語で包括されています。そのような変異は、以前に想定されていたよりもヒトゲノムではるかに頻繁に発生します。これまでに1,400以上のCNVを含む領域が特定されており、ヒトゲノムの約12パーセントを占めています。CNVの機能的な重要性とその進化的起源はほとんど理解されていません。しかし、CNV遺伝子座の研究が近い将来、精神医学に影響を与えることが予想されます。いずれにせよ、進化論的な観点からは、わずか1パーセントの適応度の利点をもたらす変異体は、約4,000世代、つまりヒトでは10万年から12万年以内に、集団頻度が0.1パーセントから99.9パーセントに増加することに注目することが重要です。特定の対立遺伝子が適応度の利点をもたらしたかどうかを判断することがいかに難しいかは、以下の例で説明できます。
カテコール-O-メチルトランスフェラーゼ(COMT)のアミノ酸鎖の158位におけるホモ接合体のバリン/バリン(val/val)多型は、ドーパミンの酵素的分解に関与するタンパク質であり、メチオニン/メチオニン(met/met)変異体よりも速く働きます。言い換えれば、met/met変異体はシナプス間隙にドーパミンをより長く残し、それがワーキングメモリに測定可能な影響を与えます。val/valキャリアーは、met/metキャリアーよりもワーキングメモリタスクの成績が悪く、val/val変異体は、特にキャリアーが青年期初期にカンナビスを摂取した場合、統合失調症を発症する脆弱性も伝えることがわかっています。両方のホモ接合体変異体とヘテロ接合体のval/met対立遺伝子は、集団内で平衡状態で存在します。しかし、val/val対立遺伝子はそれ自体が病理ではなく、おそらく十分に理解されていない他の多くの要因と組み合わさってのみ、そのような対立遺伝子変異が統合失調症を説明するわずかな分散に寄与する可能性があります。最近の研究では、実際、met/metキャリアーは、不快な感情刺激に対して辺縁系、前頭前野、側頭領域で反応性が増加しており、ネガティブな気分状態に対する回復力の低下と、より高い感情調節障害を示唆しています。同様に、1つのmet対立遺伝子の存在は、(男性において)強迫性障害のリスク増加も伝える可能性があります。本質的に、これは、COMT遺伝子型変異が、ワーキングメモリと感情反応性に対する有益な効果のトレードオフを表していることを示唆しています。他の遺伝的感受性遺伝子座のそのようなトレードオフについてはほとんど知られていませんが、将来の研究は、特定の障害の「遺伝子」としての地位を早まって割り当てられた他の対立遺伝子のそのような効果をほぼ確実に明らかにするでしょう。
新しい遺伝子は、既存の遺伝子の重複と、祖先遺伝子の近くのゲノムへの挿入によって生じる可能性があります。新しい遺伝子は新しい機能を獲得する可能性があり、元の遺伝子はそのまま残ります。時には、染色体全体またはゲノム全体が重複することもあります。多くの対立遺伝子変異は選択的に中立、つまり個体の繁殖適応度に影響を与えません。遺伝子流動は、遺伝子が集団の遺伝子プールに出入りできるプロセスです。集団が新しい種に進化するのを防ぐため、進化において高度に保守的な要因と見なされています。対照的に、遺伝的浮動は、小さな集団で対立遺伝子がサンプリングエラーによって失われた場合、そのような影響を与える可能性があります。
遺伝子は、エピジェネティックなメカニズムによって差分的に発現またはサイレンスされる可能性があります。遺伝子を不活性化する一般的なメカニズムの1つはメチル化であり、特にシトシンとグアニンヌクレオチドが豊富な領域で発生します。そのようなCpG「アイランド」は、全遺伝子の約50パーセントの一端に存在し、そのうち60〜70パーセントがメチル化されています。ヒトでは、DNA塩基の約1パーセントがメチル化されます。DNAメチル化は、正常な細胞分化における重要なメカニズムですが、腫瘍抑制遺伝子をサイレンシングすることによって発癌にも関与することが示されています。分子生物学の中心的なドグマの1つは、体のタンパク質からDNAへの情報の伝達は不可能であるということです。したがって、獲得形質の遺伝は起こりません。しかし、RNA分子をコード化した元の遺伝子座以外の相同なDNA配列との相互作用を介して、RNAが遺伝子サイレンシングを誘導する可能性があるという発見は、この規則に違反します。そのようなメカニズムがヒトに存在するかどうかは、まだ不明です。
メチル化と遺伝子サイレンシングは、1つのX染色体の不活性化と、ゲノムインプリンティングと呼ばれるプロセスにも関与しています。ゲノムインプリンティングは、雄性または雌性DNAの差分的なメチル化の結果として生じます。これは、胚に存在する父性遺伝子と母性遺伝子の異なる「利益」によって部分的に説明できます。進化論的な観点からは、母親から伝達された遺伝子と父親から伝達された遺伝子の間には、母親の投資量に関して議論の余地のある対立があります。例えば、父性遺伝子は、母親にとっての最適量よりも多くの資源を抽出するように選択された可能性があります。対照的に、母性遺伝子は、母親の最適量に近い、またはそれよりもわずかに少ない量の資源を要求するように選択された可能性があります。インプリントされた遺伝子はゲノムのわずか1パーセントを占めますが、成長調節に関与し、高度に多面的な効果を持つことが多いため、大きな影響を与える可能性があります。インプリントされた遺伝子の標的組織は胎盤であり、そしておそらく脳です。胎盤と脳はどちらも、遺伝的に関連がありながら部分的に異なる適応度の利益を持つ個体間の資源移動のプロセスにおける重要な器官です。したがって、脳におけるインプリントされた遺伝子の不均衡な発現は、機能レベルで大きな変化を引き起こす可能性があると予想できます。この仮説を支持して、プラダー・ウィリー症候群とアンジェルマン症候群は、ゲノムインプリンティングによるそのような反対の効果を反映している可能性があります。一方の親の15番染色体の長腕にある特定のDNA領域の欠失、または両方の染色体領域のコピーが一方の親から伝達された片親性ダイソミーは、母親または父親の遺伝子のみの発現につながる可能性があります。母親由来の遺伝子のみの発現はプラダー・ウィリー症候群を引き起こし、これは乳児期の吸啜不良と弱い泣き声が特徴です。逆に、両方のコピーが父親由来の場合、その結果はアンジェルマン症候群です。アンジェルマン症候群の乳児は、強くはあるが協調性の低い吸啜反応を示します。最近、ゲノムインプリンティングのプロセスは、自閉症と統合失調症において、部分的に社会情報の処理に関連する機能不全の正反対の症状として議論されています。したがって、精神病は、社会認知に関与する特定の脳領域の発達不全につながる母性インプリント遺伝子から生じる可能性があり、一方、父性インプリンティングは、脳の過成長の一般的なパターンを持つ自閉症に関与している可能性があります。これらの両極端な変異は、それぞれ脳の非対称性の減少または増強、ならびに接続性と機能的相関の反対のパターンに反映されています(詳細については、第6章と第10章を参照)。エピジェネティックなメカニズムは、最近、依存症、うつ病、統合失調症を含むいくつかの精神疾患で議論されています。興味深いことに、多くの精神疾患における重要な要因である慢性的なストレスや早期のストレスの多いライフイベントは、メチル化を介した遺伝子サイレンシングに寄与する可能性があります。動物モデルは、これらの通常は長期にわたる効果が薬理学的に可逆的であることを示唆しており、この事実は将来、より大きな注目を集めるでしょう。
非常にまれな疾患、特にハンチントン舞踏病などの常染色体優性遺伝疾患では、疾患の重症度が世代を経るごとに増加し、疾患の発症年齢が若くなることが観察されています。このプロセスはアンチシペーションとして知られるようになり、親から子への伝達中に反復トリヌクレオチド要素の数の増加と関連しています。これが他の精神疾患においてどの程度役割を果たすかは依然として議論の余地がありますが、アンチシペーションは統合失調症、双極性障害、および不安障害で議論されています。
最後に、多くの点で、時には遠く離れた遺伝子間でかなりの相互作用があり、これはエピスタシスと呼ばれるプロセスであることに注意することが重要です。さらに、形質はしばしば複数の遺伝子の制御下にあり、その状態は多遺伝子遺伝と呼ばれます。逆に、複数の表現型側面に影響を与える遺伝子は多面発現性と呼ばれます。これらの事実は、病的な変異の根底にある可能性のある対立遺伝子変異の探索を複雑にしますが、過去数年間に洗練されてきたいくつかの高度な方法が、精神病理への遺伝的寄与の理解を大きく進歩させました。
解説補足
- 構造遺伝子(こうぞういでんし、structural genes): タンパク質をコードする遺伝子。
- ヒトDNA(ヒトDNA、Human DNA): 人間のデオキシリボ核酸。
- 塩基対(えんきつい、base pairs): DNAの二重らせん構造において、対になっている塩基の組み合わせ(A-T、G-C)。
- コーディングDNA(コーディングDNA、coding DNA): タンパク質の設計図となる遺伝情報を持つDNA領域(エクソン)。
- ノンコーディングDNA(ノンコーディングDNA、non-coding DNA): タンパク質の設計図とならないDNA領域(イントロン、遺伝子間領域など)。
- 機能的な遺伝子(きのうてきないでんし、functional genes): タンパク質や機能性RNAをコードする遺伝子。
- 突然変異(とつぜんへんい、mutation): DNAの塩基配列が変化すること。
- ジャンクDNA(ジャンクDNA、’junk’ DNA): かつては機能がないと考えられていたノンコーディングDNAの通称。現在では一部に重要な機能があることがわかっています。
- イントロン(intron): 遺伝子の中で、タンパク質をコードしない挿入配列。転写後、スプライシングによって除去されます。
- 開始コドン(かいしコドン、starting codons): mRNAの翻訳を開始する目印となる3つの塩基配列(AUG)。
- エクソン(exon): 遺伝子の中で、タンパク質をコードする配列。イントロンが除去された後、連結されて成熟mRNAとなります。
- 鋳型(ひなぎた、template): 核酸合成の際に、新しい鎖の配列を決定するために用いられる既存の鎖。
- 転写(てんしゃ、transcription): DNAの遺伝情報がmRNAに写し取られる過程。
- メッセンジャーRNA(mRNA、messenger ribonucleic acid): DNA
はい、承知いたしました。第1章の「4.1. 精神病理への遺伝的寄与の決定」の部分を、高校の生物の教科書のような文体で平易に理解しやすく翻訳し、必要に応じて解説を補足します。
4.1. 精神病理への遺伝的寄与の決定
SNP、複雑な遺伝子相互作用、相加的な遺伝子効果、遺伝子サイレンシングとインプリンティング、多面的な効果やアンチシペーションが精神病理にどのように寄与するかは、ごく最近になって理解され始めたばかりです。個人の遺伝子構成が精神病理への脆弱性と関連している可能性はほとんど疑いの余地がありませんが、遺伝的影響と環境的影響を解きほぐす作業は非常に困難であることが証明されています。これは、主要な精神障害のほとんどすべてが、単純な優性劣性の関係を持つメンデル遺伝のパターンに従わないという事実による部分があります。さらに、遺伝子は可変的に発現する可能性があり、特定の遺伝子型に対する特定の表現型の確率として定義される浸透率が不完全である可能性も考慮に入れる必要があります。同様に、精神病理学的症候群は程度の問題、つまり集団内で量的または連続的に分布しており、「正常」とはカテゴリカルに区別されるものではありません。これらすべてには、多遺伝子遺伝と複数の環境要因によってもたらされる連続的な脆弱性分布を仮定する、数学的に複雑な多因子脆弱性閾値モデルが必要です。
遺伝率は、環境変動と比較して、遺伝的変動によって説明される表現型分散の割合として定義されます。厳密な意味では、遺伝率は集団における遺伝的変動を指し、個々のレベルでは有効な概念ではありません。
さらに、数学的モデルは、共有された環境要因と共有されない環境要因の推定値を考慮に入れる必要があります。
精神病理学的症候群への遺伝的寄与を推定する最も簡単な方法は、家族研究、ならびに双生児研究および養子研究です。これらは、障害の生涯期待値と、近親者間の共通の遺伝子構成の相対的な寄与を推定するために開発されました。家族研究では、生涯に影響を受けた親族の数を親族の総数で割ることによって、障害の生涯有病率を計算できます。ただし、決定された生涯リスクの実際の数値は、実際のリスクよりも低くなります。なぜなら、すべての親族がリスク期間を通過したとは限らないからです。さらに、家族研究は、世代間の生涯リスクを比較するのには適していません。障害の発症年齢に応じて、親族の異なる繁殖成功によって生じるバイアスも存在する可能性があります。双生児研究、特に一卵性(MZ)双生児と二卵性(DZ)双生児の比較は、形質の表現に対する遺伝的および共有された環境的影響の推定を可能にするため、より有益です。養子研究は、障害の遺伝的および環境的原因を区別するのに役立ちます。MZは遺伝物質を100パーセント共有するのに対し、DZは50パーセントしか共有しないためです。MZとDZは、胎内、および子供時代と青年期(一緒に育てられた場合)において同様の程度まで共有された環境にさらされますが、妊娠中の違いは、共通の絨毛膜を共有しないことに起因する可能性があります。一方、兄弟姉妹、さらにはMZでさえ、家族内で異なる「生態的ニッチ」を占めており(兄弟姉妹に関しては、部分的には出生順序効果による)、これは共有されない環境的影響として考慮に入れる必要があります。
いずれにせよ、障害への遺伝的および環境的寄与を測定するためには、生物学的に罹患した親を持つ子供が養子に出され、非罹患家族の養子と比較されたり、罹患した養子が非罹患の養親と比較されたりする養子研究が有益です。養子研究の特別なケースは、罹患した養親を持つ養子の子供を、非罹患の養親を持つ罹患した養子と比較するクロスフォスタリングデザインです。
ただし、遺伝子-環境相互作用と相関関係を解きほぐして定量化することは困難です。受動的な遺伝子-環境相関は、子供が親から遺伝子を受け継ぐだけでなく、特定の遺伝子を持つ親が自身の遺伝子構成に応じて環境を作り出す可能性があるという事実によって発生する可能性があります。例えば、不安な親は、子供にリスクがほとんどまたはまったくない環境を提供しようとするかもしれません。対照的に、注意欠陥/多動性障害(ADHD)の親の子供は、衝動的な反応と感情的な不安定さを促進する家族環境で育つ可能性があります。対照的に、能動的な遺伝子-環境相関は、子供が自身の遺伝子構成に最も適した環境条件を好む可能性があることを指します。例えば、ADHDの子供は、不安な子供よりも危険な状況をより頻繁に求めるかもしれません。したがって、遺伝子-環境相互作用は、遺伝的効果、環境的効果、および遺伝子-環境相関の合計として定義できます。
連鎖解析と関連解析は、SNPを検出したり、ゲノム全体または個々の染色体上に位置する候補遺伝子をテストしたりする方法です。これらは、ポリメラーゼ連鎖反応(PCR)を用いたDNAの指数関数的な複製を利用します。連鎖解析と関連解析の手順は、反対の利点と欠点を持つ点で相補的です。連鎖の方法は、染色体上の2つの点の間で交叉または組み換えが発生する可能性に関係しています。2つの点が近いほど、交叉が発生する可能性は低くなります。交叉が発生する頻度は、種間、雄性配偶子と雌性配偶子間、個々の染色体間、および染色体の異なる部分間で異なります。実験は、ヒトゲノムでは100万塩基対(1メガベース、Mb)のDNAごとに1回の交叉が発生することを示唆しています。染色体上の一つ以上のマーカーの位置がわかっている場合、障害の表現型に寄与する遺伝子は、比較的狭い関心領域に囲い込むことができるかもしれません。連鎖は家族解析で使用され、長い遺伝的距離にわたる大きな効果を検出できます。連鎖解析は、元々メンデル遺伝に従う障害に適用されましたが、量的形質を調べるための複雑な統計的手順が利用可能になり、これはほとんどの精神障害にとってはるかに適切です。関連研究は、遺伝的に無関係なサンプルに適用できます。最も単純な比較は、障害を持つ個人と適合する対照被験者との間で行われます。特定の染色体遺伝子座における特定の対立遺伝子と障害の関連性は、因果関係を示唆する可能性があります。連鎖不平衡(LD)は、異なる遺伝子座にある2つの対立遺伝子がどれだけ密接に連鎖しているかの尺度です。高いLDは、2つの対立遺伝子が一緒に伝達されることを示唆していますが、同じ距離の遺伝子座間でも大きなばらつきがあるため、関連は短い距離でのみ意味があります。一方、関連研究は、比較的統計的検出力の小さい遺伝的効果を検出できます。関連研究は、アポリポタンパク質多型がアルツハイマー病の危険因子であることや、ドーパミントランスポーター遺伝子多型がADHDの危険因子であることなど、既知の(病態)生理学的効果を持つ候補遺伝子の対立遺伝子変異を特定するのに最も成功しています。
複雑な精神障害の定量的遺伝学は、かなりの進歩にもかかわらず、まだ初期段階にあります。これは、異なる遺伝子座にある複数の対立遺伝子の複雑な相互作用によるものであり、その結果、個々の発見が全体的な集団リスクを説明する力が小さいため、発見の再現がしばしば失敗しています。現在、複雑な形質は単一の遺伝子ではなく、複数の遺伝子によって遺伝し、それぞれの遺伝子が実際の表現型にわずかな分散を寄与する(非遺伝的影響に加えて)ことが広く合意されています。複数の遺伝子システムにおける遺伝子は、量的形質遺伝子座(QTL)と呼ばれており、各QTLが形質の変動に寄与する程度は、ほとんどの特性で不明です。
別の問題は、異なる民族的背景を持つサンプルの比較可能性にあります。ただし、最も差し迫った困難は、精神障害の診断基準が、特定の表現型に寄与する遺伝子を特定するには曖昧すぎるツールであることです。ましてや、精神障害が健康な状態が統計的規範を表す症状の重症度の連続体における極端な点として存在するという問題はなおさらです。これらすべては、共有された生物学的または行動学的マーカーに基づく内表現型の徹底的な特徴付けを不可欠にします。内表現型は、障害の完全な表現型変動と比較して、遺伝的に複雑でないと想定されています。理想的には、内表現型は遺伝性であり、したがって、複数の個人が罹患している家族で疾患と共分離します。それは、一般集団よりも高い頻度で、指標例の非罹患親族に見られます。内表現型は状態非依存性、つまり疾患活動によって変動しません。
別の有望な方法は、動物(例えば、マウス)DNAにヒトDNAを組み込むことであり、少なくともいくつかの行動的または生理学的相関関係がモデル動物で観察されることが期待されます。
ただし、多くの精神障害の感受性遺伝子の探索がこれまで挫折感を伴うものであった理由の1つは、人間の行動が非常に複雑であり、おそらく何百、何千もの遺伝子によって影響を受けているという事実にある可能性があります。しかし、進化論的な観点からは、精神障害が一般集団で非常に一般的な理由の説明が必要です。高い有病率は、むしろ、精神障害を起こしやすい遺伝子が選択によって排除されたことを示唆するでしょう。一つの説明は、精神障害を起こしやすい遺伝子が、おそらくバランスの取れた多面的な効果を通じて、人間の進化の歴史の中で何らかの隠れた適応上の利点を持っていた可能性があるということです。統合失調症を含むいくつかの精神障害は、ヘテロ接合体キャリアーではマラリアからの優れた保護に関連し、ホモ接合体個体では有害である鎌状赤血球貧血の例に例えられてきました。そのようなシナリオは統合失調症では非常にありそうもないことが証明されていますが、このテーマの変形は、人生の早期に適応上の利点をもたらす遺伝子が、人生の後半で有害な影響を与える可能性があることを示唆しています。そのようなメカニズムは、選択圧の力が通常低下する生殖期以降の遅発性障害にとってあり得るかもしれません(例えば、認知症に関する第8章を参照)。バランス選択は、2つの対立遺伝子が進化の時間を通じて適応度の点で等しく重要であったと仮定します。
バランス選択の特別なケースは、頻度依存性選択であり、特定の対立遺伝子によってもたらされる適応度が、その対立遺伝子がまれになるにつれて増加する場合に発生します。そのようなメカニズムは、低頻度での特定の形質の維持を簡潔に説明でき、例えば、「精神病質」として知られる反社会的行動傾向を説明するモデルとなる可能性があります(詳細については、第14章を参照)。
対照的に、(現代の精神遺伝学で暗黙のうちに、ただし誤って一般的になっている仮定である)精神障害の感受性を高める対立遺伝子が選択的に中立であるという可能性は、多くの精神障害に関連する重大な繁殖上の不利益を考慮すると、ありそうにありません。選択によって精神障害が排除されない別の説明は、対立遺伝子変異の単なる合計が、集団内の任意の遺伝子座ではまれであるが、集合的には非常に一般的な対立遺伝子が、個体を精神障害を発症しやすくするということです。最近発表された「分水嶺モデル」は、特定の遺伝子座におけるいくつかまたは多くの不利なSNPが、連動して機能不全を引き起こす可能性を示しています。COMT遺伝子型のval/val変異体は、実際には前頭前野におけるドーパミン代謝回転率を高める可能性があり、これおよび他の相加的なメカニズムは、met/met遺伝子型のキャリアーよりもわずかに低いワーキングメモリ機能を寄与する可能性があります。ワーキングメモリ機能は、他のメカニズムとともに、認知表現型に影響を与える可能性があります。ただし、これらのメカニズムのそれぞれは、ノイズ以上のものをほとんど生成しません。十分なノイズが存在する場合、閾値に達し、表現型的に関連する症状または症候群を引き起こし、最終的には個体の適応度に影響を与える可能性があります。各個人のヒトゲノムに存在する可能性のある有害な対立遺伝子の数の推定値は、500から2,000に達します。これと、遺伝子がタンパク質をコードし、認知、感情、または行動に直接翻訳されないという事実は、「特定の障害の遺伝子」が存在しないことを示唆しています。環境要因との密接な相互作用においてのみ、遺伝子は特定の表現型を生み出します。精神病理を完全に理解したいのであれば、これらの重要な洞察を考慮に入れる必要があります。
遺伝学を含む精神障害を理解するための大きな機会は、ティンバーゲンとローレンツが非常に巧妙に想定したように、近接レベルと究極レベルの両方における精神病理の進化的概念化の基礎にあります。
解説補足
- SNP(スニップ、Single Nucleotide Polymorphism): 一塩基多型。ゲノムDNA配列における一個のヌクレオチドの変異。
- 遺伝子相互作用(いでんしそうごさよう、gene interactions): 複数の遺伝子が互いに影響を与え合い、表現型を決定する現象。
- 相加的な遺伝子効果(そうかてきないでんしこうか、additive gene effects): 複数の遺伝子がそれぞれ独立に表現型に影響を与え、その効果が足し合わされる現象。
- 遺伝子サイレンシング(いでんしサイレンシング、gene silencing): 特定の遺伝子の発現が抑制される現象。
- 遺伝子インプリンティング(いでんしインプリンティング、gene imprinting): 親由来によって遺伝子の発現が異なる現象。
- 多面的な効果(ためんてきなこうか、pleiotropic effects): 1つの遺伝子が複数の異なる表現型に影響を与える現象。
- アンチシペーション(anticipation): 遺伝性疾患において、世代を経るごとに発症年齢が若くなり、症状が重くなる現象。
- メンデル遺伝(メンデルいでん、Mendelian inheritance): 単一の遺伝子の変異によって引き起こされ、単純な優性劣性の法則に従う遺伝形式。
- 浸透率(しんとうりつ、penetrance): 特定の遺伝子型を持つ個体が、対応する表現型を示す確率。
- 精神病理学的症候群(せいしんびょうりてきしょうこうぐん、psychopathological syndromes): 特定の精神障害に見られる症状のまとまり。
- 量的形質(りょうてきけいしつ、quantitative trait): 身長や体重のように、連続的な値で表現される形質。多くの精神障害は量的形質として捉えられます。
- 多因子脆弱性閾値モデル(たしいんしぜいじゃくせいきいちモデル、multifactorial liability-threshold models): 遺伝的要因と環境的要因が累積的に個体の脆弱性を高め、ある閾値を超えると障害が発症すると考えるモデル。
- 遺伝率(いでんりつ、heritability): 集団における表現型のばらつきのうち、遺伝的な要因によって説明される割合。
- 表現型分散(ひょうげんがたぶんさん、phenotypic variance): 集団における表現型のばらつき。
- 遺伝的変動(いでんてきへんどう、genetic variation): 集団における遺伝子型のばらつき。
- 共有された環境要因(きょうゆうされたかんきょうよういん、shared environmental factors): 家族内で共有される環境要因(例:家庭環境、育児スタイル)。
- 共有されない環境要因(きょうゆうされないかんきょうよういん、non-shared environmental factors): 家族内で共有されない個別の環境要因(例:友人関係、学校生活)。
- 生涯期待値(しょうがいきたいち、lifetime expectations): ある年齢までに特定の状態を発症する確率。
- 生涯有病率(しょうがいゆうびょうりつ、lifetime prevalence): 生涯のある時点で特定の疾患を経験した人の割合。
- 一卵性双生児(いちらんせいそうせいじ、monozygotic twins – MZ): 一つの受精卵が分裂してできた双子で、遺伝子を100%共有します。
- 二卵性双生児(にらんせいそうせいじ、dizygotic twins – DZ): 異なる2つの受精卵から生まれた双子で、遺伝子を平均50%共有します(通常の兄弟姉妹と同じ)。
- 養子研究(ようしけんきゅう、adoption studies): 生物学的な親と育ての親の影響を比較する研究。
- クロスフォスタリングデザイン(クロスフォスタリングデザイン、cross-fostering design): 疾患を持つ養親に育てられた養子と、疾患を持たない養親に育てられた疾患を持つ養子を比較する研究。
- 遺伝子-環境相互作用(いでんし-かんきょうそうごさよう、gene-environment interactions): 遺伝的要因と環境的要因が互いに影響し合い、表現型を決定する現象。
- 受動的な遺伝子-環境相関(じゅどうてきないでんし-かんきょうそうかん、passive gene-environment correlations): 子供が親から遺伝子を受け継ぐだけでなく、親が自身の遺伝子型に基づいて環境を作るために生じる相関。
- 能動的な遺伝子-環境相関(のうどうてきないでんし-かんきょうそうかん、active gene-environment correlation): 子供が自身の遺伝子型に合った環境を好んで求めるために生じる相関。
- 連鎖解析(れんさかいせき、linkage analysis): 家族内で疾患と特定の遺伝子マーカーが一緒に遺伝する傾向を調べることで、疾患の原因遺伝子の染色体上の位置を特定する方法。
- 関連解析(かんれんかいせき、association analyses): 集団内で疾患を持つ人と持たない人で、特定の遺伝子多型の頻度に違いがあるかを調べることで、疾患と関連する遺伝子を特定する方法。
Selected further reading
Allman, J. 1999, Evolving Brains, Scientific American Library, New York.
Bowlby, J. 1969, Attachment and Loss. Vol. 1. Attachment, Basic Books, New York.
Buss, D. M. 1999, Evolutionary Psychology. The New Science of the Mind, Allyn and Bacon, Boston.
Darwin, C. 1859, On the Origins of Species by Means of Natural Selection, Murray, London.
Darwin, C. 1871, The Descent of Man, and Selection in Relation to Sex, Murray, London.
Dawkins, R. & Krebs, J. R. 1979, ‘Arms races between and within species’. Proceedings of the Royal Society
of London Series B, vol. 205, pp. 489-511.
Gilbert, P. 1998, ‘Evolutionary psychopathology: why isn’t the mind designed better than it is?’ British
Journal of Medical Psychology, vol. 71, pp. 353-373.
Hrdy, S. B. 2000, Mother Nature, Vintage, London.
Keller, M. C. & Miller, G. in press, ‘Resolving the paradox of common harmful, heritable mental disor-
ders: Which evolutionary genetic models work best?’, Behavioral and Brain Sciences.
Lorenz, K. 1973, Die Rückseite des Spiegels: Versuch einer Naturgeschichte menschlichen Erkennens,
Piper, München.
Mayr, E. 2001, What Evolution Is, Basic Books, New York.
McGuffin, P., Owen, M. J. & Gottesman, I.I. (eds) 2005, Psychiatric Genetics and Genomics, Oxford
University Press, Oxford.40
Ridley, M. 2004, Evolution. 2nd ed., Oxford University Press, Oxford.
Tinbergen, N. 1963, ‘On aims and methods of ethology’, Zeitschrift für Tierpsychologie, vol. 20,
pp. 410-433.
Tooby, J. & Cosmides, L. 1990, ‘The past explains the present. Emotional adaptations and the structure
of ancestral environments’, Ethology and Sociobiology, vol. 11, pp. 375-424.
Trivers, R. L. 1971, The evolution of reciprocal altruism. The Quarterly Review of Biology, vol. 46,
pp. 35-57.
Trivers, R. 1972, ‘Parental investment and sexual selection’ in Sexual Selection and the Descent of Man,
ed. B. Campbell, Aldine-Atherton, Chicago, pp. 136-179.
Trivers, R. L. 1974, ‘Parent-offspring conflict’, American Zoologist, vol. 14, pp. 249-264.
Williams, G. C. 1966, Adaption and Natural Selection, Princeton University Press, Princeton, NJ.
サイドノートまとめ
近位メカニズム(個体発生と生理学)と究極原因(系統発生と適応機能)は、行動の理解に不可欠と考えられる補完的な次元である。
精神病理学的兆候と症状は、適応形質の機能不全的な極端な変異を構成するという理解の下で、同様に分析することができる。
自然選択と性選択による進化は、人間の認知、感情、行動を形作ってきた歴史的なプロセスである。
安定化選択は、交配集団内の個体間の差異を減少させる傾向があるが、個体間のわずかな差異でさえ、異なる繁殖成功に寄与する。
ほとんどの突然変異は有害である。したがって、選択は主に除去のプロセスである。しかし、まれに、正の選択が集団内でそれぞれの形質の急速な広がりにつながることがある。
選択は主に個体レベルで行われる。ほとんどの適応は設計によって最適ではなく、妥協の産物である。これは、一部の不適応な形質(精神病理に関連するものを含む)が選択によって除去されない理由を部分的に説明する。
ヒトにおいては、個体選択と群選択の両方が、協力、道徳、社会認知能力の進化に寄与した可能性がある。
自然選択と性選択は、種の進化を駆動する2つの別々のプロセスである。種間および種内の「軍拡競争」は、適応的修正の速度に寄与する。
性選択は、チャールズ・ダーウィンにとって長年の謎であった。なぜなら、クジャクの尾のような一部の形質は、明らかに生存の点で適応上の不利益をもたらすからである。性的に選択された形質は、「遺伝的浮動」によって出現したり、優れた遺伝的品質を示す可能性がある。
肥沃な雌へのアクセスを巡る雄間の同性内競争は、体の性的二型と精子競争の進化につながった。対照的に、ほとんどの種(ヒトを含む)では、雄の配偶者としての質を評価に基づいて配偶者を選ぶのは雌である(雌の選択原理)。
包括適応度理論は、個体の適応度は、その個体自身の繁殖成功と遺伝的に関連する個体の繁殖成功の合計であると示唆する。
互恵的利他主義の理論は、遺伝的に関連のない個体間の利他主義は、利他的行動が報復され、非協力的な行動を検出し報復を強化する適切なメカニズムが存在する場合にのみ持続すると提唱する。
ヒトにおいては、同情、恥、罪悪感などの社会的な感情のいくつかは、おそらく互恵性に関連する問題への対応として進化した。
迫害妄想のような精神病理学的症候群は、欺瞞行動の検出に関与するメカニズムの極端な変異と見なすことができる。対照的に、精神病質は、自身の利益のために他人を欺くことを過度に行う行動傾向を反映している。
有性生殖は、無性生殖よりも、有害な突然変異の蓄積を相殺し、急速に進化する病原性細菌からの脅威をより良く防ぐという利点がある。
哺乳類では、雌は雄よりも潜在的な子孫にはるかに多く投資する。その結果、雌は配偶者選択においてより識別力が高い(雌の選択原理)。
霊長類(ヒトを含む)では、繁殖成功は、延長された幼年期、遅い繁殖、および長い出産間隔によって制約されており、他の多くの哺乳類と比較して、父性投資の増加を促進する。
ヒトにおける比較的大きな雄と雌の親投資は、父性の確保と感情的なコミットメントを確実にするための嫉妬のような感情的な適応を選択してきた。
穏やかな性的二型、小さな精巣サイズ、および隠蔽された排卵は、祖先条件下のヒトが、一夫一婦制または穏やかな一夫多妻制が一般的であった社会で生活していたという仮説を支持する。
妄想性嫉妬や色情狂のような精神病理学的症候群は、ヒトの配偶行動に関与する適応メカニズムの歪みである。
共有された遺伝物質の量のために、親世代と子世代の間の対立は、親の側の親投資を制限し、子の側の親投資を増加させるためのさまざまな適応を生み出してきた。
親子間の対立は、子宮内環境(エネルギー供給量について)から青年期および成人期初期(子の配偶者選択について)まで、子の異なる発達段階で生じうる。哺乳類に典型的な親子間の対立は、離乳である。
「退行」は、親投資を増やそうとする試みとして、子に典型的に見られる。
親子間の対立の特別なケースは、子の性別に応じた異なる親投資に関連する。良好な環境条件下では、親は雌と比較して雄の繁殖能力が高いため、雄の子を好む傾向があるかもしれない。不良な環境条件下では、環境条件が雌に与える影響が小さいため、雌の子への投資を好むかもしれない。
霊長類とヒトは、出生後まで親投資に関する決定を延期してきた。新生児殺しは、狩猟採集民や現代社会においてさえ、未婚の十代の母親が新生児を遺棄することがあるなど、産児制限の方法であった。
進化心理学の前提は、ヒトの認知、感情、行動は、解剖学と本質的に同じ方法で、自然選択と性選択によって形作られてきたということである。
ほとんどのヒトの心理的メカニズムは「開かれたプログラム」であり、適切に発達するためには適切な環境刺激に大きく依存する。したがって、不十分な環境入力は機能不全を引き起こす可能性がある。
多くの進化した心理的メカニズムは領域特異的であり、経済的、信頼性が高く、効率的で、正確な方法で、特定の種類の情報のみを処理する。進化した心理的メカニズムのモジュールは階層的に組織化されており、トリガーアルゴリズムによって活性化される。
心理的メカニズムは、必ずしも論理を伴わない、適応上の重要性の高い問題の解決に偏っている。
ケアを引き出す、ケアを与える、配偶、同盟形成などのヒトの生物社会的目标は、個人の実際の行動を導く。
進化した心理的メカニズムは設計によって最適ではなく、それが機能不全を起こしやすい理由の一つである。
ヒトの精神性の多くの側面は、狩猟採集生活という祖先環境条件下で進化した。ヒトは約150人の個人的な知り合いがいる、小さく緊密な血縁に基づいたコミュニティで生活するように適応している。
群内協力と群間競争(戦争を含む)は、ヒトの進化の長い期間にわたって一般的であった。
最新の適応に先立ち、視覚野の狭窄が社会集団の形成を必要としたため、双眼視が霊長類の社会性の進化に貢献した可能性がある。
すべての人類は遺伝的に密接に関連している。ヒトの進行中の適応的修正を示す兆候があり、その一部は言語と文字の進化に関連している。
進化心理学的理論から導き出された仮説の経験的検証は、進化論から直接仮説を導き出すトップダウンアプローチと、観察主導型のボトムアップアプローチの2つの方法で行うことができる。
すべての遺伝情報は個人のDNAに保存されており、DNAはアデニン、グアニン、シトシン、またはチミンを運ぶデオキシヌクレオチドで構成されている。生命の遺伝コードは、進化の歴史を通じて高度に保存されてきた。
ヒトゲノムは約3万個の機能遺伝子を含むと考えられている。遺伝子は、メッセンジャーRNAが転写される単位または鋳型を表す。コーディングDNAの約55パーセントはヒトの脳で発現しており、脳が突然変異選択プロセスの主要な標的となっている。
ヒトは23対の染色体を持っており、DNAを運んでいる。これらの染色体のうち2つ、XとYは性染色体と呼ばれる。染色体数の異常である異数性は、しばしば行動異常を引き起こす。
ランダムに選ばれた2人のヒトのDNAは99パーセント同一である。集団の1パーセント以上に関連する単一ヌクレオチドの変異はSNPと呼ばれる。ゲノムのコーディング領域に位置する場合、1つ以上の塩基の挿入、欠失、および重複はかなりの表現型効果を発揮する可能性がある。そのようなコピー数変異(CNV)の機能的意義と進化的起源は、ほとんど不明である。
ヘテロ接合性多型がそのホモ接合性対応物よりも選択的に有利な場合、「バランス多型」と呼ばれる。
遺伝子はエピジェネティックなプロセスによってサイレンスされる可能性があり、その中でメチル化が最も一般的なメカニズムである。メチル化は、雌性生物における1つのX染色体のサイレンシングに寄与する。
ゲノムインプリンティングは、雌性または雄性DNAの異なるメチル化の結果として生じる。遺伝子のわずか1パーセントしかインプリントされていないにもかかわらず、インプリントされた遺伝子は、母親または父親から伝達されたかどうかによって深刻な影響を与える可能性がある。
プラダー・ウィリー症候群とアンジェルマン症候群は、ゲノムインプリンティングの既知の例である。インプリンティングは、自閉症や統合失調症でも議論されている。
アンチシペーションとは、反復遺伝要素の数の増加が、疾患の発症をより若い年齢にシフトさせる可能性があるという観察を指す。アンチシペーションはハンチントン舞踏病で知られており、統合失調症、双極性障害、および不安障害で議論されている。
遺伝子は、エピスタシスと呼ばれる複雑な方法で相互作用する可能性がある。さらに、複数の遺伝子の制御下にある形質は多遺伝子性と呼ばれる。逆に、複数の形質に影響を与える遺伝子は多面的と呼ばれる。
精神障害の遺伝的背景は多遺伝子性である。
遺伝率は、遺伝的変動によって説明される表現型分散の割合として定義される。
家族研究、双生児研究、および養子研究は、疾患を発症する生涯リスクを推定するために使用されてきた。一卵性(MZ)双生児と二卵性(DZ)双生児の研究により、形質の表現に対する遺伝的および共有された環境的影響の推定が可能になる。養子研究は、疾患の遺伝的原因と環境的原因を区別するのに役立つ。
遺伝子-環境相互作用は、遺伝的効果、環境的効果、および遺伝子-環境相関の合計である。
連鎖研究と関連研究は、ゲノムワイドスキャンでSNPまたは候補遺伝子を検出するのに適している。連鎖不平衡(LD)は、異なる遺伝子座にある2つの対立遺伝子がどれだけ密接に連鎖しているかの尺度である。
内表現型の研究は、疾患の完全な表現型変動を研究するよりも有望である可能性がある。内表現型は、理想的には遺伝性であり、疾患の活動とは独立しており、非罹患親族に見られる生物学的および/または行動学的マーカーによって特徴付けられる。
精神障害は一般集団で一般的である。なぜなら、それらはそれぞれ表現型変動にわずかな影響しか与えず、したがって選択による除去を免れる多くの遺伝子の相加効果を反映している可能性があるからである。
