
第3章
人間の生活史
1. 序論
生活史理論は、個体の生涯にわたって変化する、生存と繁殖に関連する問題に対する、種に典型的な解決策を扱います 。
K選択種は、ゆっくりと発達し、繰り返し繁殖し(反復産卵性)、一度の出産での子どもの数は少なく、長い妊娠期間を含む長期にわたる集中的な親の投資を行い、乳幼児期の死亡率は低いです 。
生殖活動の開始は遅く、K選択種は長寿命を享受します 。
一般的に、K選択種は大きな体を持ち、K選択種の成体間の競争は激しいです。
なぜなら、寿命が長いため、競争が生存と繁殖の成功を決定するからです 。
K選択種は通常、ランダムな環境変動が少ない条件で生息しています。
逆に、r選択は反対のパターンを含みます 。
r選択種は、急速に発達し、生涯に一度だけ繁殖することが多く(単回繁殖性)、子どもの数は多いですが(非常に多い)、成体の段階まで生き残るのはごく一部です 。
r選択種では、出生(または孵化)後の親の投資は少ないか、または全くなく、性的成熟は早く始まり、個体は早く死にます 。
r選択種は通常、小さな体を持ち、r選択種ではランダムな環境変動が大きいです 。
K選択種の子どもは、出生時に比較的成熟していますが(早成性)、r選択種では、子どもはしばしばはるかに未熟です(晩成性) 。
哺乳類の中で、多くのげっ歯類は一般的にr選択的な生活史パターンに従う一方で、霊長類は非常にK選択的な種です 。
特に、類人猿とヒトは、極端なK選択的な生活史で際立っており、この発達パターンは、個体の生涯にわたる変化する要求とともに、種の心理的な構造に深い影響を与えます 。
興味深いことに、ヒトの極端なK選択は、おそらく、異時性的な幼形進化とそれに続く過形性の過程と密接に結びついています。
過形性は、おそらく、ヒトを精神病理を発症しやすくするでしょう 。
ホモ・サピエンスが、大きな人間の脳を含むK選択的な特徴を獲得した最も重要なメカニズムです。
人間の生活史の最も際立った特徴は、典型的な霊長類のパターンが発達のすべての段階で延長されていることです。
しかし、この章で後述するように、生活史パターンには種内でのばらつきがあり、一部の人間の生活史は、r選択的なパターンにわずかに近づいているように見えるものもあれば、より極端なK選択的なパターンに近づいているように見えるものもあります。
r選択的ではないパターンが、精神病理の発達の素因となるかどうかは、現在のところ不明ですが、多くの実証的な発見は、この仮説を支持するものと解釈できます。
- 乳幼児期
生物学的に言えば、すべての新生児にとって最も重要な課題は、性的成熟に達するために、乳幼児期をなんとか生き延びることです。
人間の赤ちゃんも、この点で例外ではありません。
しかし、彼らは出生時に極めて未熟であるという問題に直面しています。
出生時の未熟さは、二足歩行による骨盤の解剖学的構造の変化に関連する、設計上の妥協を表しています。
人間の進化における脳の拡大は、早期の分娩への圧力を高めました。
他の類人猿と比較すると、例えば、人間の新生児は、生理学的に約13ヶ月早く生まれています。
言い換えれば、もし人間の赤ちゃんが、チンパンジーの赤ちゃんと同じくらい成熟した状態で生まれてくるなら、人間の妊娠期間は約22ヶ月続くことになります。
人間の早期分娩は、進化的な設計上の妥協を表しており、それは最終的に二足歩行の進化と関連しています。
直立歩行は、人間の骨盤の解剖学的構造の変化を伴い、その結果、産道が狭くなりました。
祖先のヒト上科の種は、約350 cm³の比較的小さなチンパンジーのような脳のサイズ、つまり小さな頭蓋骨を持っていたので、これは問題ではありませんでした。
しかし、進化の過程で脳(と体)が大きくなり始めると、狭い産道の問題はより重大になりました。
人間の祖先の種における脳の拡大は、おそらく不連続に起こりました。
脳の最初の成長の急増は、約180万年前に始まり、おそらく、より多くのタンパク質を摂取する食生活の変化によって促進され、ホモ・ハビリスにおいて脳のサイズが350から800 cm³に倍増しました。
祖先のヒト上科の種は、ずっと以前から直立歩行をしていたことに注意してください。
したがって、二足歩行と脳の拡大は、以前考えられていたほど密接に関連していません。
脳のサイズの2回目の「飛躍」は、約40万年前に起こり、その時、古人類のホモ・サピエンスが進化し、平均脳サイズが1,300 cm³になりました。
この脳のサイズの2回目の増加は、おそらく、人間の社会脳を生み出す社会構造の変化と関連していました(第2章の考察を参照)。
これらの連続的な脳のサイズの増加は、出産を早めるように選択し、その結果、出生時の子どもの未熟さが増しました。
それにもかかわらず、この進化した設計上の妥協にもかかわらず、人間の出産プロセスは、母親と赤ちゃんの両方にとって、非常に危険な試みとなっています。
産道を通過するために、人間の赤ちゃんは頭、体、四肢を2回ねじる運動をしなければならず、そのため、出産は赤ちゃんとお母さんの両方を、起こりうる合併症の広い範囲から死亡する大きな危険にさらします。
図3.1 人間の出産の段階
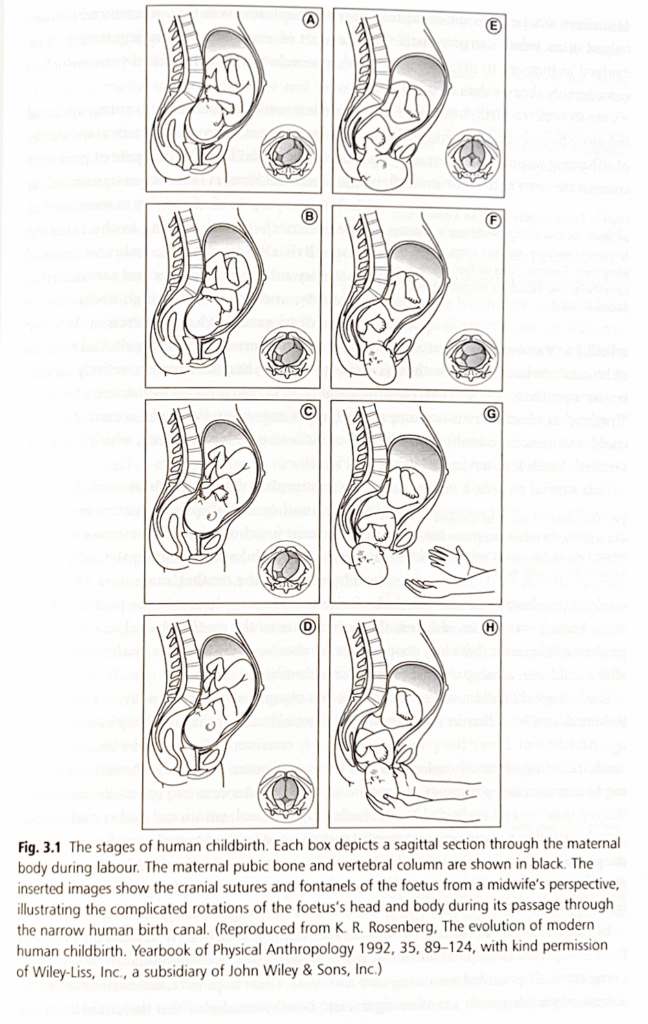
各ボックスは、分娩中の母親の体の正中矢状断面を示しています。
母親の恥骨と脊柱は黒で示されています。
挿入された画像は、助産師の視点から見た胎児の頭蓋縫合線と泉門を示しており、狭い人間の産道を通過する間の胎児の頭と体の複雑な回転を示しています。
(K. R. Rosenberg, The evolution of modern human childbirth. Yearbook of Physical Anthropology 1992, 35, 89-124, Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.のご厚意により転載)
助産術は、おそらく初期の人類における最初の職業であり、経験豊富な女性が他の女性の出産を助けました 。
これは、出産間隔を他の類人猿よりもかなり短い約3年に短縮するために、人類で進化した協力繁殖戦略の一部でした 。
早産のため、人間の新生児は若い類人猿と比較して早成性が低く、したがって二次的にいくらか晩成的な特徴を保持しています 。
早成性とは、生まれた直後に自分の足で動く能力を指します 。
早成動物の目と耳は、出生時またはその後すぐに開いています 。
人間の赤ちゃんは、出生時に目と耳が開いている、母親と交流し、運ぶ人にしがみつく能力などの早成的な特徴を示します 。
対照的に、晩成性は、出生時に目と耳が閉じている、不動性、無毛などの未熟さの兆候を含み、集中的な親のケアが必要です 。
巣、穴、または巣穴での発達期間が必要です 。
自分で動き回ったり、積極的に快適さや避難場所を求めたりすることができないことは、人間の新生児の晩成的な特徴です 。
しがみつくための毛皮がないため、人間の赤ちゃんは母親によって積極的に運ばれます 。
ドイツ語では、人間の新生児のために導入された生物学的用語は「Tragling」であり、これに相当する英語の表現はありません(「運ばれる者」) 。
もちろん、これは二足歩行と組み合わせてのみ可能になり、それによって母親の手が子ども(と食料)を運ぶために解放されました。
人間の新生児のこの特別な状況は、母親と子どもとの間のダイアドの形成が、初期の人類における最も重要な心理的適応の1つになったことを意味します。
幼児の愛着行動による母親と子どもとの間のダイアドの形成は、初期の人類における最も重要な心理的適応の1つになりました。
赤ちゃんの側の愛着と母親側の絆は、心理的な幸福のために、人類の進化を通して今日まで、すべての既知の文化において不可欠なままでした 。
さらに、母親への協力者の依存は比例して増加し、女性間の緊密な協力、子どもを持つ母親と父親間の協力、および血縁関係の同盟が積極的に選択されました 。
愛着の生物学的機能は、ジョン・ボウルビィによって最初に認識され、彼自身とその追随者によって、幼児期から老年期までの普遍的な人間の行動の理論に発展しました 。
愛着理論は、人間の霊長類、人間の正常な愛着と絆、および里親養育や、身体的、感情的、性的虐待を含む幼児期および幼年期の他の逆境などの病理学的状態に関する実証的研究を含む、進化論的な枠組みと完全に一致することが証明されています 。
愛着理論は、安全な愛着と不安な愛着の心理的、生理学的、および行動的結果について予測を行うために使用できる強力なツールを表しています 。
これらの予測は経験的検証に開かれており、愛着研究に関連する複雑な遺伝子-環境相互作用は、解明され始めたばかりです 。
当初、ボウルビィは、里親の家で育った多くの若い男の子が、定期的に基本的なケアと食料を提供されていたにもかかわらず、非行や反社会的な行動を示し、感情的に不器用であったという事実に衝撃を受けました 。
しかし、これらの男の子は、母親像との密接な関係を奪われており、ボウルビィは、これが彼らの行動問題の中核にある可能性があると結論付けました 。
人間の霊長類以外に関する研究は、後に、若いサルと生物学的または代理の母親との親密さの顕著な重要性を確認しました 。
愛着の重要性は、人間の霊長類以外に関する多くの研究で実験的に示されています。
母親から幼い頃に引き離され、社会的孤立の中で育てられた新生児のサルは、ミルクを提供するメッシュで覆われたダミーの母親よりも、毛皮で覆われたダミーの母親にしがみつくことを好みます 。
介護者からの孤立の期間に応じて、サルは重度のうつ病を発症します 。
大人になると、これらのサルは社会的に無能になり、自分自身の子どもを育てることができません 。
これら多くの点で無慈悲な実験は、その後の生活史段階全体を通して、心理的な幸福と社会的有能さにとって、人生の早い段階での母親像との社会的親密さの重要性を明確に確認しました 。
生活史と、同種の主に辺縁系の脳構造に表される進化した心理的愛着メカニズムの大きな類似性を考慮すると、人間の健全な発達は、おそらく出生前から、非常に早い時期からの近さと感情的な温かさに決定的に依存していることは明らかです(この章の考察を参照) 。
人間を含む若い霊長類は、実際に近さを求めるように生物学的にプログラムされています 。
愛着行動は、幼児の生存の可能性を高め、社会的学習を促進するために選択されました 。
たとえば、人間の新生児の体温調節システムはまだ未熟であり、脱水症状を起こしやすい可能性があります 。
これは、部分的には、毛皮による断熱がないこと(ただし、皮下脂肪層によってある程度補償されています)、および総エネルギー摂取量のほぼ3分の2が発達中の脳によって消費されるという事実によるものです 。
注目すべきことに、人間の出産が生理学的に早いため、脳は出生後も出生前と同じペースで1年以上成長し続けます 。
その後初めて、脳のエネルギー消費量は、体のエネルギー需要と比較してゆっくりと減少します 。
この成長パターンは、出生後すぐに脳が体の成長と比較してゆっくりと成長し始める他の霊長類の成長パターンとは異なります(図2.3を比較) 。
ただし、すでに指摘したように、愛着形成は、母親と赤ちゃんの間の強い愛情の絆が栄養失調から最もうまく保護する可能性があるにもかかわらず、主に摂食の必要性や欲求とは関連していません 。
幼児の主な目標は、親密さと安全を求めることです。
人間の幼児は、これを達成するためのいくつかの手段を持っています 。
生後4週で、人間の幼児はすでに人間の顔の輪郭を強く好むようになります 。
彼らは人間の声、特にお母さんの声に反応し、泣いたり笑ったりして積極的に接触を求めます 。
興味深いことに、他の霊長類と比較して移動能力の発達が遅れるのとは対照的に、赤ちゃんの笑顔と遊び顔は、他の霊長類よりも発生的にはるかに早く起こります 。
生後3〜4か月後、人間の幼児は見慣れた個人と不慣れな個人を区別することを学びます 。
生後6か月からは、異物恐怖症、つまり見知らぬ人を恐れることが現れます。
これは、潜在的な危険性に対する適応反応を表す行動であると考えられます。
安全な愛着
もし母親または他の愛着のある人が、幼児の合図に積極的に反応し、その結果、幼児が必要なときに母親が利用可能で反応が良いと認識する場合、安全な愛着が最も容易に発達します。 安全な愛着は、子どもへの投資に対する親の意欲と能力を反映しており、したがって、高い親の投資を示しています。 典型的に、安全な愛着のある子どもは、再会の際に母親に近づき、親密さを求めますが、母親の存在下で遊びを再開するかもしれません。 安全な愛着のある子どもは、母親を安全基地または安全な避難所として利用し、そこから好奇心を持って環境を探索できます。 成人期には、安全な愛着は、愛着に関連する経験の一貫した記述によって反映され、これは「自律的な心の状態」と呼ばれます。 安全な愛着は、文化を超えて最も一般的な愛着スタイルであり、おそらく進化の過程で肯定的に選択されてきました。
ただし、親の投資理論(第1章を参照)と一致して、慰めの行動と利用可能性の量に関して、子どもと介護者の間に葛藤がある可能性があります。 ストレンジシチュエーションのシナリオでは、たとえば、一部の幼児が再会の際に母親を避けたり無視したりすることが観察されています。 回避的な愛着のある幼児の愛着のある人は、幼児のニーズに対して拒絶的で冷淡な態度で反応することがわかっています。 彼らは、安全な愛着のある子どもの介護者と比較して、投資が少なく、子どもに自立するように促すようです。 幼児の回避的な反応は、見捨てられるリスクを減らしながら、ある程度の親密さを維持しようとする試みとして解釈できます。 成人として、回避的な愛着のある人は、愛着に関連する経験を理想化し、否定的な経験を軽視することが多く、これは初期の愛着が現在の生活に与える影響を最小限に抑えようとする試みと見なすことができます。 この心の状態は、「無視型」と呼ばれます。
「両価的」または「抵抗型」と呼ばれる別の愛着スタイルは、再会の際の幼児による接触希求と怒りの反応の混合によって特徴付けられます。 両価的な愛着のある子どもの多くの介護者は、その行動に一貫性がありません。 投資をしばしば嫌がる回避的な愛着のある子どもの介護者とは対照的に、両価的な愛着のある子どもの介護者は、感情的な利用可能性の欠如の結果として、投資できないことが多いです。 愛着のある人は、原則として幼児に近づくことができますが、子どもがストレスを感じているときに慰めの行動を提供することができません。 したがって、かんしゃくは、親の努力を最大化するための適応反応である可能性があります。 成人期には、両価的な愛着は、「とらわれ型」の心の状態に反映され、愛着に関連する経験を説明する際の曖昧さとして現れることがあります。
最後に、混乱型または失見当型の愛着スタイルは、再会の際の子供の矛盾した行動のパターンによって特徴付けられます。 回避は、強い接触希求行動に続くか、または同時に表示されることがあります。 「凍りつき」は、混乱した愛着の結果として起こりうる別の形の行動です。 苦痛の表現は、介護者から離れる動きと組み合わせて発生する可能性があります。 成人として、混乱型の愛着のある人は、愛着関係を説明するときに連想の緩みやその他の推論の誤りが明らかになる「未解決」または「混乱した心の状態」を示します。 混乱型の愛着のある子どもの愛着のある人は、しばしば恐ろしいと経験されるか、または自分の幼児に怯えていると感じるかもしれません。 虐待的な行動は、混乱した愛着のある幼児の介護者に最も多く見られ、そのような子どもは、異常に高いレベルの攻撃性、解離症状、うつ病、不安、または仲間や物体への方向転換された攻撃性を含む、さまざまな精神病理を発症する特に脆弱な状態になります。 このような行動は、愛着のある人の利用可能性に対する認識された脅威への反応として説明でき、その結果、恐怖、怒り、悲しみなどの否定的な感情がより広範囲になる可能性があります。
後の3つの愛着スタイルは、安全な愛着とは対照的に、「不安定な愛着」という用語に分類できます。 不安定な愛着のある子どもは、強い愛着行動を示す可能性があり、愛着のある人の感情的な温かさと利用可能性のために苦労する可能性があります。 ただし、愛着行動の強さは、幼児と介護者の間の愛情の絆の強さと混同してはなりません。 不安定な愛着のある子どもは、安全な愛着のある子どもとは異なる、自分の社会的環境がどのように反応するかについての期待を発達させます。 社会的相互作用のこれらの期待の違いは、内的作業モデルの違いと因果的に関連しています。 たとえば、拒絶を予期する子どもは、慰められると確信している子どもと比較して、再会の際に母親に慎重に近づきます。 幼児の問題は、虐待的な介護者にさえ愛着を持たざるを得ない可能性があり、脅威と危険の源である介護者に愛着を持つという両価的な状況に陥る可能性があることです。 このような状況は、激しい恐怖の感情状態を生み出す可能性があり、それがさらに愛着行動を増加させます。 これは、大きな不確実性と予測不可能性の状況を引き起こし、子どもは安定した内的作業モデルを発達させることができない可能性があります。
不安定な内的作業モデルは、不安定な愛着の結果として、個人が他者の思考や意図についてどのように考えるか、つまり「心の理論」と呼ばれるものに大きな影響を与えます(精神分析の文献で使用される別の表現は「反射機能」です。 「社会脳」の詳細については、第2章の考察を参照)。 逆説的な状況は、安全な愛着のある人が他者の精神状態についてより速く考えることを学ぶ可能性があることです。 一方で、安心感と感情的な温かさは、実際に心の理論システムをオフにする可能性があります。 安全な愛着のある人の介護者は、不安定な愛着のある人の拒絶的な介護者よりも「心的」な表現を使用する可能性が高いためです。 一方で、安心感と感情的な温かさは、実際に心の理論システムをオフにする可能性があります。 対照的に、不安定な愛着は、発達的に遅延した、または欠陥のある心の理論システムと結びついている可能性があり、それは否定的な感情に関連する状況でより頻繁に活性化される可能性があります。 この逆説は、強化された猜疑心やパラノイア反応を含む、さまざまな精神病理学的兆候や症状を引き起こす可能性があります(第17章を参照)。 同様に、子どもと介護者の間の言葉によるコミュニケーション
養育者と子供の間の愛着とコミュニケーションの違いは、さまざまな形で相互作用し、愛着スタイルは世代を超えて「受け継がれる」可能性があります。 これらの違いは主に環境条件によって引き起こされますが、重要な遺伝子-環境相互作用の相当な証拠があります。 たとえば、モノアミノオキシダーゼ阻害剤(MAO-AおよびMAO-B)コード遺伝子の特定の多型を保有する人は、MAO-Aの活性低下につながり、人格障害または反社会的行動を発症するリスクが高くなります。 遺伝的変異が異なる社会的行動を説明するのは、それらが発達の初期段階で悪影響にさらされた場合のみであり、これらの遺伝子-環境相互作用は、良好な初期生活条件下では現れません。 言い換えれば、MAO-A活性だけでは行動の個体差を説明できませんが、研究結果は、遺伝的変異が発達の初期段階における社会的経験に対する感受性の違いを説明し、これらの相互作用が人格発達に大きな影響を与えることを強く示唆しています。
生物学的な観点から見ると、青年期および成人期は、個人が同盟を結び、ロマンチックな関係を求める準備をする、主要な生理学的および社会的変化をもたらします。 養育者の感情的な利用可能性は、養育者からの自立を促し、子供が社会的スキルを獲得し、社会的および物理的世界に関する内的作業モデルを洗練することを促進します。 養育者の幼児のニーズへの敏感な対応は、依存を強めるのではなく、後の人生における子供の自立を育むことを強調する必要があります。 この事実はしばしば誤解されています。 したがって、子供時代の経験、特に幼児期の愛着の性質が、仲間との関係や親密な関係、自分の子供との関係、そしておそらくは思春期の発生時期、性的活動、自分の子供への親の投資の量に関連する後の人生の行動に与える影響は、どれだけ強調してもしすぎることはありません。 これは、個人の初期の養育条件が、対人関係の信頼性や安定性を含む、資源の利用可能性と予測可能性に関する個人の期待を導く発達中の内的作業モデルを決定するためです。 ボウルビィは当初、安全な愛着と信頼できる内的作業モデルが、進化によって設計された唯一のものであり、他の形の愛着は規範からの逸脱であると考えていました。 しかし、進化論的な観点からは、環境条件の変動は、おそらく限られていますが、重要な生物社会的目標を達成するためにさまざまな戦略を選択することによって、最終的に生殖適応度を促進するための、環境的偶発性に対する一連の柔軟な適応行動反応を選択した可能性があると考えるのが妥当です。 たとえば、現実のものであれ知覚されたものであれ、厳しい環境条件が幼少期の経験と一致する場合、個人の内的作業モデルが将来の資源の利用可能性の予測不可能性を示唆する可能性があるため、個人は他者の資源の搾取を含む即時の資源抽出、早期の交配および生殖を目指す可能性が高くなります。 対照的に、子供にとって不利な養育環境は、心理的および生物学的な準備を引き起こし、それはr選択的な方向へと移行する可能性があります。
感情的な温かさと豊富な資源は、K選択的な方向性を高める可能性があります。 したがって、青年期および成人期は、小児期および青年期に獲得された内的作業モデルを洗練し、特定の環境条件に適応させる機会を提供します。 したがって、人生の初期の経験は、後の人生の出来事に対する個人の行動と期待に大きな影響を与える可能性があります。
青年期
青年期は、約10〜12歳で始まり、約18〜20歳で終わる期間であり、人間の生活史における別の重要な段階を表しています。 青年期は、霊長類の中で最も長く、青年は繁殖年齢に達します。 青年期は、他の霊長類と比較して、性的成熟の開始の遅れ、つまり思春期の開始の遅れによっても特徴付けられます。 人間の思春期は、他の霊長類と比較して、約4〜5年遅れて始まります。 人間の思春期の延長は、おそらく、認知能力の発達を可能にするために選択されたでしょう。 青年期は、脳の再編成を伴う、人間の生活史の重要な段階です。 この再編成は、社会認知、つまり他者の精神状態を理解し、それに応じて行動する能力にとって重要な脳領域である前頭前野に特に影響を与えます。 青年期は、他の霊長類と比較して、脳の成長と発達が著しく延長されることによって特徴付けられます。 脳の成長は、10代前半にピークに達し、その後、灰白質の体積が減少します。 この灰白質の減少は、おそらく、思春期の間にシナプス刈り込みと呼ばれるプロセスを反映しており、それによって、あまり使用されないシナプス結合が排除され、より効率的な神経処理につながります。 興味深いことに、シナプス刈り込みは、思春期に前頭前野で最も顕著です。
青年期は、子供から大人への移行期間と見なすことができ、その間に個人は、繁殖年齢に達し、仲間との同盟を形成し、ロマンチックな関係を求める準備をします。 この移行は、生物学的、心理的、および社会的な変化を伴います。 生物学的には、青年期は、生殖能力の発達と二次性徴の出現によって特徴付けられます。 心理的には、青年期は、アイデンティティの形成、自律性の発達、および社会的認知の増加によって特徴付けられます。 社会的には、青年期は、家族からの独立性の高まり、仲間との関係の重要性の高まり、および新しい社会的役割の探求によって特徴付けられます。
青年期は、人間の生活史の重要な段階であり、その間に個人は、繁殖年齢に達し、仲間との同盟を形成し、ロマンチックな関係を求める準備をします。 青年期は、生物学的、心理的、および社会的な変化を伴う、子供から大人への移行期間です。 青年期は、脳の再編成を伴う、人間の生活史の重要な段階であり、それは社会認知にとって重要な脳領域である前頭前野に特に影響を与えます。 養育環境は、個体の生活史戦略に大きな影響を与える可能性があります。 たとえば、子供にとって不利な養育条件は、心理的および生物学的な準備を引き起こし、それはr選択的な方向へと移行する可能性があります。 対照的に、感情的な温かさと豊富な資源は、K選択的な方向性を高める可能性があります。
攻撃性、非従順、衝動性の高さによって証明されるように、問題を外在化する可能性が高くなります。 若い男性として、不安定な愛着を持つ個人は、無差別で日和見的な性的関係を持つ可能性が高くなります。 非ランダムな(分類的な)配偶選択のために、うつ病の女性は反社会的な特徴を持つ男性と結婚する可能性が高く、親の投資レベルが低いことが「遺伝する」ようになります。
対照的に、配偶者の調和、低いストレスレベル、および十分な経済的資源を持つ家庭で育つ子供は、介護者のより大きな感情的な温かさと利用可能性に触れる可能性もあります。 これらの子供は、信頼できる内的作業モデルを発達させ、世界を安全な場所として経験する可能性が高くなります。 安全な愛着を持つ人は、本質的に、思春期になるのが遅く、性交を早まる可能性が低いです。 成人として、安全な愛着を持つ人は、長期的な関係を持つことが多く、自分の子供にもっと投資する傾向があります(図3.2)。
図3.2は、資源の利用可能性に応じた、異なる生殖戦略の進化的発達経路を示しています。
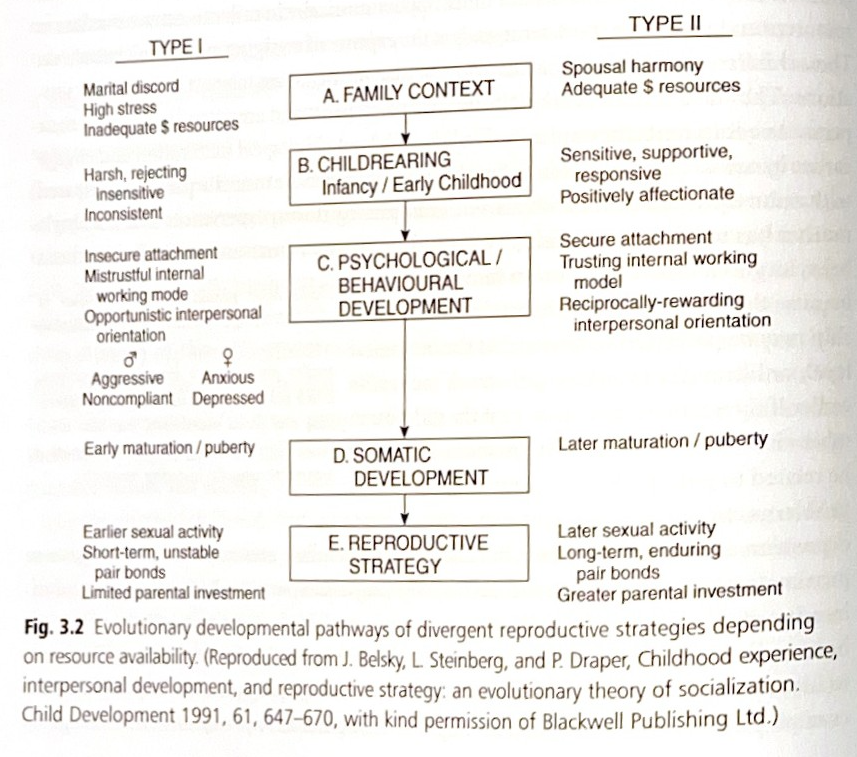
これらの正反対の生活史戦略は、種内のr選択とK選択の間の、ありとあらゆる中間的な変異を含む両極端と見なすことができます。 生物学的に言えば、それらは同じ種の中の「形態」を表しており、その頻度は特定の境界内で個体群レベルで維持されています。 およそ3分の2の子供が、集団全体で安定した愛着を示し、約20%が回避型愛着、約10%が両価型愛着を示します。 無秩序型愛着に関する異文化間のデータはありません。 いずれにせよ、不安定な愛着スタイルは、広範囲の精神病理に対する(非特異的な)危険因子となりますが、個々の遺伝的構成と重大な相互作用をします。 すでに述べたように、養育スタイルは、母親と子供両方の予想される、または実際に利用可能な資源に応じて、世代を超えて「受け継がれる」可能性があります。 将来の環境条件の無意識的な推定は、現代生活の副産物ではありません。むしろ、それは人類の進化の歴史を通じて適用されてきました。 人間の母親間の高いレベルの協力が、出産間隔を数年短縮することを可能にし、それが初期の人類の繁殖上の適応度を高めました。 しかし、他の霊長類や類人猿とは異なり、人間の母親は同時に複数の依存した子供を養育する負担を抱えており、それは男性とのパートナーシップを強化するメカニズムを進化させたからこそ可能になったのです。その中でも、隠れた排卵と強い эмоциональнаяな絆が「核家族」の形成に最も貢献しました。 加えて、人間の女性は自身の生殖可能期間を超えた特定の役割を担うように選択されてきました。これについては次のセクションで扱います。
- 生殖可能期間後の期間
人間の生物学における大きな謎の一つは、女性が閉経後20〜30年生きるという進化的な謎に関連しています。 一見すると、これは生殖上の適応度という観点からは妥当ではないように思えます。 閉経は動物界ではそれ自体珍しいことではありませんが、人間の女性の閉経後の長い寿命は珍しいです。 人間の生殖能力は、初潮から徐々に増加し、30歳前後でピークに達した後、着実に減少し、最終的には停止します。 他の類人猿と同様に、人間の女性は50歳前後で排卵を停止します。これは、流産や遺伝的な先天性欠損症のリスクが年齢とともに増加するため、適応的です。 しかし、他の霊長類とは異なり、人間の女性だけがさらに20〜30年生き、この生活史パターンは現代の生活環境の副産物ではありません。狩猟採集社会の女性は、60歳に達すると、さらに10〜20年生きる可能性が高いのです。 人間の閉経後の長い寿命は、おそらく、すべての段階における全体的な生活史の延長と密接に関連しています。 人間が繁殖を始めるまでには通常約20年かかり、親世代への依存もそれに応じて長いです。 この観点からすると、最後の子孫が自立するまで生き残る可能性が低いため、50歳を超えて繁殖することは意味がありません。進化は、閉経後の女性が孫の生存に貢献したため、女性の長い生殖可能期間後の(閉経後の)寿命を選択しました(「祖母仮説」)。
さらに、選択は近親者への利他主義を好み、閉経後の女性による追加資源の提供は、特に彼女自身の子供たちの生存を助けたと主張されてきました。これは、「祖母仮説」として知られるシナリオです。 実際、狩猟採集社会の閉経後の女性は、自分たちが消費できるよりもはるかに多くの食用資源を部族に提供します。 彼女たちは、生殖年齢の女性よりも多くの時間を食料の収集と加工に費やします。 ほとんどの狩猟採集社会では(人間の祖先の状態のモデルとして)、この余剰は主に近親者の子孫、特に孫に利益をもたらします。 これは進化論的に理にかなっています。なぜなら、祖先の環境では、離乳したばかりの子供は飢餓の危険にさらされており、祖母からの追加のカロリー提供が生存の可能性を高めたからです。 したがって、老化の遅延と長寿の選択は、そもそも、非常に減速した人間のライフサイクルと、すでに離乳したものの依存度の高い子孫の生存の可能性を、追加の高カロリー食を提供することによって高める必要性と関連していた可能性があります。 悲しいことに、私たちは現在、アフリカで何百万人ものHIV孤児の悲劇を目の当たりにしています。子供の両親が後天性免疫不全症候群(AIDS)で亡くなった後、彼らの祖母が依存している孫の養育と世話を提供する上で実際に非常に価値があります。 閉経後の女性は、貴重な資源の場所や季節変動に関する知識、および紛争解決を含む高度な社会的スキルも評価されていたと考えられます。 同様に、高齢の男性も、狩猟スキルや部族社会内での社会問題の交渉という点で、同様の理由で評価されていた可能性があります。 この仮定と一致して、意味記憶の貯蔵に必要な結晶性知能は高齢になっても安定しているのに対し、流動性知能と認知の柔軟性は年齢とともに低下することは言及する価値があります。 しかし、高齢の女性への敬意は、狩猟採集への依存度が低い人間文化でははるかに低かったです。 ごく最近まで、食事に肉の量が多い部族社会では、高齢の女性は食料の提供にあまり貢献できなかったため、安楽死(社会的に認められた男性によって実行される)さえ行っていました。 この負の側面は、おそらく、母親と子供の間の特別な愛情の絆と、人間における利他主義と血縁の大きな重要性によって相殺されたのでしょう。 養育者への愛着は、高齢の親が若い成人、通常は自分の成人した息子または娘との親密さを求める役割逆転さえも引き起こす可能性があります。 しかし、高齢者の価値低下は、狩猟採集社会に特有のものではありません。 現代文化は多世代家族の崩壊につながり、その結果、高齢者の家族生活への関与が欠如していることが、高齢者集団における非常に高い抑うつ率の一因となっているのかもしれません。
追記:出生前環境と生物心理社会的準備
母親の出生前ストレスに関する文献が増えており、後の人生における子孫の心理的問題は、出生前のストレッサーの強度とタイミングに関連している可能性があることを示唆しています。 「胎児プログラミング」の概念は、子宮内環境が出生後の状態に胎児の организмаを準備することを示唆しています。 たとえば、妊娠中の母親のうつ病や不安は、出生時体重の低下や早期出産、および学齢期の子供の行動問題と関連しています。 たとえば、妊娠12〜22週の母親のネガティブな感情が高いレベルにあると、8〜9歳および14〜15歳の子供(男の子は女の子よりも多い)の衝動性および多動性を予測します。 さらに、母親の出生前ストレスは、子供の不安症状、外在化問題、および注意欠陥を予測する可能性があります。 興味深いことに、出生前ストレスは側方性の低下も予測します。これは、自閉症および統合失調症と関連しています。 統合失調症のこの可能な病態生理学的メカニズムは、妊娠の最初の3か月で近親者を亡くしたことによる母親のストレスが、子孫の統合失調症のリスクを大幅に高めるという観察によって、最近裏付けられました。 母親のストレスの多い経験が子孫の行動に深刻な影響を与える可能性があることは明らかですが、ストレッサーの特異性とタイミングの問題についてはほとんど合意がありません。 長期間にわたる慢性的なストレスは、単一のストレスの多い出来事よりも胎児に大きな影響を与えると考えられます。 夫婦の不和または別居は、特に重大であるようです。 母親と胎児の血漿コルチゾールレベルが高度に相関していることが示されており、動物モデルは、出生前ストレスに関連するエピジェネティックな因子(メチル化)が、視床下部-下垂体-副腎皮質軸(HPA軸)を介したストレス誘発コルチゾール反応のフィードバック制御を損なう可能性があるという事実を示しています。 これらの発見と一致して、出生前ストレスは、ヒト以外の霊長類において、海馬の体積の減少と神経新生の減少、HPA機能の変化、および新しい刺激または社会的孤立に対する慢性のストレス反応の亢進を引き起こします。 これらの影響は、霊長類で妊娠初期にストレス曝露が起こると顕著になるようです。 しかし、人間の脳の発達期間の延長と生理学的な早産を考慮すると、人間の胎児と乳児の脳は、かなり長い期間、母親のストレスに対して脆弱であると考えられます。 これらのシナリオは、種内のr選択戦略とK選択戦略の変異モデルの論理的な拡張であるように思われます。 進化的に適応した環境(EEA)にいる人間の母親は、社会的理由または危険が増大した時期に、より苦痛を感じ、不安で警戒心が強かったため、出生前の子孫を予測不可能な環境条件に備えさせ、それによって子孫の行動を即時の資源抽出と日和見的な行動のr選択方向にわずかに押し進めた可能性があります。 もう1つの例は、ある程度推測的かもしれませんが、人間の心理と行動の関係を説明する上での生物学的原理の妥当性を示しているかもしれません。ニューヨーク市でのテロ攻撃後2か月で、女性と比較して男性の胎児死亡率が不均衡に増加しました。これは、第1章で概説したトリバーズ=ウィラード仮説を彷彿とさせる発見です。 女性は(もちろん無意識のうちに)選択的に男児胎児を中絶させたのでしょうか?なぜなら、出生後の生殖上の適応度の見通しが女児の子孫の方が大きかったからでしょうか? これらの仮説的な因果関係は、将来的に前向きに研究する必要があります。
参考文献
Bakermans-Kranenburg, M. J. & van Ijzendoorn, M. H. (印刷中)、「愛着の安定と不安定の行動遺伝学および分子遺伝学:遺伝的脆弱性または差異感受性?」行動脳科学
Belsky, J.、Steinberg, L. & Draper, P. 1991、「幼少期の経験、対人発達、および生殖戦略、社会化の進化理論」、Child Development、vol. 62。 pp. 647-670。
Bowlby, J. 1969、「愛着と喪失」。第1巻。愛着、Basic Books、ニューヨーク。
Bowlby, J. 1973、「愛着と喪失」。第2巻。分離:不安と怒り、Basic Books、ニューヨーク、
Bowlby, J. 1980、「愛着と喪失」。 第3巻。喪失、Basic Books、ニューヨーク。
Bradshaw, J. 1997、「人間の進化。神経心理学的視点」、Psychology Press、ホーヴ。
Carter. A. S.、Garrity Rokous E.、Chazan-Cohen, R.、Little, C. & Briggs-Gowan, M. J. 2001、「母親のうつ病と併存疾患、初期の育児、愛着の安定性、および幼児の社会情動問題とコンピテンシーの予測」、Journal of the American Academy of Child and Adolescent Psychiatry、vol. 40、pp. 18-26。
Cassidy, J. & Shaver. P. R. (編)1999、「愛着ハンドブック。理論、研究、および臨床応用」、Guilford Press。ニューヨーク。
Fonagy, P. 1991、「思考について考える:境界性患者の治療におけるいくつかの臨床的および理論的考察」、International Journal of Psychoanalysis、vol. 72、pp. 639-656。
Harlow, H. F. & Zimmermann, R. P. 1959、「幼児のサルの愛情反応」、Science、vol. 130、pp. 421-432。
Hrdy, S. B. 2000、「Mother Nature」、Vintage、ロンドン。
Insel. T. R. 1997、「社会的愛着の神経生物学的基礎」、American Journal of Psychiatry、vol. 154、pp. 726-735。
Kraemer, G. W. 1997、「アカゲザルの初期の社会的愛着の精神生物学。臨床への応用」、Annals of the New York Academy of Sciences、vol. 807、pp. 401-418。
Noftle, E. E. & Shaver, P. R. 2006、「愛着の次元とビッグファイブ性格特性:関係の質の予測における関連性と比較能力」、Journal of Research in Personality、vol. 40、pp. 179-208。
Pearce, J. W. & Pezzot-Pearce, T. D. 2001、「里親の子供への精神療法アプローチ:愛着理論からのガイダンス」、Child Psychiatry and Human Development、vol. 32、pp. 19-44。
Shaver. P. R. & Mikulincer, M. 2005、「愛着理論と研究:人格への精神力動的アプローチの復活」、Journal of Research in Personality、vol. 39、pp. 22-45。
Simpson, J. A. 1999、「現代の進化論的視点における愛着理論」、「愛着ハンドブック。理論、研究、および臨床応用」、J. Cassidy & P. R. Shaver編、Guilford Press、ニューヨーク。
Talge, N. M.、Neal, C. & Glover, V. 2007、「出生前の母親のストレスと子供の神経発達への長期的影響:どのように、そしてなぜ?」、Journal of Child Psychology and Psychiatry、vol. 48、pp. 245-261。
Van Ijzendoorn, M. H. & Sagi, A. 1999、「愛着の異文化パターン:普遍的および文脈的次元」、「愛着ハンドブック。理論、研究、および臨床応用」、J. Cassidy & P. R. Shaver編、Guilford Press、ニューヨーク。
Voland, E. 1998、「人間の生殖の進化的生態学」、Annual Review of Anthropology、vol. 27、pp. 347-374。
第4章
精神病理の原因
- はじめに
精神病理が存在する理由という問いには、さまざまな相互に補完的な科学的視点からアプローチできます。 伝統的なアプローチは、遺伝的素因(遺伝的リスク)、幼少期の逆境体験(第3章を参照)、個人の生涯のいつでも起こりうる他の心理的外傷となる出来事、明らかな脳損傷、または老化などの障害の近位原因(第1章も参照)に限定されています。 このアプローチに根本的な誤りはありません。 精神病理のさまざまな考えられる原因に関する知識は、主に精神医学遺伝学、脳イメージング技術の進歩、および幼少期の逆境体験と後の人生における精神病理の発達との関係の理解の向上により、過去20年間で大幅に増加しました。 しかし、臨床精神医学と実験神経科学の両方に役立つ現代的な概念的枠組みは、今日まで未解決の追加の問題に取り組む必要があります。 その1つは、精神医学内の専門分野が乖離する傾向があり、相互交流が限られた別々の生活を送るという事実に関連しています。 言い換えれば、一方では精神医学遺伝学、神経科学、および精神障害の動物モデルの分野と、他方では精神療法、社会精神医学、または精神障害の異文化モデルのさまざまな分野との間に、共通の基盤がほとんどないようです。 前者はしばしば「生物学的」と認識され、後者は「心理学的」と認識されます。 ごく最近になって、精神医学内の生物学的および心理学的「学派」間で対話が始まりました。 それにもかかわらず、精神病理学的状態の近位原因の科学的探求だけでは、その説明力は依然として限られています。
精神医学遺伝学、神経科学、および行動臨床的視点は、機能不全の「原因」(メカニズム)、「いつ」機能不全が始まるか(個体発生)、「なぜ」メカニズムの元の機能がそのように設計されているか(適応機能)、およびそれが「どのように」進化したか(系統発生)という問いを統合することができません。 これらの4つの「W」の問いは、もともとニコラス・ティンバーゲンによって提唱されたもので(第1章を参照)、機能と機能不全の近位原因と究極原因の両方を含むメタ理論的枠組みの基礎を形成しています。 これは、いかなる精神病理学的兆候または症状も、それ自体に持つ適応的価値がないと言うことではありません。 定義上、それらは現在の環境および祖先の環境の両方において不適応です。
90
定義上、精神病理学的兆候と症状は、現在の(近位の)視点と究極の視点の両方において不適応です。 なぜなら、それらは個人に害を及ぼし、異常な強度(機能低下、機能亢進、または調節障害)、不適切な状況での出現、および/または異常な持続時間によって機能不全を引き起こすからです 。 しかし、すべての兆候または症状は、進化した心理学的メカニズムのセットの一部として、その機能的な対応物を持っています 。 したがって、機能不全(または精神病理学的兆候と症状と呼ぶことができるもの)は、正常な適応メカニズムの極端な変動として記述でき、その進化した機能を探求することができます 。 しかし、異常と認識されるものは、文化的な規範や価値観から自由な客観的な評価の問題だけではありません 。 それどころか、何が異常と見なされるかは、文化的背景によって大きく異なる場合があります 。 現在の知識は主に高収入国での研究に基づいており、現在の診断マニュアル(精神障害の診断と統計マニュアル、DSM。および国際疾病分類、ICD)はヨーロッパと北米で編集されているという事実にもかかわらず、またはおそらくその事実のために、これを見過ごしてはなりません 。 言い換えれば、DSMおよびICDの分類システムは、西洋以外の文化的背景にとっては狭すぎる可能性があります 。 私たちの文化排他的な視点は、私たちが精神病理を概念化する方法が科学的に正当化された唯一の方法であり、「文化結合症候群」はシステムへの異質な例外として扱われるべきであることを示唆しています 。 しかし、より広い視点では、「アモク」、「ラタ」、「コロ」がそうであるように、注意欠陥/多動性障害(ADHD)、摂食障害、または薬物およびアルコール依存症の流行も同様に「文化結合」である可能性があります 。 診断の慣例は文化内でも異なる場合があります。 特定の状態が障害になる閾値は、固定されているというよりも柔軟です。 これは、精神障害の早期発見と混同してはなりません 。
精神医学的診断はある程度、文化から独立しているわけではありません。 現在の概念化は、西ヨーロッパの小さな地域で生成されています 。 したがって、精神医学的障害の診断は、少なくとも同じくらい価値観や診断の慣例に依存しています。 後者は文化内で盛衰することさえあり、時には精神医学のような分野が診断の閾値を変更することによって独自の市場を作り出すほどになるかもしれません。 すべての悲しみがうつ病に発展するわけではなく、落ち着きのない好奇心旺盛な子供が必ずしもADHDであるとは限らないなどです 。 この分野で働く人々は、精神医学的状態の過剰診断を避けるために、この自己批判を心に留めておくかもしれません 。 図4.1は、変化する診断閾値に関連する問題を示しています。
ただし、この閾値の問題は、できるだけ早く正確な診断を下すための努力と混同してはなりません 。 精神障害の一次予防が現実的な目標であるかどうかは、激しく議論されています 。 いずれにせよ、
(a)
平均的な条件下での「正常」
はじめに 91
(b)
極端な条件下での「正常」
(c)
緩和された診断閾値の下での「正常」
図4.1 (a)変動の極端としての精神病理学的兆候と症状。 適応特性の重大な機能低下または機能亢進のみが病理学的と見なされます 。 (b)極端な環境条件下では、診断閾値がX軸に沿って移動する可能性があり、その結果、一方の端での極端な特性の変動は依然として「正常」と見なされる可能性があります(グラフの右側)。 一方、もう一方の端での変動は、平均的な条件下では「正常」であると考えられる病理学的と見なされる可能性があります(左) 。 たとえば、差し迫った脅威や危険(戦争など)の時代には、パラノイアの瀬戸際にある極端な疑念が「正常」と見なされる場合があります。 一方、信じやすさの境界にある信頼は「異常」と見なされる場合があります 。 同様に、極端な条件下では、高い警戒心と不安が「正常」と見なされる場合があります。 一方、低いレベルの警戒心と不安はすでに正常範囲外です 。 (c)おそらく診断基準の変化、または医療業界が軽度の非特異的な病気の「治療」のための新しい分野を開きたいという願望の結果としての、精神病理学的兆候および症状に対する緩和された診断閾値 。
92
現在の精神医学の概念化では、精神病理学的兆候や症状の現れ方の性の違いを説明することもできません。 たとえば、(近位の)性ホルモンの役割を強調しても、男女間の認知、感情、行動の違いが存在する理由や、それが精神障害の診断体系にどのように反映されるかはまったく説明できません。 たとえば、境界性パーソナリティ障害は女性に非常に多く(または診断される)のに対し、反社会性パーソナリティ障害は男性に多くなっています。 色情症、つまり社会的に地位の高い人に愛されているという妄想は、女性に桁違いに多く見られます。 対照的に、妄想性嫉妬はほぼ男性にのみ発生します。 これらの矛盾に対する説明は、標準的な精神医学的分類学では示されていません。 さらに、精神障害は広く重複しており、統合失調症と双極性感情障害、強迫性障害と妄想性障害など、カテゴリー的に明確な疾患実体は存在しないという証拠が増えています。 同様の関連問題は、併存疾患の診断に関係しています。 DSMおよびICDの無理論的な診断マニュアルでは、無限の数の併存する精神障害の診断が可能ですが、特定の併存疾患は他の疾患よりも頻繁に併発します。 たとえば、再発性うつ病、社交不安障害、および回避性パーソナリティ障害は、1人の個人に妥当に診断できますが、統合失調症または双極性障害の患者に不安障害を診断するのははるかに困難です。
この概論の中心は、精神医学内の専門分野を統合する努力を促進することです。 なぜなら、診断と治療は、年齢、性別、生物学的素因、逆境となる(初期の)経験、および社会経済的背景を考慮に入れて個別に調整されることが患者の利益になるからです。 これは、精神分析と行動療法が驚くほど長く存続しており、(一部の)医学校でまだ教えられているという事実を認識する必要があります。 ただし、エディプスコンプレックスの妥当性や、すべての行動が学習され、したがって学習解除できるという仮説を裏付ける経験的証拠はほとんどありません。 私たちが精神障害の有病率の増加と、人が心理的問題を発症しやすくする個々の状況を理解したい場合、私たちの種の生物学的歴史、特に私たちの種の進化した心理学的メカニズムを考慮に入れた進化論的視点は、精神障害の有病率の世界的な増加を説明するだけでなく、精神障害に対する回復力に関する研究に情報を提供する可能性があります。 世界保健機関(WHO)は、精神病理のリスク因子と保護因子をいくつか特定しました。その多くは、私たちの種の生得的な脆弱性または進化した心理的ニーズに対応する、それぞれ不利な状況または好ましい状況を反映しています(表4.1)。
表4.1 精神障害のリスク因子と保護因子
リスク因子
- 注意欠陥
- 児童虐待とネグレクト
- 攻撃性、暴力、トラウマへの曝露
- コミュニケーション逸脱
- 家族の対立または家族の組織不全
- 神経化学的不均衡
- ストレスの多いライフイベント
- 妊娠中の物質使用
- 慢性不眠症
- 慢性疼痛
- 早期妊娠
- 高齢者虐待
- 感情的な未熟さと制御不能
- 過度の物質使用
- 孤独
- 低出生体重
- 低い社会階級
- 医学的疾患
- 親の精神疾患
- 言葉のスキルと習慣の欠如
- 読字障害
- 親の物質乱用
- 周産期合併症
- 個人的な喪失、死別
- 感覚障害または器質的障害
- 社会的能力の欠如
保護因子
- 思いやり
- 学業的成功
- 社会情動的成長
- 運動
- 安心感
- 習得と制御の感覚
- 肯定的な愛着と早期の絆
- 肯定的な親子相互作用
- 向社会的な行動
- 生活スキル
- ストレスに対処する能力
- 逆境に立ち向かう能力
- 適応性
- 早期の認知刺激
- 良い子育て
- 読み書き能力
- 問題解決スキル
- 自尊心
- 社会的および紛争管理スキル
- 家族や友人の社会的支援
- 自律性
- ストレス管理
出典:World Health Organization Summary Report on the Prevention of Mental Disorders (2004), Box 5, page 23, with kind permission of the World Health Organization
精神病理の究極の原因を統合する上で重要な側面は、私たちの種で進化した心理学的メカニズムが、精神病理学的状態の発症に対する回復力に関する研究をどのように強化できるかという知識です。
94
次のセクションで見るように、精神病理の最も重要な理由の1つは、一部の人々が、過去の世代からの行動生物学的遺産と現代の生活環境との不一致に苦しむ可能性が他の人々よりも高いということです(精神医学遺伝学と、精神障害を引き起こす遺伝子が集団の遺伝子プールに存続するというパラドックスの進化的説明の概要については、第1章を参照) 。
- 心理的適応の進化的制約
臨床医は通常、身体や脳の機能はすべて悪くなる可能性があることを当然のことと考えており、その理由について考えることはありません 。 進化論的な観点からすると、身体の機能不全や精神病理がそもそも存在する理由はそれほど単純ではありません 。 進化過程に関する一般的な理解では、病理は、それが身体的、認知的情緒的、または行動的のいずれであっても、生存と生殖において適応度の不利益をもたらし、したがって何世代にもわたって選択によって排除されるはずであることを示唆しています 。 しかし、この仮定は、少なくとも2つの容易に見過ごされる問題を無視しています 。 第一に、適応の大部分は設計上最適ではありません 。 選択による進化は通常「倹約的な」プロセスであるため、進化した身体的または心理的特徴は、その機能を適切に果たすのに十分に適した設計であるにすぎません 。 さらに、進化は、まったく新しい身体的または精神的特徴を作り出すことはできません 。 新しい適応は、既存の構造から派生します 。 既存の特徴の修正によって新しい機能獲得する進化的プロセスは、時々誤解を招くように「共用」と呼ばれます 。 しかし、それは目標指向(目的論的)または進歩的な発達と間違われてはなりません。 定義上、選択による進化は、目的論的でも進歩的でもありません 。 設計の最適性の問題を理解するために、ヒト科の二足歩行の進化の例を考えてみましょう 。 直立歩行は、おそらくアフリカ南部でのゆっくりとした劇的な気候変動のために、人間の祖先の種で進化したと考えられています 。 約250万年前、熱帯雨林が後退し、サバンナのような環境に取って代わり、木々がより散在するようになったため、二足歩行は木々の間のより大きな距離を移動し、捕食者の存在を周囲を調査するために進化したのです 。 直立歩行には、骨盤と椎骨の解剖学的構造の複雑な再編成が必要でした 。 解剖学的に現代の人類では、椎骨は二重の「S」字カーブを形成しており、これにより、人間は比較的低いエネルギーコストで長距離を移動できます 。 しかし、内臓の垂直方向の整列と、股関節のすぐ上の人体重心の上昇は、椎間板が椎骨間で圧迫され、馬尾または脊髄を圧迫するために滑る可能性があるため、変性問題に対して脆弱性を引き起こす可能性があります 。 したがって、腰椎または頸椎の椎間板ヘルニアは、二足歩行に組み込まれた設計上の妥協点です。 なぜなら、適応問題の解決策は十分ではあったが、最適ではなかったからです 。 しかし、椎間板ヘルニアは確かに適応的ではありませんが、腰痛を発症する避けられない脆弱性因子です 。 同様に、脳機能は設計上の妥協点に満ちています 。 祖先の人類種で脳のサイズと皮質のひだが増加するにつれて、多くのエネルギー関連の問題と、狭い産道を通る問題が発生しました(詳細については、第2章と第3章を参照) 。 これらの問題に対する2つの明白な解決策は、高タンパク質の食事の量を増やし、それとともに腸のサイズを小さくすること、および分娩をより未熟な方向へ前倒しにすることでした 。 さらに、大きな人間の脳は、頭部外傷と乾燥症の影響を受けやすい静脈洞の拡張システムによって維持される、十分に発達した冷却装置を必要としました 。 脳内の多くの設計上の妥協点は、拡張されたニューラルネットワークで情報が伝わる大きな距離、およびすべての新皮質からのトップダウン情報が中脳と脳幹を通過しなければならないという問題に関係しています 。 したがって、(第2章も参照)三位一体の脳の最下層への損傷は、他の2つの機能に深刻な影響を与えます 。 心理的メカニズムの設計上の準最適性に関連する問題は、刺激が反応を引き出すことができる閾値の変動にあります 。 この問題は、特に個人に脅威や危険を知らせる防御メカニズムに関係しています 。 選択による進化は、防御メカニズムに低く、おそらく不安定な刺激閾値を備えさせました 。 たとえば、恐怖と不安は、差し迫った危険または実際の脅威に対する役立つ内部信号です 。 不安反応を引き起こす閾値は通常低いです(個人差があります)。 なぜなら、脅威に反応しない結果として負傷したり死亡したりするコストは、誤報に反応するよりもはるかに高いからです 。 一方で、恒久的に不安になることは役に立ちません。 なぜなら、不安に関連する近位メカニズム(視床下部-下垂体-副腎皮質ストレス軸の活性化によるエピネフリンやコルチゾールの分泌など; HPA)、高血圧、頻脈は、長期的には組織損傷を引き起こす可能性があるからです(これは、心的外傷後ストレス障害の実際の症例です) 。 恐怖反応は、過去に適応的な役割を果たした刺激(ヘビ、クモ、高さ、見知らぬ人、オープンスペース)によってはるかに簡単に引き起こされますが、自動車、放射能、肥満などの現代の脅威によって引き起こされることははるかに少ないです 。 恐怖反応を引き起こすメカニズムは、少量の煙を検出するのに十分な感度を持つ低い閾値で動作する「煙探知器の原理」に例えられています 。 ただし、煙探知器とは対照的に、不安反応の閾値は個人内で変動する可能性があります 。 たとえば、暗闇の中を一人で歩いている人は、信頼できる人々と一緒にいる場合よりも、恐怖反応の閾値がほぼ確実に低くなります 。 したがって、個人の安全と安心感は、環境状況に大きく依存しており、他者との親密さの経験が最も重要であると考えられます 。 第二に、現代の環境は、祖先の生活条件とはほとんど共通点がありません 。 たとえば、私たちの進化の歴史のほとんどの間、おそらく数十万年の間、祖先の人類は、誰もが個人的に他の誰を知っていた緊密なコミュニティに住んでいました 。 祖先の人類社会内の最小機能単位は、父、母、子供からなる核家族でした。
96
精巣のサイズと性的二形性は、穏やかな一夫多妻制が祖先の人類には一般的であったことを示唆していますが、重要なことに、男性が十分な資源を提供するという点で、複数の配偶者(彼が父親である子孫を含む)を「余裕がある」かどうかによって決まりました。 個々の家族は拡大家族に囲まれており、コミュニティは相互扶助と協力に大きく依存していました。 匿名性は事実上存在せず、何千人もの見知らぬ人との日常的な対立は、過去には適応上の問題ではありませんでした。 したがって、現代の人間は祖先の状態に似た社会状況を作り出そうと努めていますが(たとえば、個人の知人の平均数は、新皮質比から予測されたものと驚くほど類似している約150人と推定されています。 第1章と第2章を参照)、資源、社会的地位、および親密なパートナーとの協力、他者との協力、および受け入れられる社会的地位の達成のための競争は、私たちの進化の過去よりもおそらくはるかに高くなっています。 したがって、社会競争が激化した現代の環境条件が、個人のケアを求める、世話をする、仲間を引き付ける、他者と協力する、受け入れられる社会的地位を達成するなどの生物社会的目的を阻止するリスクを高める可能性は、私たちの種の心理的構成が進化した進化適応環境(EEA)と比較して、大幅に増加しています。 過去に進化し、現在の環境ストレス要因となった心理的メカニズムの不一致は、おそらく精神病理の最も重要な進化的原因の1つです。 不一致に対する暫定的な説明は、人間の文化的進化が生物学的進化に追いつき、その結果、生物学が文化的変化に追いつくことができなかったということです。 不一致の問題は、心理的メカニズムにのみ影響を与えるわけではありません。 高血圧、脳卒中、心筋梗塞、II型糖尿病のリスクを含む、多くの「文明病」は、不一致のシナリオに沿って解釈できます。 環境(最も重要な)社会的ストレス要因によるHPA軸の慢性的な活性化が、心血管系に影響を与える障害の主な原因因子である可能性があります。 さらに、「倹約遺伝子」は、カロリーが豊富な食事が豊富な時代に、これらの障害の原因因子となる可能性があります。 「倹約遺伝子仮説」は、最大カロリー抽出のための遺伝子が選択されたと仮定しており、現在、過剰供給と高カロリー食が豊富な時代に害を引き起こしています。 たとえば、倹約遺伝子は、甘いものやコレステロールが豊富な食品の好みを説明する可能性があり、最近まで環境制限下で生活していた北米および南米の先住民集団は、これらの食品を無制限に利用できるようになったときに、肥満、高血圧、およびII型糖尿病を発症する可能性が特に高くなります。
94
次のセクションで見るように、精神病理の最も重要な理由の1つは、一部の人々が、過去の世代からの行動生物学的遺産と現代の生活環境との不一致に苦しむ可能性が他の人々よりも高いということです(精神医学遺伝学と、精神障害を引き起こす遺伝子が集団の遺伝子プールに存続するというパラドックスの進化的説明の概要については、第1章を参照)。
- 心理的適応の進化的制約
臨床医は通常、身体や脳の機能はすべて悪くなる可能性があることを当然のことと考えており、その理由について考えることはありません。 進化論的な観点からすると、身体の機能不全や精神病理がそもそも存在する理由はそれほど単純ではありません。 進化過程に関する一般的な理解では、病理は、それが身体的、認知的情緒的、または行動的のいずれであっても、生存と生殖において適応度の不利益をもたらし、したがって何世代にもわたって選択によって排除されるはずであることを示唆しています。 しかし、この仮定は、少なくとも2つの容易に見過ごされる問題を無視しています。 第一に、適応の大部分は設計上最適ではありません。 選択による進化は通常「倹約的な」プロセスであるため、進化した身体的または心理的特徴は、その機能を適切に果たすのに十分に適した設計であるにすぎません。 さらに、進化は、まったく新しい身体的または精神的特徴を作り出すことはできません。 新しい適応は、既存の構造から派生します。 既存の特徴の修正によって新しい機能獲得する進化的プロセスは、時々誤解を招くように「共用」と呼ばれます。 しかし、それは目標指向(目的論的)または進歩的な発達と間違われてはなりません。 定義上、選択による進化は、目的論的でも進歩的でもありません。 設計の最適性の問題を理解するために、ヒト科の二足歩行の進化の例を考えてみましょう。 直立歩行は、おそらくアフリカ南部でのゆっくりとした劇的な気候変動のために、人間の祖先の種で進化したと考えられています。 約250万年前、熱帯雨林が後退し、サバンナのような環境に取って代わり、木々がより散在するようになったため、二足歩行は木々の間のより大きな距離を移動し、捕食者の存在を周囲を調査するために進化したのです。 直立歩行には、骨盤と椎骨の解剖学的構造の複雑な再編成が必要でした。 解剖学的に現代の人類では、椎骨は二重の「S」字カーブを形成しており、これにより、人間は比較的低いエネルギーコストで長距離を移動できます。 しかし、内臓の垂直方向の整列と、股関節のすぐ上の人体重心の上昇は、椎間板が椎骨間で圧迫され、馬尾または脊髄を圧迫するために滑る可能性があるため、変性問題に対して脆弱性を引き起こす可能性があります。 したがって、腰椎または頸椎の椎間板ヘルニアは、二足歩行に組み込まれた設計上の妥協点です。 なぜなら、適応問題の解決策は十分ではあったが、最適ではなかったからです。 しかし、椎間板ヘルニアは確かに適応的ではありませんが、腰痛を発症する避けられない脆弱性因子です。 同様に、脳機能は設計上の妥協点に満ちています。 祖先の人類種で脳のサイズと皮質のひだが増加するにつれて、多くのエネルギー関連の問題と、狭い産道を通る問題が発生しました(詳細については、第2章と第3章を参照)。 これらの問題に対する2つの明白な解決策は、高タンパク質の食事の量を増やし、それとともに腸のサイズを小さくすること、および分娩をより未熟な方向へ前倒しにすることでした。 さらに、大きな人間の脳は、頭部外傷と乾燥症の影響を受けやすい静脈洞の拡張システムによって維持される、十分に発達した冷却装置を必要としました。 脳内の多くの設計上の妥協点は、拡張されたニューラルネットワークで情報が伝わる大きな距離、およびすべての新皮質からのトップダウン情報が中脳と脳幹を通過しなければならないという問題に関係しています。 したがって、(第2章も参照)三位一体の脳の最下層への損傷は、他の2つの機能に深刻な影響を与えます。 心理的メカニズムの設計上の準最適性に関連する問題は、刺激が反応を引き出すことができる閾値の変動にあります。 この問題は、特に個人に脅威や危険を知らせる防御メカニズムに関係しています。 選択による進化は、防御メカニズムに低く、おそらく不安定な刺激閾値を備えさせました。 たとえば、恐怖と不安は、差し迫った危険または実際の脅威に対する役立つ内部信号です。 不安反応を引き起こす閾値は通常低いです(個人差があります)。 なぜなら、脅威に反応しない結果として負傷したり死亡したりするコストは、誤報に反応するよりもはるかに高いからです。 一方で、恒久的に不安になることは役に立ちません。 なぜなら、不安に関連する近位メカニズム(視床下部-下垂体-副腎皮質ストレス軸の活性化によるエピネフリンやコルチゾールの分泌など; HPA)、高血圧、頻脈は、長期的には組織損傷を引き起こす可能性があるからです(これは、心的外傷後ストレス障害の実際の症例です)。 恐怖反応は、過去に適応的な役割を果たした刺激(ヘビ、クモ、高さ、見知らぬ人、オープンスペース)によってはるかに簡単に引き起こされますが、自動車、放射能、肥満などの現代の脅威によって引き起こされることははるかに少ないです。 恐怖反応を引き起こすメカニズムは、少量の煙を検出するのに十分な感度を持つ低い閾値で動作する「煙探知器の原理」に例えられています。 ただし、煙探知器とは対照的に、不安反応の閾値は個人内で変動する可能性があります。 たとえば、暗闇の中を一人で歩いている人は、信頼できる人々と一緒にいる場合よりも、恐怖反応の閾値がほぼ確実に低くなります。 したがって、個人の安全と安心感は、環境状況に大きく依存しており、他者との親密さの経験が最も重要であると考えられます。 第二に、現代の環境は、祖先の生活条件とはほとんど共通点がありません。 たとえば、私たちの進化の歴史のほとんどの間、おそらく数十万年の間、祖先の人類は、誰もが個人的に他の誰を知っていた緊密なコミュニティに住んでいました。 祖先の人類社会内の最小機能単位は、父、母、子供からなる核家族でした。 精巣のサイズと性的二形性は、穏やかな一夫多妻制が祖先の人類には一般的であったことを示唆していますが、重要なことに、男性が十分な資源を提供するという点で、複数の配偶者(彼が父親である子孫を含む)を「余裕がある」かどうかによって決まりました。 個々の家族は拡大家族に囲まれており、コミュニティは相互扶助と協力に大きく依存していました。 匿名性は事実上存在せず、何千人もの見知らぬ人との日常的な対立は、過去には適応上の問題ではありませんでした。 したがって、現代の人間は祖先の状態に似た社会状況を作り出そうと努めていますが(たとえば、個人の知人の平均数は、新皮質比から予測されたものと驚くほど類似している約150人と推定されています。 第1章と第2章を参照)、資源、社会的地位、および親密なパートナーとの協力、他者との協力、および受け入れられる社会的地位の達成のための競争は、私たちの進化の過去よりもおそらくはるかに高くなっています。 したがって、社会競争が激化した現代の環境条件が、個人のケアを求める、世話をする、仲間を引き付ける、他者と協力する、受け入れられる社会的地位を達成するなどの生物社会的目的を阻止するリスクを高める可能性は、私たちの種の心理的構成が進化した進化適応環境(EEA)と比較して、大幅に増加しています。 過去に進化し、現在の環境ストレス要因となった心理的メカニズムの不一致は、おそらく精神病理の最も重要な進化的原因の1つです。 不一致に対する暫定的な説明は、人間の文化的進化が生物学的進化に追いつき、その結果、生物学が文化的変化に追いつくことができなかったということです。 不一致の問題は、心理的メカニズムにのみ影響を与えるわけではありません。 高血圧、脳卒中、心筋梗塞、II型糖尿病のリスクを含む、多くの「文明病」は、不一致のシナリオに沿って解釈できます。 環境(最も重要な)社会的ストレス要因によるHPA軸の慢性的な活性化が、心血管系に影響を与える障害の主な原因因子である可能性があります。 さらに、「倹約遺伝子」は、カロリーが豊富な食事が豊富な時代に、これらの障害の原因因子となる可能性があります。 「倹約遺伝子仮説」は、最大カロリー抽出のための遺伝子が選択されたと仮定しており、現在、過剰供給と高カロリー食が豊富な時代に害を引き起こしています。 たとえば、倹約遺伝子は、甘いものやコレステロールが豊富な食品の好みを説明する可能性があり、最近まで環境制限下で生活していた北米および南米の先住民集団は、これらの食品を無制限に利用できるようになったときに、肥満、高血圧、およびII型糖尿病を発症する可能性が特に高くなります。
参考文献
Bakerman-Kramelburg, M. J. & van Ijzenboorn, M. H. (印刷中). 「愛着の安全性と不安定性の行動的および分子遺伝学: 遺伝的脆弱性または差異的感受性」
Behavioral Brain Science.
Bekky, J., Steinberg, J. & Draper, P. 1991, 「幼少期の経験: 対人的発達と生殖戦略: 社会化の進化論」, Child Development, vol. 62, pp. 647–670.
Bowlby, J. 1969, 愛着と喪失. Vol. 1, 愛着, Basic Books, New York.
Bowlby, J. 1973, 愛着と喪失. Vol. 2, 分離: 不安と怒り, Basic Books, New York.
Bowlby, J. 1980, 愛着と喪失. Vol. 3, 喪失, Basic Books, New York.
Bradshaw, J. 1997, 人間の進化: 神経心理学的視点, Psychology Press, Howe.
Carter, A. S., Garrity-Rokous, E., Chazan-Cohen, R., Little, C. & Briggs-Gowan, M. J. 2001, 「母親のうつ病と併存症: 早期の育児、愛着の安全性、幼児の社会情緒的問題と能力の予測」, Journal of the American Academy of Child and Adolescent Psychiatry, vol. 40, pp. 18–26.
Cassidy, J. & Shaver, P. R. (編) 1999, 愛着のハンドブック: 理論、研究、臨床応用, Guilford Press, New York.
Fonagy, P. 1991, 「思考についての思考: 境界例患者の治療におけるいくつかの臨床的および理論的考察」, International Journal of Psychoanalysis, vol. 72, pp. 639–656.
Harlow, H. F. & Zimmermann, R. P. 1959, 「幼いサルにおける愛情の反応」, Science, vol. 130, pp. 421–432.
Hrdy, S. B. 2000, 母なる自然, Vintage, London.
Insel, T. R. 1997, 「社会的愛着の神経生物学的基盤」, American Journal of Psychiatry, vol. 154, pp. 726–735.
Kraemer, G. W. 1997, 「アカゲザルの初期社会的愛着の心理生物学: 臨床的意義」, Annals of the New York Academy of Sciences, vol. 807, pp. 401–418.
Noftle, E. E. & Shaver, P. R. 2006, 「愛着の次元とビッグファイブ人格特性: 関連性と関係の質を予測する能力の比較」, Journal of Research in Personality, vol. 40, pp. 179–208.
Perez, J. W. & Perez-Perez, T. D. 2001, 「里子の子供たちへの心理療法的アプローチ: 愛着理論からの指針」, Child Psychiatry and Human Development, vol. 32, pp. 19–44.
Shaver, P. R. & Mikulincer, M. 2005, 「愛着理論と研究: 人格への心理力学的アプローチの復活」, Journal of Research in Personality, vol. 39, pp. 22–45.
Simpson, J. A. 1999, 「現代の進化的視点における愛着理論」, 愛着のハンドブック: 理論、研究、臨床応用, J. Cassidy & P. R. Shaver (編), Guilford Press, New York.
Talge, N. M., Neal, C. & Glover, V. 2007, 「出生前の母親のストレスと子供の神経発達への長期的影響: どのように、そしてなぜ?」, Journal of Child Psychology and Psychiatry, vol. 48, pp. 245–261.
Van Ijzenboorn, M. H. & Sagi, A. 1999, 「愛着の文化的パターン: 普遍的および文脈的次元」, 愛着のハンドブック: 理論、研究、臨床応用, J. Cassidy & P. R. Shaver (編), Guilford Press, New York.
Voland, E. 1998, 「人間の生殖の進化的生態学」, Annual Review of Anthropology, vol. 27, pp. 347–374.
