
要約
基本的な発達過程
- 思春期と成人期には生理的・社会的な大きな変化が起こり、社会的絆や恋愛関係の形成準備がなされる
- 人間は長い子ども時代を通じて社会的スキルを獲得し、内的作業モデルを洗練させる
- 養育者が乳幼児のニーズに敏感に応えることは、依存性ではなく後の自立を促進する
内的作業モデルとライフヒストリー戦略
- 初期の養育経験が内的作業モデルを形成し、対人関係や資源の利用可能性に対する期待を導く
- ライフヒストリー戦略には種内変異があり、環境条件によって調整される
- 厳しい環境条件では、r選択(量重視)の方向に傾く傾向がある
- 温かく安定した環境では、K選択(質重視)の方向に傾く傾向がある
不安定な愛着の影響
- 高ストレスや拒絶的な家庭環境は、不信感の強い内的作業モデルを発達させる
- 不安定な愛着を持つ子どもは忍耐力が低く、共感性が乏しい傾向がある
- 思春期には生物学的成熟が早まり、性行動を早期に開始する傾向がある
- 成人になると短期的な関係を多く持ち、自分の子どもへの投資が少なくなりやすい
安定した愛着の影響
- 夫婦の調和や適切な資源のある家庭では、信頼できる内的作業モデルが発達する
- 安定した愛着を持つ子どもは、相互性と他者への信頼を重視する
- 思春期の到達が遅く、性行動の開始も遅い傾向がある
- 成人になると長期的な関係を持ち、自分の子どもにより多く投資する傾向がある
愛着の種類と分布
- 人口全体で約2/3が安定型、20%が回避型、10%が両価型の愛着を示す
- 不安定な愛着スタイルは精神病理のリスク要因となる
生殖後期間の意義
- 女性の長い閉経後期間は進化的に選択されてきた特性である
- 「祖母仮説」:閉経後の女性は離乳した孫の生存を助けるため進化した
- 高齢者が社会から価値を下げられることは、高齢者のうつ病率の高さと関連する可能性がある
出生前環境の影響
- 母親の出生前ストレスは子どもの行動問題と関連している
- 「胎児プログラミング」:子宮内環境が胎児を出生後の環境に備えさせる
- 妊娠中のストレスは脳の発達に影響を与え、行動傾向をr選択の方向に押し進める可能性がある
第3章
人間のライフヒストリー
1. はじめに
生涯史理論は、個体の一生の中で変化する生存と繁殖に関わる問題に対する、種に特有の解決策を扱います。生涯の構成とタイミングには種によってかなりの違いがあります。K選択とr選択は、生存と繁殖の問題に対する正反対の解決策を反映する対照的な戦略です。
K選択された種は、成長が遅く、繰り返し繁殖し(反復繁殖)、一度の出産で産まれる子どもの数は少なく、長期間の集中的な親の投資(長い妊娠期間を含む)があり、乳幼児期の死亡率は低くなっています。繁殖活動の開始は遅れ、K選択種は長寿を享受します。一般的に、K選択種は大きな体に成長し、寿命が長いため、K選択種の成体間の競争は激しくなります。なぜなら、競争が生存と繁殖の成功を決定するからです。K選択種は通常、環境変動が少ない条件下で生活しています。
反対に、r選択は逆のパターンを持ちます。r選択種は速く成長し、生涯で一度だけ繁殖することが多く(一回繁殖)、子孫の数は多い(または膨大)ですが、成体になるまで生き残るのはほんの一部です。r選択種では出生(または孵化)後の親の投資は少ないか全くなく、性的成熟が早く訪れ、個体は若くして死にます。r選択種は通常、小さな体に成長し、環境変動が激しい中で生活しています。K選択種の子孫は出生時に比較的成熟しており(早成性)、一方r選択種の子孫はより未熟であることが多いです(晩成性)。
哺乳類の中では、多くのげっ歯類がr選択の生涯史パターンを示す一方、霊長類は高度にK選択された種です。特に類人猿とヒトは極端なK選択の生涯史を持つことが際立っており、この発達パターンは種の心理的な特性に大きな影響を与え、個体の一生を通じて変化する要求をもたらします。興味深いことに、ヒトの極端なK選択は、幼形成熟(パエドモーフォシス)とそれに続く過形成(ハイパーモーフォシス)という異時的過程と密接に関連しています。過形成は、大きな脳を含むK選択の特性をヒト(ホモ・サピエンス)が獲得した最も重要なメカニズムかもしれません。人間の生涯史の最も顕著な特徴は、典型的な霊長類のパターンがすべての発達段階で拡張されていることです。
しかし、この章の後半で見るように、生涯史パターンには種内変異があり、一部の人間の生涯史はr選択に近いパターンを示し、他の人はより極端なK選択パターンを示します。r選択に近いパターンが精神病理の発達を促進するかどうかは現在のところ不明ですが、多くの実証的な発見はこの仮定を支持するものと解釈できます。
2. 乳幼児期と児童期
生物学的に言えば、すべての新生児にとって最も重要な課題は、乳幼児期と児童期を生き抜き、性的成熟に達することです。人間の赤ちゃんもこの点では例外ではありません。しかし、人間の赤ちゃんは出生時に非常に未熟であるという問題に直面しています。
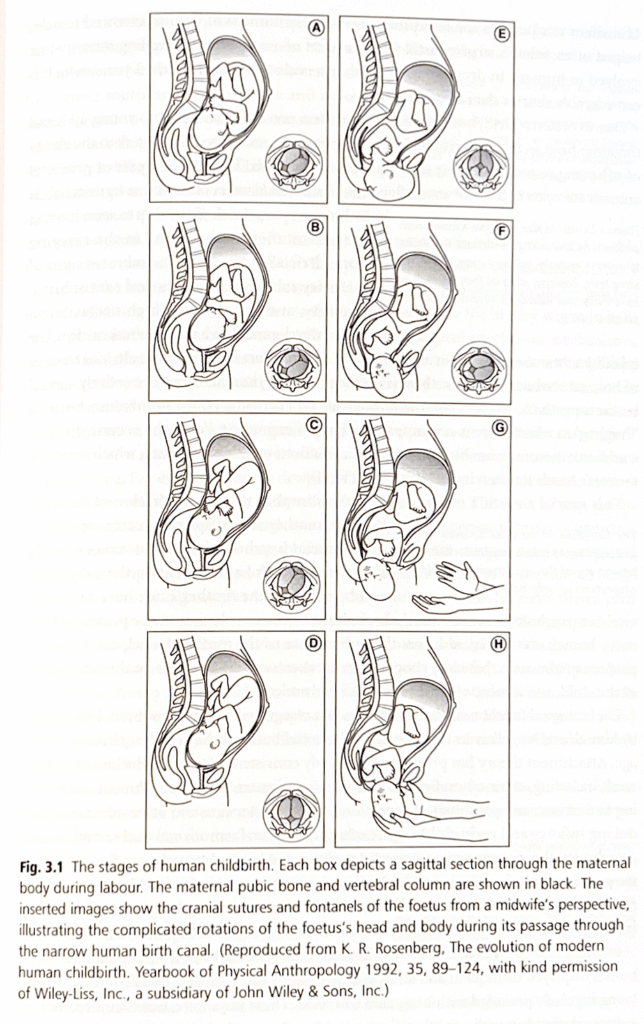
例えば、他の類人猿と比較すると、人間の新生児は生理学的には約13ヶ月早く生まれてきたと言えます。言い換えれば、人間の赤ちゃんがチンパンジーの赤ちゃんと同じ成熟度で生まれるとすれば、人間の妊娠期間は約22ヶ月続くことになります。人間の早期出産は進化的な設計上の妥協を表しており、これは最終的には二足歩行の進化と関連しています。直立歩行に伴い人間の骨盤の解剖学的構造が変化し、その結果、産道が狭くなりました。
これは、初期のヒト科の種が比較的小さなチンパンジーのような約350cm³の脳容量(したがって小さな頭蓋骨)を持っている限りは問題ではありませんでした。しかし、進化の過程で脳(と体)が大きくなるにつれ、狭い産道の問題はより深刻になりました。
人類の祖先種における脳の拡大は、おそらく不連続に起こりました。脳の最初の成長スパートは約180万年前に始まり、おそらくタンパク質の摂取量が増える食生活の変化によって促進され、ホモ・ハビリスの脳容量は350cm³から800cm³へと倍増しました。祖先のヒト科の種はそれよりずっと前から二足歩行をしていたことに注意してください。つまり、二足歩行と脳の拡大は以前考えられていたほど密接に関連していないのです。
脳容量の2回目の「飛躍」は約40万年前に原始的なホモ・サピエンスが進化した時に起こり、平均脳容量は1,300cm³になりました。この2回目の脳容量の増加は、おそらく社会構造の変化と関連しており、人間の社会的脳の発達につながりました(第2章の後記を参照)。
これらの連続的な脳容量の増加により、出産が早まる選択が行われましたが、その代償として出生時の子孫はより未熟になりました。それにもかかわらず、この進化的な設計上の妥協にも関わらず、人間の出産過程は母親と赤ちゃんの両方にとって非常に危険な営みとなりました。
産道を通過するために、人間の赤ちゃんは頭、体、四肢で2回のねじれ運動を行わなければならず、そのため出産は赤ちゃんと母親の両方を様々な合併症で死亡する大きなリスクにさらします。
助産は、おそらく初期の人類における最初の職業であり、経験豊かな女性が他の女性の出産を手伝いました。これは人間で進化した共同育児戦略の一部であり、出産間隔を約3年に短縮しました。これは他の類人猿よりもかなり短い期間です。
人間の乳児の特徴と愛着行動について
人間の赤ちゃんは早期出産のため、若い類人猿と比べると早成性(自立能力)が低く、二次的に一部の晩成性の特徴を保持しています。早成性とは、出生後すぐに自分の足で移動する能力を指します。早成性動物の目や耳は出生時またはその直後に開きます。
人間の新生児の早成性の特徴には、出生時に目が開いていること、すぐに母親と交流できること、抱いている人にしがみつく能力などがあります。一方、晩成性は、出生時に目や耳が閉じていること、動けないこと、裸であることなどの未熟さの兆候を含み、巣や穴での発達期間を必要とします。自力で移動できないことや積極的に快適さと保護を求められないことは、人間の新生児の晩成性の特徴です。
毛皮にしがみつくことができないため、人間の赤ちゃんは母親に積極的に抱かれています。ドイツ語では、人間の新生児に対して「Tragling」(「抱かれる者」)という生物学的用語が導入されましたが、英語には同様の表現はありません。これが可能になったのは、もちろん二足歩行と関連しており、母親の手が子供(や食物)を運ぶために自由になりました。
人間の新生児のこの特殊な状況は、母親と子供の二者関係の形成が初期の人類における最も重要な心理的適応の一つとなったことを意味します。赤ちゃん側の成功した愛着は、人類の進化を通じて、そして現在に至るまでのあらゆる文化において心理的な健康に不可欠です。さらに、母親のヘルパーへの依存が比例して増加し、女性同士の緊密な協力、子どもの母親と父親の間の協力、親族の同盟関係が積極的に選択されるようになりました。
愛着の生物学的機能は、ジョン・ボウルビーによって初めて認識され、彼と彼の後継者によって乳児期から老年期までの普遍的な人間行動の理論へと発展しました。愛着理論は、非ヒト霊長類の実証研究、人間の正常な愛着と絆、そして里親養育や乳幼児期の身体的、感情的、性的虐待などの逆境を含む病理学的状態と完全に一致していることが証明されています。
愛着理論は、安全な愛着と不安定な愛着の心理的、生理学的、行動的結果を予測するための強力なツールです。これらの予測は実証的なテストに開かれており、愛着研究に関連する複雑な遺伝子-環境相互作用の解明は始まったばかりです。
当初、ボウルビーは、里親家庭で育った多くの少年たちが、基本的なケアと食事を定期的に提供されているにもかかわらず、非行や反社会的行動を示し、感情的に不適切であるという事実に衝撃を受けました。しかし、これらの少年たちは母親的存在との緊密な関係を奪われており、ボウルビーはこれが彼らの行動問題の核心にある可能性があると結論づけました。
非ヒト霊長類の研究は後に、若いサルが生物学的母親または代理母との親密さの重要性を確認しました。早い段階で母親から離され、社会的孤立で育てられた新生児のサルは、ミルクを提供するメッシュワイヤーのダミー母親よりも、毛皮で覆われたダミー母親にしがみつくことを好みます。孤立の期間によっては、若いサルは母親や仲間との再会でさえ改善しない重度のうつ病を発症します。さらに、成体になるとこれらのサルは社会的能力に欠け、自分の子孫を育てることができません。
これらの—多くの点で容赦ない—実験は、生涯の心理的健康と社会的能力のために、人生の早い段階での母親的存在との社会的な親密さの重要性を明確に確認しました。生涯史と進化した心理的愛着メカニズム(相同の前頭辺縁脳構造で表される)の大きな類似性を考えると、健全な人間の発達は、おそらく出生前からでさえ、非常に早い段階からの近接性と感情的温かさに依存していることは明らかです。
若い霊長類(人間を含む)は、実際、近接性を求めるよう生物学的にプログラムされています。愛着行動は、乳児の生存可能性を高め、社会的学習を促進するために選択されました。例えば、人間の新生児の体温調節システムはまだ未熟であり、脱水症状にもなりやすいです。これは部分的には、毛皮による断熱材の欠如(ある程度は皮下の褐色脂肪層で補償されていますが)と、全エネルギー摂取量のほぼ3分の2が発達中の脳によって消費されるという事実によるものです。注目すべきは、人間の出産の生理的早期化により、脳は出生後も出生前と同じペースで1年以上成長し続けるということです。
脳のエネルギー消費と発達パターン
その後になってようやく、脳のエネルギー消費は体のエネルギー需要に対して徐々に減少していきます。この成長パターンは、出生後すぐに脳が体の成長に比べてより遅く成長し始める他の霊長類とは異なります(図2.3を比較)。しかし、すでに指摘したように、愛着形成は主に食事を与えられる必要性や欲求とは関連していません。母親と赤ちゃんの間の強い愛情の絆が栄養失調から最も保護してくれるかもしれませんが、幼い乳児の主な目標は近接性と安全を求めることです。
乳児の愛着行動の発達
人間の乳児はこれを達成するためのいくつかの手段を持っています。生後4週間の時点で、人間の乳児はすでに人間の顔の輪郭に強い好みを示します。彼らは人間の声、特に母親の声に反応し、泣いたり笑ったりして積極的に接触を求めます。興味深いことに、他の霊長類と比較して運動発達の遅れとは対照的に、赤ちゃんの笑顔と遊び顔は発生的に他の霊長類よりもはるかに早く現れます。
生後3〜4ヶ月後、人間の乳児は見慣れた人と見慣れない人を区別することを学びます。外国人恐怖症(見知らぬ人への恐怖)は生後6ヶ月から現れ始めます。この行動は、おそらく他の(特に男性の)人間の潜在的な危険性に対する適応反応を表しています。生後6〜9ヶ月の年齢で、乳児は限られた範囲内でハイハイすることによって、愛着対象との近接性をコントロールできるようになります。
しかし、親の養育は行動的手段によってのみ引き出されるのではありません。さらに、人間の乳児は、すべての哺乳類の幼獣や孵化したばかりの鳥のように、大きな目、丸い顔と頭蓋骨、短い四肢などの特徴を示します。これらは通常、攻撃性を減少させ、「かわいい」と認識されます。これらの赤ちゃんらしい特徴に対する反応は生物学的に深く根付いていますが、母親の子供のシグナルに対する反応性は、乳児側の近接性追求行動よりもずっとプログラムされていません。
「愛着」と「ボンディング」の違い
そのため、「愛着」とは対照的に、「ボンディング」という用語は、安定した母子の二者関係の発達を支援する母親の行動を表すために選ばれました。母親は通常、赤ちゃんのニーズに高い反応性を持ち、授乳は母乳育児によって刺激されるオキシトシン分泌を含む心理的および生理学的プロセスを通じて、母親と子供の間の愛情の絆を強化します。しかし、パートナーや親族からの支援的資源を含む資源の利用可能性は、母親が赤ちゃんを受け入れ、絆を結ぶ能力に大きな影響を与えます。
出生後の最初の約24時間は、ボンディングが発達するかどうかに関わらず、人類の進化を通じて重要であり、新生児が母親による新生児殺しや遺棄によって死亡するリスクは、現代社会でさえもこの重要な期間内が最も大きくなります。対照的に、乳児殺し(新生児期後)のリスクは、継父(子供と生物学的に関係のない)と一緒に育つ子供たちで最大です。ここでは、持続的な泣き声と(継)父と子の間の愛情の絆の欠如が悪循環を生み出し、暴力的な行動と子供の被害につながる可能性があります。
母親によって行われる乳児殺しは、はるかに頻繁に母親の精神病理、特にうつ病と関連しています。重度のうつ病を持つ母親は、時には殺人-自殺未遂として、不利な条件下で成長することから乳児を「救出」しようとさえすることがあります。これは新生児殺しや新生児の遺棄とは根本的に異なります。後者は厳密な意味では母親の精神病理と関連することはほとんどなく、しばしば絶望的な独身のティーンエイジャーの母親によって行われます(詳細については、第16章を参照)。
母子の二者関係と愛着の絆
母親と子供の二者関係と彼らの間の愛情の絆は、一度確立されると、持続的で実質的に不可逆的なものです。愛着は特定の個人に特別に向けられており、他の人と交換できません。愛着は複数の人、例えば母親と父親の両方に形成されることもありますが、通常は階層的な方法で形成されます。父親の投資(および感情的関与)は、おそらく他の哺乳類と比較して人間の父親で最も高く、一人の愛着対象がいくつかの領域で重要である一方、別の愛着対象が他の領域で重要になることがあります。興味深いことに、うまく機能している個人は、多くの場合、母親と父親の両方と安全な関係を持っています。
内部作業モデルの発達
愛着が確立されると、乳幼児は愛着対象と自己の感情的・認知的表象を発達させます。これらの「内部作業モデル」は、本質的に子供の環境を探索する能力を促進したり阻害したりします。なぜなら、子供は内部作業モデルに基づいて将来の出来事について予測を立てるからです。これは主に意識の外で起こります。子供が世界を安全だと認識する場合(安全な愛着によって最も一貫して支えられる感覚)、彼または彼女は社会的環境を含む環境の探索を始めます。このような状況では愛着システムは「オフライン」になり、子供は仲間との近接性を求め、社会化、模倣、食料調達技術の習得を奨励されることがあります。
逆に、環境を危険だと体験することは愛着行動を活性化します。これは暗闇、突然の動き、大きな音、高さ、または孤独感によって引き起こされます。喜びや愛などの肯定的な感情は、乳児に近さと安全によってもたらされますが、不安や悲しみなどの否定的な感情は、母親的存在からの認識された、または実際の分離によって生じます。分離の期間によって異なります。しかし、子供の否定的な反応は、愛情の絆の質や性質、および分離の期間に依存します。
分離に対する反応の段階
分離後の初期段階は通常、抗議によって支配されています。恐怖、怒り、泣き叫び、かんしゃくの表示は、おそらく人類の進化の過去における実際の脅威に対する適応反応として機能してきました。放置された乳児は捕食や怪我の大きなリスクにさらされていました。したがって、抗議は愛着対象が即座に戻るよう強制することを目的としています。
抗議段階は数時間から数日続くことがありますが、主な養育者が戻らなければ最終的に絶望の段階に移行します。絶望の段階は、絶望感、引きこもり、離脱の兆候の増加を特徴とします。運動活動は減少し、乳児は捕食者を引き寄せないよう適応として静かになります。多くの点でこの段階は深い悲しみやうつ病に似ています。
最終段階は分離(デタッチメント)を特徴とし、子供は積極的に環境に注意を向け、他の養育者を受け入れることがあります。しかし、主要な愛着対象が予期せず戻ってくると、子供は典型的に顔をそむけ、愛着対象を無視します。これに続いて、しがみつきと愛着対象の再喪失への恐れが交互に現れることがあります。進化的観点から見ると、分離は新しい愛情の絆への道を開くための最後の手段と見なすことができます。
愛着スタイルの種類
愛着理論家は4つの愛着スタイルを区別しており、これらは「ストレンジ・シチュエーション」と呼ばれる実験的に作成された分離シナリオで研究されています。このシナリオを通じて、乳児の分離苦痛と対処スタイルを体系的に検討することができます。生後12〜18ヶ月の乳児は、短時間母親と離れると異なる反応を示し、これは乳児の認識された安全性と強く関連しています。異なる愛着スタイルは、親の投資の量と質の違い、または危害回避や新奇性追求に関する乳児の生物学的素因の違いを反映している可能性があります。
安全な愛着
安全な愛着は、母親または他の愛着対象が乳児のシグナルに肯定的に応答できる場合、乳児が必要なときに母親を利用可能で反応的と認識するような形で最も容易に発達します。安全な愛着は、親が子供に投資する意欲と能力を反映しており、高い親の投資量を示しています。
典型的に、安全に愛着した子供たちは再会時に母親に近づき、近接性を求めますが、母親の存在下で遊びを再開することがあります。安全に愛着した子供たちは、好奇心を持って環境を探索できる安全基地または安全な避難所として母親を利用します。成人期には、安全な愛着は愛着関連の経験の一貫した描写として反映され、これは「自律的な心の状態」と呼ばれます。安全な愛着は文化を超えて最も一般的な愛着スタイルであり、進化の過去において積極的に選択されてきた可能性があります。
しかし、親の投資理論(第1章参照)に沿って、慰めの行動と利用可能性の量をめぐって子供と養育者の間に葛藤が生じる可能性があります。ストレンジ・シチュエーションのシナリオでは、例えば、一部の乳児が再会時に母親を避けたり無視したりすることが観察されています。回避的に愛着した乳児の愛着対象は、乳児のニーズに拒絶的で冷たい方法で反応することが発見されています。彼らは安全に愛着した子供の養育者と比較して、投資が少なく、子供に独立するよう促す傾向があります。乳児の回避反応は、放棄されるリスクを減らしながらある程度の近接性を維持する試みとして解釈できます。
成人として、回避的に愛着した個人は愛着関連の経験を理想化し、否定的な経験を軽視することがよくあります。これは現在の生活に対する初期の愛着の影響を最小化する試みと見なすことができます。この心の状態は「無視型」と呼ばれます。
アンビバレント(両価的)な愛着
「アンビバレント」または「抵抗的」と呼ばれる別の愛着スタイルは、再会時に乳児による接触追求と怒りの反応の混合によって特徴づけられます。アンビバレントに愛着した子供の多くの養育者は、その行動に一貫性がありません。回避的に愛着した子供の養育者が投資することを望まないことが多いのとは対照的に、アンビバレントに愛着した子供の養育者は、情緒的な利用可能性の欠如の機能として、投資することができないことがより多いです。
愛着対象は原則として乳児にとってアクセス可能かもしれませんが、子供がストレスを感じているときに慰めの行動を提供することができません。かんしゃくは、したがって、親の努力を最大化するための適応反応かもしれません。成人期では、アンビバレントな愛着は「とらわれ状態の心」に反映され、これは愛着に関連する経験を述べるときの曖昧さとして現れることがあります。
無秩序/方向喪失型の愛着
最後に、無秩序または方向喪失型の愛着スタイルは、再会時の子供たちの矛盾した行動のパターンによって特徴づけられます。回避は強い接触追求行動の後に続くか、同時に表示されることがあります。「凍結」は、無秩序な愛着の結果として発生する可能性のある別の形の行動です。苦痛の表現は、養育者から離れる方向への動きと共に生じる可能性があります。
成人として、無秩序に愛着した個人は「未解決」または「無秩序な心の状態」を表示し、これは愛着関係を記述するときの連想のゆるみや他の推論のずれとして明らかになります。無秩序に愛着した子供の愛着対象は、しばしば恐ろしいものとして経験されるか、または自分の乳児によって恐れを感じることがあります。虐待行動は無秩序な愛着を持つ乳児の養育者に最も一般的であり、これによってそれらの子供は異常に高いレベルの攻撃性、解離症状、うつ病、不安、または仲間やオブジェクトへの転嫁された攻撃性を含む様々な精神病理を発達させる特に脆弱になります。
このような行動は、愛着対象の利用可能性に対する認識された脅威への反応として説明することができ、恐怖、怒り、悲しみの否定的な感情がより広範になる可能性があります。
「不安定な愛着」の概念
後者の3つの愛着スタイルは、安全な愛着とは対照的に、「不安定な愛着」という用語の下にまとめることができます。不安定に愛着した子供たちは強い愛着行動を示すかもしれません—彼らは感情的な温かさと愛着対象の利用可能性のために苦闘するかもしれません。しかし、愛着行動の強さは、乳児と養育者の間の情緒的な絆の強さと混同してはなりません。
不安定に愛着した子供たちは、社会的環境がどのように反応するかについての期待を発達させ、これは安全に愛着した子供のものとは異なります。社会的相互作用の予想におけるこれらの違いは、内部作業モデルの違いと因果関係があります。例えば、拒絶を予想する子供は、慰められることを確信している子供と比較して、再会時に慎重に母親に近づくでしょう。
幼い乳児にとっての問題は、虐待的な養育者にさえ愛着を形成するよう強いられる可能性があること、そして脅威と危険の源である養育者に愛着を形成するというアンビバレントな状況に陥る可能性があることです。このような状況は激しい恐怖の感情状態を生み出す可能性があり、これはさらに愛着行動を増加させます。これは大きな不確実性と予測不可能性の状況を引き起こし、子供は安定した内部作業モデルを発達させることができないかもしれません。
メンタライジングと愛着の関係
不安定な内部作業モデルは、不安定に愛着した個人が他の人の考えや意図についてどのように考えるか(「メンタライジング」と呼ばれる、精神分析文献で使用される代替表現は「反射的機能」です;「社会的脳」の詳細については、第2章の後記を参照)にも深い影響を与えます。
逆説的な状況が発生する可能性があります。安全に愛着した個人は、安全に愛着した子供の養育者が不安定に愛着した子供の拒絶的な養育者よりも「心理的な」表現をより頻繁に使用する可能性があるため、他の人の精神状態について考えることをより速く学びます。一方で、安全と感情的な温かさの感覚は実際にメンタライジングシステムをオフにする可能性があります。
対照的に、不安定な愛着は発達的に遅れたり欠陥のあるメンタライジングシステムと結びつく可能性があり、これは否定的な感情と関連する状況でより頻繁に活性化される可能性があります。この逆説は、増強された疑い深さや妄想的反応を含む様々な精神病理学的な兆候や症状を引き起こす可能性があります(第17章を比較)。
同様に、子供と養育者の間の言語的コミュニケーションは、オープンなコミュニケーションを維持し、愛着対象の利用可能性への脅威を作り出す両方の新しい可能性を開くかもしれません。子供と養育者の間の愛着とコミュニケーションの違いは多くの方法で相互作用し、愛着スタイルは連続する世代にわたって「継承」される可能性があります。
遺伝子-環境相互作用
これらの違いは大部分が環境条件によって引き起こされますが、重要な遺伝子-環境相互作用の証拠がかなりあります。例えば、MAO-Aの活性を減少させるモノアミノオキシダーゼ阻害剤(MAO-AおよびMAO-B)コーディング遺伝子の特定の多型の保因者は、初期の生活で暴力、虐待、または放置などの逆境にさらされた場合にのみ、パーソナリティ障害や反社会的行動を発達させるリスクが高くなりますが、好ましい初期の生活条件下ではそのような関連は現れません。
言い換えれば、MAO-Aの活性だけでは行動の個人差を説明できませんが、遺伝的変異が発達の初期段階における社会的経験への異なる感受性を説明するという強い証拠があり、これらの相互作用は次のセクションで示されるように、パーソナリティの発達に深い影響を与えます。
日本語で高校の理科教科書のような平易な文体で翻訳いたします。
3. 思春期と若年成人期
生物学的な観点から見ると、思春期と成人期には大きな生理的・社会的変化が起こります。これらの変化は、個人が社会的な絆を形成し、恋愛関係を求めるための準備となります。
人間は非常に長い子ども時代と若年期を通じて、社会的スキルを身につけ、社会的・物理的世界についての内的作業モデル(心の中の世界観)を洗練させていきます。これによって個人は養育者からの自立が可能になります。重要なのは、養育者が乳幼児のニーズに敏感に応えることは、依存性を強めるのではなく、後の人生における自立を促すということです。これはよく誤解されている事実です。
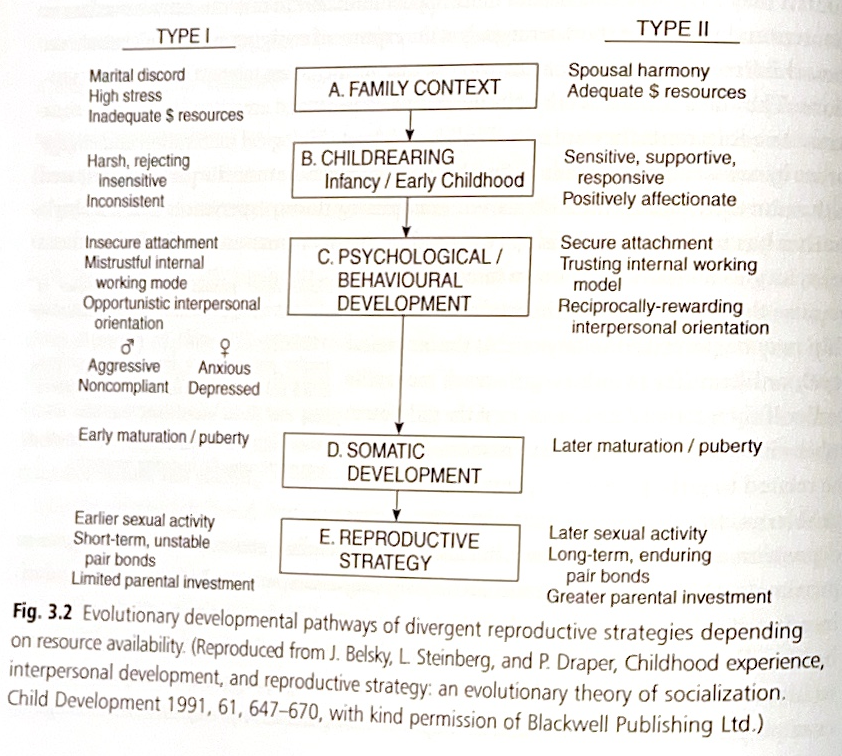
したがって、幼少期の経験、特に乳児期の愛着関係の性質が、その後の人生における友人関係や親密な関係、自分の子どもとの関係、さらには思春期の発達のタイミングや性行動、自分の子どもへの養育投資の量などに与える影響は、とても大きいといえます。これは、個人の初期の養育環境が発達中の内的作業モデルを決定し、その内的作業モデルが、資源の利用可能性や予測可能性、対人関係の信頼性や安定性についての個人の期待を導くからです。
ボウルビー(愛着理論の創始者)は当初、安定した愛着と信頼できる内的作業モデルだけが進化によって設計されたもので、他の形の愛着は標準からの逸脱だと考えていました。しかし進化的観点からすると、環境条件の変動—たとえ限られていたとしても—が、環境の偶然性に対する一連の柔軟な適応行動を選択してきた可能性が考えられます。これは最終的に、重要な生物社会的目標を達成するための異なる戦略を選ぶことで、繁殖の適応度を高めるためです。
例えば、幼少期に厳しい環境条件—実際のものであれ認識されたものであれ—を経験した場合、その個人は即座の資源獲得(他者の資源の利用を含む)や早期の交配・繁殖を目指す可能性が高くなります。なぜなら、その個人の内的作業モデルは将来の資源利用可能性の予測不可能性を示唆するかもしれないからです。
対照的に、養育者の情緒的な温かさと利用可能性があれば、他者は信頼でき、世界は比較的安全な場所であるという内的作業モデルが形成される可能性が高くなります。言い換えれば、人間は種のレベルでは極端なK戦略者(少数の子孫に多くの投資をする戦略)ですが、環境条件によって個人間のライフヒストリー戦略に違いが生じるのです。
子どもの視点から見て好ましくない養育条件は、心理的・生物学的な準備状態を引き起こし、その結果、青年期や成人期の行動が種の特性の範囲内でr選択(「量」重視)の方向へ傾く可能性があります。一方、豊富な資源と養育者の情緒的な利用可能性は、種の特性の範囲内でK選択(「質」重視)の方向性を強める可能性があります。
このことは、回避型や両価型の愛着が、ある程度、予測不可能な環境条件下での生存を確保し、繁殖成功を最大化するために進化した適応戦略を反映している可能性を示唆しています。
こちらも高校の理科教科書の文体で平易に翻訳します。
家庭環境と発達への影響
高いストレスレベルの家庭、母親からの拒絶、経済的資源の不足といった好ましくない環境で育った子どもたちは、他者は信頼できず機会主義的であるという不信感の強い内的作業モデルを発達させることがあります。
このような子どもたちは、思春期になると、最大限の資源を得るために、リスクの高い行動をとってでも短期的な目標を達成しようとする機会主義的な行動をとりやすくなります。そのため、不安定な愛着の背景を持つ子どもたちは、欲求不満の状況での忍耐力が低くなります。また、困っている仲間に対する共感も少なく、同年代から好かれにくいことがよくあります。
思春期になると、彼らは生物学的成熟が早まり、性行動も早くから始める傾向があります。成人になると、安定した愛着を持つ人と比べて、平均的に多くの親密なパートナーを持ちます。例えば、シングルマザーが複数のパートナーと親密な関係を持っているのを見て育った少女は、安定した関係を築く見込みが乏しいと感じるため、その母親の行動を自分の将来の行動モデルとして内面化するかもしれません。
生物学的レベルでは、他の霊長類とは異なり、少女たちは父親がいない場合や、ストレスの多い家庭環境にいる場合、初潮が早くなります。これは、少女たちが問題を内在化しやすい傾向と関連しているかもしれません。この内在化は悲しみ、うつ状態、社会的引きこもりを引き起こすだけでなく、代謝の低下と脂肪蓄積の増加をもたらし、それが初潮を早める直接的な原因となります。
若い成人として、不安定な愛着の背景を持つ女性は、複数の短期的な関係を持ち、性的な忠誠心が低く、自分が育てる意欲の低い子どもを産む可能性が高くなります。今日の10代の妊娠件数の爆発的増加は、このシナリオの一例といえるでしょう。
男子の場合も同様の発達が見られることがありますが、女子とは対照的に、問題を外在化しやすい傾向があり、それは高レベルの攻撃性、非協調性、衝動性として現れます。若い男性として、不安定な愛着を持つ個人は、無差別で機会主義的な性的関係を持つ可能性が高くなります。非ランダム(同類婚)の配偶により、うつ状態の女性は反社会的特性を持つ男性と結婚する可能性が高くなり、そのため低レベルの養育投資が「遺伝する」ように見えることもあります。
対照的に、夫婦の調和、低いストレスレベル、適切な経済的資源のある家庭で育った子どもたちは、より大きな情緒的温かさと養育者の利用可能性を経験することができます。これらの子どもたちは、信頼できる内的作業モデルを発達させ、世界を安全な場所として経験する可能性が高くなります。彼らの対人関係の姿勢は、より相互性を重視し、他者への信頼が強いです。
そのため、彼らは即座の資源獲得にあまり依存せず、報酬の遅延にもより耐えることができます。若年期には、安定した愛着を持つ個人は成熟が遅く、思春期に達する年齢も遅く、早期の性交渉を持つ可能性も低くなります。成人として、安定した愛着を持つ個人はより長期的な関係を持ち、自分の子どもにより多くの投資をする傾向があります(図3.2)。
進化的観点から見た愛着と生活史戦略
これらの正反対の生活史戦略は、種内でのr選択(多産少保護)とK選択(少産多保護)の間の極端なケースと見ることができ、その間にはあらゆる変化の可能性があります。
生物学的には、これらは同じ種の中の「モルフ」(形態型)を表しており、その頻度は個体群レベルで一定の範囲内に維持されています。集団全体で見ると、およそ3分の2の子どもたちは安定型の愛着を示し、20%が回避型の愛着、約10%が両価型(アンビバレント型)の愛着を持っています。これらの割合は文化的背景によってある程度変動します。無秩序型の愛着については、文化を超えたデータは得られていません。
いずれにしても、不安定な愛着スタイルはさまざまな精神病理(心の病気)のリスク要因になりますが、個人の遺伝的特性と重要な相互作用を持ちます。
すでに述べたように、養育スタイル(子育ての方法)は、母親と子どもの両方にとって予測される、あるいは実際に利用可能な資源に従って、世代を超えて「受け継がれる」可能性があります。将来の環境条件についての無意識の見積もりは現代生活の副産物ではなく、人類の進化の歴史全体を通じて当てはまってきたことです。
子育てにおける人間の母親同士の高いレベルの協力により、出産間隔を数年短縮することが可能になり、初期の人類の繁殖適応度を高めました。しかし、他の霊長類や類人猿とは異なり、人間の母親は同時に複数の依存している子どもを抱えるという負担を負います。これは、男性とのパートナーシップを強化する進化的メカニズムを発達させたからこそ可能になりました。
その中でも、排卵の隠蔽(いつ排卵しているかわからないこと)と強い感情的な絆が「核家族」の形成に最も貢献してきました。さらに、人間の女性は自分自身の生殖期間を超えて特別な役割を担うように進化してきました。これについては次の章で扱われます。
4. 生殖後期間
人間生物学における大きな謎のひとつは、女性が閉経後も20~30年も生存するという進化的なパズルに関連しています。一見すると、これは繁殖適応度の観点からは説得力がないように思えます。閉経自体は動物界では珍しくありませんが、人間の女性の長い生殖後寿命は特異的です。
人間の生殖能力は初潮から徐々に増加し、30歳前後でピークに達した後、着実に低下し、最終的には停止します。他の類人猿と同様に、人間の女性も約50歳で排卵を停止します。これは適応的であり、年齢とともに流産や遺伝的な先天異常のリスクが高まるからです。
しかし、他の霊長類とは異なり、人間の女性だけがさらに20~30年生きます。この生活史のパターンは、現代の生活環境の副産物ではありません—狩猟採集社会で60歳に達した女性は、さらに10~20年生きる可能性が高いのです。人間の長い閉経後寿命は、おそらくすべての段階で全体的に延長された生活史と密接に関連しています。
人間は繁殖を始めるまでに約20年かかり、親世代への依存期間もそれに応じて長いです。この観点からすると、50歳を超えて繁殖することは意味がありません。なぜなら、最後の子孫が独立するまで生存を維持することが難しくなるからです。
さらに、自然選択は近親者への利他主義を優先してきたと考えられています。閉経後の女性が追加の資源を提供することは、特に自分の子の子ども(孫)の生存を助けました。この状況は「祖母仮説」として知られています。
実際、狩猟採集社会における生殖後の女性は、自分が消費できる以上の食料資源を部族に提供しています。彼女たちは生殖年齢の女性よりも多くの時間を食料の収集や加工に費やします。ほとんどの狩猟採集社会(人間の祖先の状態のモデルとして)では、このような余剰は主に自分の親族の子孫、特に孫の利益になります。
これは進化的に理にかなっています。なぜなら、祖先環境では離乳したばかりの子どもたちは飢餓のリスクがあり、祖母からのカロリー追加供給が彼らの生存確率を高めたからです。したがって、老化の遅延と寿命の選択は、まず第一に、非常に減速した人間のライフサイクルと、高カロリーの食事を追加で提供することによって、非常に依存的ではあるが既に離乳した子孫の生存可能性を高める必要性に関連していたかもしれません。
悲しいことに、現在我々はアフリカでHIV孤児の悲劇を目の当たりにしています。彼らの両親がエイズで亡くなった後、彼らの祖母たちは、依存している孫たちに養育とケアを提供するという点で、実際に大きな価値を持っています。
生殖後の女性は、価値ある資源の場所や季節変動についての知識、および紛争解決を含む高度な社会的スキルについても評価されていたでしょう。同様に、年配の男性も部族社会内での狩猟スキルや社会的問題の交渉において、同様の理由で価値があったかもしれません。
この仮説に沿って、意味記憶の貯蔵に必要な結晶性知能は高齢になっても安定していることは注目に値します。一方、流動性知能と認知的柔軟性は年齢とともに低下します。
しかし、採集への依存度が低い人間文化では、年配の女性への尊敬は大幅に低くなっていました。つい最近まで、食事に多量の肉を含む部族社会では、女性が食料の提供にあまり貢献できなかったため、年配の女性の安楽死(社会的に認められた男性によって実行される)さえ行われていました。
このような否定的側面は、おそらく母親と子どもの間の特別な愛情の絆、そして人間における利他主義と血縁関係の重要性によって相殺されていました。養育者への愛着は、逆に役割の逆転を促すかもしれません。これを通じて、高齢の親は若い大人、通常は自分の成人した息子や娘との近さを求めます。
しかし、高齢者の価値下げは確かに狩猟採集社会に特有のものではありません。現代文化は多世代家族の崩壊をもたらし、その結果として高齢者が家族生活に関与できないことは、おそらく高齢者層における非常に高い鬱病率の原因のひとつでしょう。
後記:出生前環境と生物心理社会的準備
母親の出生前ストレスに関する研究が増えており、子どもが後の人生で経験する心理的問題が、出生前のストレス要因の強さとタイミングに関連している可能性が示唆されています。
「胎児プログラミング」という概念は、子宮内環境が胎児の体を出生後の条件に備えさせるということを意味しています。例えば、妊娠中の母親のうつ病や不安は、低出生体重や早期分娩と関連しており、また学齢期の子どもの行動問題とも関連しています。
例として、妊娠12~22週の間に母親が強い否定的感情を持つと、8~9歳と14~15歳の子ども(女子より男子に多い)の衝動性と多動性を予測できることがわかっています。さらに、母親の出生前ストレスは、子どもの不安症状、外在化問題(攻撃性など外に向かう問題行動)、注意欠陥を予測することもあります。
興味深いことに、出生前ストレスは脳の左右差の減少も予測し、これは自閉症や統合失調症と関連していることがわかっています。この統合失調症の病態生理学的メカニズムの可能性は、妊娠第1期に近親者を亡くした母親のストレスが、子どもの統合失調症リスクを有意に高めるという最近の観察によって裏付けられています。
母親のストレス体験が子どもの行動に深刻な影響を与える可能性があることは明らかですが、ストレス要因の特異性とタイミングについてはあまり意見が一致していません。長期間にわたる慢性的なストレスは、一回限りのストレスイベントよりも胎児に大きな影響を与えると考えるのが妥当でしょう。夫婦間の不和や別居は特に重要と思われます。
母親と胎児の血漿コルチゾールレベルは高い相関関係にあることが示されており、動物モデルでは、出生前ストレスに関連するエピジェネティック要因(メチル化)が海馬(記憶を司る脳の部位)のグルココルチコイド受容体の発現を減少させる可能性があることが示されています。これにより、視床下部-下垂体軸(HPA)を介したストレス誘発コルチゾール反応のフィードバック制御が損なわれる可能性があります。
これらの知見に一致して、出生前ストレスは海馬の体積縮小と神経新生の減少、HPA機能の変化、および非ヒト霊長類における新奇刺激や社会的孤立に対する慢性的に上昇したストレス反応をもたらします。これらの影響は、ストレス暴露が妊娠初期に起こると霊長類において顕著であるようです。しかし、人間の脳成長の延長期間と生理学的早産を考慮すると、人間の胎児や乳児の脳は、かなり長い期間、母親のストレスに対して脆弱である可能性が考えられます。
これらのシナリオは、r選択(多産少保護)対K選択(少産多保護)戦略の種内変異モデルの論理的拡張のように思えます。進化的適応環境(EEA)において、社会的理由や危険が増大した時期に、より苦悩し、不安で警戒心の強かった人間の母親は、予測不可能な環境条件に生まれてくる子どもを準備させ、その結果、子どもの行動を即時的な資源獲得と機会主義的行動というr選択の方向にわずかに押し進めたかもしれません。
もう一つの例(ある程度推測的かもしれませんが)は、人間の心理と行動の関係を説明する生物学的原理の妥当性を示しています:ニューヨーク市でのテロ攻撃の2ヶ月後、女児に比べて男児の胎児死亡率が不釣り合いに増加しました。この発見は第1章で概説されたトリバース・ウィラード仮説を思い起こさせます。女性は(もちろん無意識のうちに)、出生後の繁殖適応度の見通しが女児の方が大きいと認識されたため、選択的に男性胎児を流産させたのでしょうか?これらの仮説的な因果関係は、将来的に前向き研究で調査される必要があります。
Selected further reading
Bakermans-Kranenburg, M. J. & van Ijzendoorn, M. H. (in press), ‘Behavioral and molecular genetics
of attachment security and disorganization: genetic vulnerability or differential susceptibility?’
Behavioral and Brain Sciences.
Belsky, J., Steinberg, L. & Draper, P. 1991, ‘Childhood experience, interpersonal development,
and reproductive strategy: an evolutionary theory of socialization, Child Development, vol. 62,
pp. 647-670.
Bowlby, J. 1969, Attachment and Loss. Vol. 1. Attachment, Basic Books, New York.
Bowlby, J. 1973, Attachment and Loss. Vol. 2. Separation: Anxiety and Anger, Basic Books, New York.
Bowlby, J. 1980, Attachment and Loss. Vol. 3. Loss, Basic Books, New York.
Bradshaw, J. 1997, Human Evolution. A Neuropsychological Perspective, Psychology Press, Hove.
Carter, A. S., Garrity-Rokous E., Chazan-Cohen, R., Little, C. & Briggs-Gowan, M. J. 2001,
‘Maternal depression and comorbidity: predicting early parenting, attachment security,
and toddler social-emotional problems and competencies’, Journal of the American Academy of
Child and Adolescent Psychiatry, vol. 40, pp. 18-26.
Cassidy, J. & Shaver, P. R. (eds) 1999, Handbook of Attachment. Theory, Research, and Clinical
Applications, Guilford Press, New York.
Fonagy, P. 1991, ‘Thinking about thinking: some clinical and theoretical considerations in the
treatment of a borderline patient, International Journal of Psychoanalysis, vol. 72, pp. 639-656.
Harlow, H. F. & Zimmermann, R. P. 1959, ‘Affectional responses in the infant monkey’, Science,
vol. 130, pp. 421-432.
Hrdy, S. B. 2000, Mother Nature, Vintage, London.
Insel, T. R. 1997, ‘A neurobiological basis of social attachment’, American Journal of Psychiatry,
vol. 154, pp. 726-735.
Kraemer, G. W. 1997, ‘Psychobiology of early social attachment in rhesus monkeys. Clinical
implications’, Annals of the New York Academy of Sciences, vol. 807, pp. 401-418.
Noftle, E. E. & Shaver, P. R. 2006, ‘Attachment dimensions and the big five personality traits:
associations and comparative ability to predict relationship quality’, Journal of Research in
Personality, vol. 40, pp. 179-208.
Pearce, J. W. & Pezzot-Pearce, T. D. 2001, ‘Psychotherapeutic approaches to children in foster care:
guidance from attachment theory’. Child Psychiatry and Human Development, vol. 32, pp. 19-44.
Shaver, P. R. & Mikulincer, M. 2005, ‘Attachment theory and research: resurrection of the
psychodynamic approach to personality’, Journal of Research in Personality, vol. 39, pp. 22-45.
Simpson, J. A. 1999, ‘Attachment theory in modern evolutionary perspectives’ in Handbook of
Attachment. Theory, Research, and Clinical Applications, eds J. Cassidy & P. R. Shaver,
Guilford Press, New York.
Talge, N. M., Neal, C. & Glover, V. 2007. ‘Antenatal maternal stress and long-term effects on child
neurodevelopment: how and why?’, Journal of Child Psychology and Psychiatry, vol. 48, pp. 245-261.
Van Ijzendoorn, M. H. & Sagi, A. 1999, ‘Cross-cultural patterns of attachment: universal and contextual
dimensions’, in. Handbook of Attachment. Theory, Research, and Clinical Applications, eds J. Cassidy
& P. R. Shaver, Guilford Press, New York.
Voland, E. 1998, ‘Evolutionary ecology of human reproduction’, Annual Review of Anthropology,
vol. 27, pp. 347-374.
サイドメモ
ライフヒストリー理論:進化と発達の概要
ライフヒストリー理論は、個体の生涯にわたって変化する生存と繁殖に関連した問題に対する、種に特有の解決策を扱います。K選択種は以下の特徴を持ちます:
- ゆっくりと発達する
- 繰り返し繁殖する(反復繁殖)
- 一度の出産あたりの子どもの数が少ない
- 長期間の集中的な親の投資がある(長い妊娠期間を含む)
- 乳幼児期の死亡率が低い
- 繁殖活動の開始が遅れる
- 長寿命である
人間のライフヒストリーはこの発達パターンに従います。
しかし、ライフヒストリーのパターンには種内変異があり、一部の人間のライフヒストリーはややr選択型のパターンに押し進められ、他はより極端なK選択型のパターンに向かうこともあります。前者は人間が精神病理(心の病気)を発症しやすくなる可能性があります。
人間の乳児の特徴
人間の赤ちゃんは他の類人猿と比較して約13ヶ月早く生まれます。この誕生時の未熟さは、二足歩行による骨盤の解剖学的変化に関連した設計上の妥協を表しています。人類の進化における脳の拡大は、早期出産への圧力を高めました。
人間の赤ちゃんは、誕生時に目や耳が開いていて、主要な養育者と交流する能力があるという早熟な特徴を示します。しかし同時に、動けないことや裸であることなどの晩成的特徴があり、これには集中的な親のケアが必要です。
愛着の重要性
母親と子どもの間の二者関係(ダイアド)の形成は、初期の人類における最も重要な心理的適応の一つとなりました。
愛着の重要性は、若いサルを対象とした多くの研究で実験的に示されています。養育者からの隔離期間によって、若いサルはうつ病の重症な形態を発症します。成体になると、これらのサルは緊張し、自分の子どもを育てることができなくなります。
人間の乳児は、養育者との近さを達成するためのいくつかの手段を持っています:
- 生後4週間で、人間の顔の輪郭に対する強い好みを示す
- 人間の声に反応する
- 泣いたり笑ったりして積極的に接触を求める
- 赤ちゃんの笑顔や遊び顔は、他の霊長類よりも発生的にはるかに早く現れる
人間の乳児は、大きな目、丸い顔や頭蓋骨、短い手足など、通常は攻撃性を減少させ、「かわいい」と認識される特徴を示します。しかし、母親の子どもの信号に対する応答性は、乳児側の近接探索行動ほど生まれつきプログラムされているわけではありません。そのため、「愛着」とは対照的に、「絆形成(ボンディング)」という用語が、安定した母子二者関係の発達を支援する母親の行動を説明するために選ばれました。
愛着と絆形成が確立されない場合、子殺しが発生する可能性があります。
愛着は母親や父親など複数の人物と形成されることもありますが、通常は階層的な方法で行われます。
内的作業モデル
発達する愛着対象と自己の情緒的・認知的表象は、内的作業モデルと呼ばれます。これらは子どもが環境を探索する方法を決定し、子どもは内的作業モデルに基づいて将来の出来事についての予測を行います。
愛着対象からの分離は、子どもに抗議、絶望、そして最終的には愛着対象への関心の喪失をもたらします。
愛着の種類
愛着理論は4つのタイプの愛着を区別し、これらは主要な養育者の情緒的利用可能性と応答性の量に依存します。
安定型の愛着は、愛着対象が乳児の信号に肯定的に応答できる場合に発達し、乳児は母親が必要なときに利用可能で応答的であると認識します。
不安定な愛着を持つ子どもたちは、近接を確保し、見捨てられることを避けることを目的としたさまざまな行動を示します。不安定な愛着は、回避型、両価型(抵抗型)、無秩序型の行動パターンに分けられます。
不安定な愛着の結果として生じる不安定な内的作業モデルは、個人が他者の考えや意図について考える方法に深い影響を与えます。これは「メンタライジング(心を読む能力)」と呼ばれます。安定した愛着を持つ個人は、他者の心的状態について考えることをより速く学びます。一方、安全感と情緒的な温かさは、実際にメンタライジングシステムをオフにする可能性があります。
遺伝と環境の相互作用
遺伝的変異は、発達の初期段階での社会的経験に対する感受性の違いを説明し、これらの遺伝子-環境相互作用は人格発達に深い影響を与えます。
養育者の情緒的利用可能性は、依存性を強化するのではなく、後の人生での子どもの独立性を促進します。
(子どもの視点から見て)好ましくない養育条件は、r選択(「量」)に向けた心理的および生物学的準備を引き起こす可能性があります。一方、豊富な資源と養育者の情緒的利用可能性は、種に特有の境界内でK選択の方向性(「質」)を高める可能性があります。
思春期と成人期
他の霊長類とは異なり、父親の不在やストレスの多い家庭環境は初潮を早めます。これは、少女たちが問題を内在化しやすい傾向と関連している可能性があります。この内在化は悲しみ、うつ状態、社会的引きこもりを生み出すだけでなく、代謝の低下と脂肪蓄積の増加をもたらし、それが初潮を誘発する直接的な原因となります。
安定した愛着を持つ個人は成熟が遅く、思春期に達する年齢も遅く、早期の性交渉を持つ可能性も低くなります。成人として、安定した愛着を持つ個人はより頻繁に長期的な関係を持ち、自分の子どもにより多くの投資をする傾向があります。
文化横断的比較では、約3分の2の子どもが安定型の愛着を持ち、20%が回避型の愛着、約20%が両価型の愛着を持ちます。無秩序型の愛着については、文化横断的なデータは得られていません。
生殖後期間
進化は、女性の長い生殖後(閉経後)寿命を選択してきました。なぜなら、閉経後の女性が孫の生存に貢献したからです(「祖母仮説」)。
現代における高齢者の軽視は、高齢者層における例外的に高いうつ病率の理由の一つかもしれません。
