
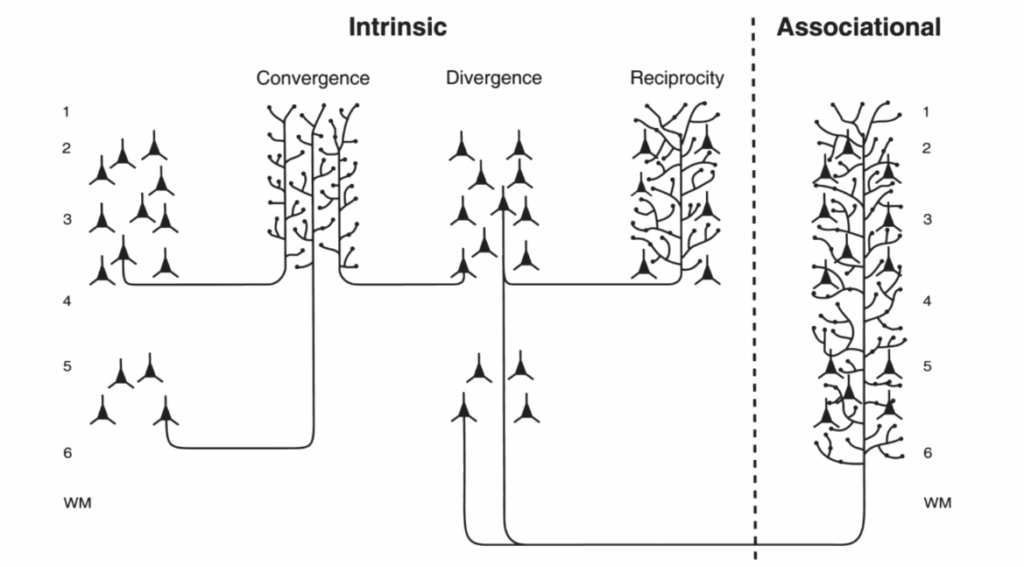
図1.2-4. サル前頭前皮質における内在性および連合性接続の空間的組織と、これらの接続における相互性、収束性、拡散性を比較した模式図。(Pucak ML, Levitt JB, Lund JS, Lewis DA. Patterns of intrinsic and associational circuitry in monkey prefrontal cortex. J Comp Neurol. 1996;376(4):614-630. Copyright @ 1996 Wiley-Liss, Inc.より許可を得て改変。)
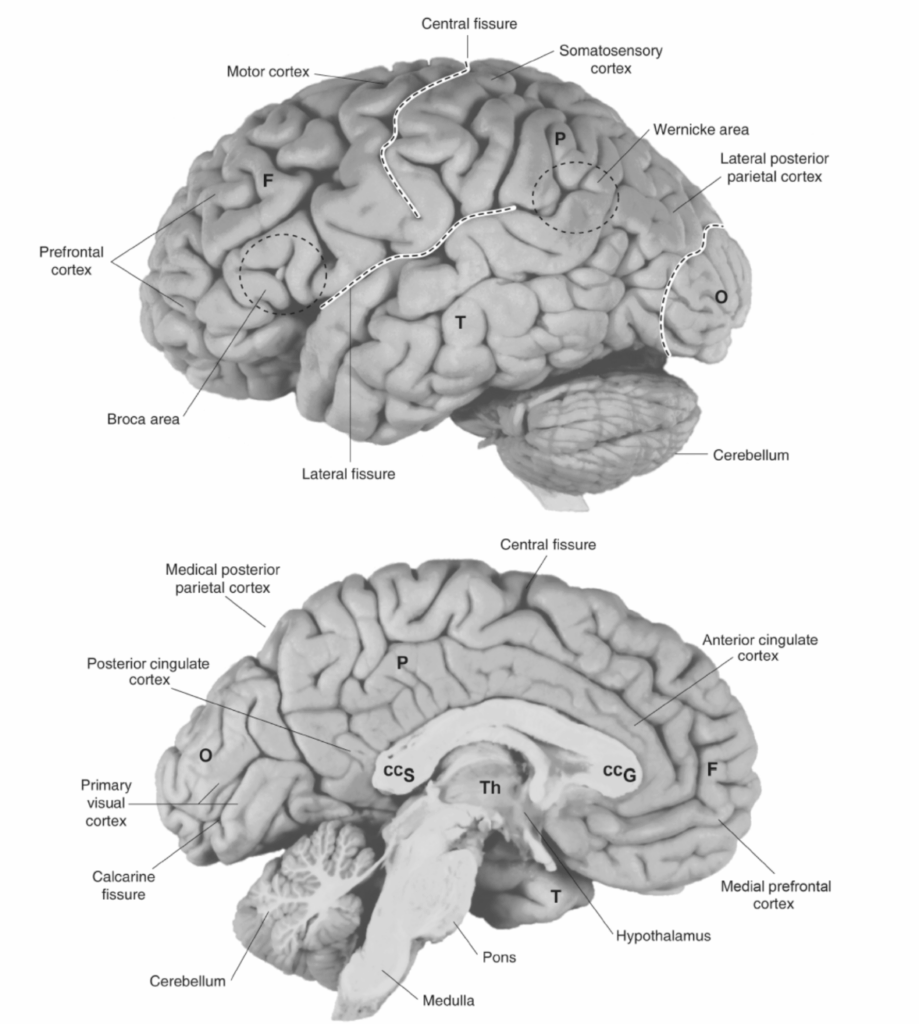
図1.2-6. 神経管の小胞とその派生物の模式的表現を伴う、ヒト脳の内側面図。
神経接続と人間の脳の特異性
神経接続の組織原理
拡散性と収束性の接続
第二に、多くの神経接続は拡散性または収束性の性質を持っている。拡散性システムは、一つのニューロンまたは離散したニューロン群から、脳の多様な部分に位置する可能性のあるはるかに多数のニューロンへの情報伝達を含む。青斑核は、脳幹にある少数のノルエピネフリン含有ニューロン群で、大脳皮質全体および他の脳領域に軸索投射を送る、高度に拡散性のシステムの例である。対照的に、複数の脳領域からの出力が単一の領域に向けられ、収束性システムを形成することがある。大脳皮質の複数の連合野から内側側頭葉の嗅内領域への投射は、収束性システムの例である。脳領域内の接続も拡散性と収束性を示す(図1.2-4参照)。例えば、サルの前頭前皮質では、個々の縞内の錐体ニューロンは他のいくつかの縞に投射する軸索を持ち(拡散性)、個々の縞は複数の縞から入力を受ける(収束性)。このシステムにおける拡散性は、空間的に制限された入力が、特定の反応を生成するために協調的な活性化が必要なニューロン群を動員することを可能にする解剖学的基盤を提供する可能性がある。このシステムにおける収束性は、縞の配列に存在する異なるモダリティからの情報を単一の場所に中継することを可能にし、それらの情報内容の統合を促進する可能性がある。
階層的・並列的組織
第三に、領域間の接続は階層的または並列的な様式、あるいはその両方で組織化されている可能性がある。視覚入力は、網膜の複数のニューロン集団から外側膝状体、一次視覚野、そして段階的に大脳皮質の複数の視覚連合野へと、連続的または階層的な様式で伝達される。階層的スキーム内では、異なるタイプの視覚情報(例:運動と形態)が、視覚系の異なる部分を通じて並列的に処理される可能性がある。
脳領域の機能特化
最後に、脳の領域は異なる機能に特化している。左下前頭回(ブローカ野)(図1.2-5)の病変は、言語産生における特徴的な障害を引き起こす。言語は複雑な機能である。しかし、それはブローカ野の完全性だけでなく、拡散性と収束性、連続的、並列的相互接続を通じた多数の脳領域にわたる情報の分散処理にも依存している。したがって、特定の行動の産生や特定の神経精神障害の病態生理学における任意の特定の脳領域またはニューロン群の役割は、孤立して見ることはできず、ニューロンを他の脳領域と接続する神経回路の文脈内で考慮されなければならない。
人間の脳の特異性
構造的特徴
他の霊長類種の脳と比較して、人間の脳は実質的に大きく、特定の領域が不均衡に拡大している。前頭前皮質は、猫では全皮質容積のわずか3.5パーセント、サルでは11.5パーセントを占めると推定されているが、人間の脳ではより大きな皮質容積の30パーセント近くを占めている。逆に、他の領域の相対的表現は人間の脳では減少している。例えば、一次視覚野は人間では大脳皮質の全面積のわずか1.5パーセントを占めるが、サルではより多くの割合(17パーセント)の大脳皮質がこの領域に割り当てられている。したがって、人間の脳の特異性は、そのサイズと、特に高次認知機能に関与する大脳皮質領域の特定領域の差異的拡大に起因している。
神経回路の組織の違い
さらに、人間の脳の拡大と分化は、神経回路の特定要素の組織における実質的な違いと関連している。例えば、齧歯類と比較して、人間の大脳皮質のドーパミン作動性神経支配ははるかに広範囲で領域特異的である。一次運動野と特定の後頭頂領域は、サルと人間では密なドーパミン神経支配を受けるが、ラットではこれらの領域はドーパミン入力をほとんど受けない。
細胞レベルでの特異性
人間の脳は細胞レベルでも特異的で、大脳皮質における特定のニューロンおよびグリア細胞のより大きな相対的パーセンテージを持つ。例えば、GABA作動性介在ニューロンの相対的割合は、齧歯類の大脳皮質では約15パーセントであるが、人間では、ほとんどの皮質領域のすべてのニューロンの20から25パーセントがGABA作動性である。さらに、分子特性解析により、齧歯類の大脳皮質には見られない人間の大脳皮質のニューロンの亜型が明らかになっている。
アストロサイトの重要性
層内および極性アストロサイトは人間にのみ見られ、人間の原形質性アストロサイトは齧歯類の対応物よりも大きく、より多くの突起を持つ。さらに、アストロサイト対ニューロン比は人間では齧歯類と比較して5倍大きく、ニューロンとシナプスのより大きなアストロサイト被覆をもたらし、したがって人間の大脳皮質における神経伝達のより強化された調節を可能にしている。人間の脳機能におけるアストロサイトのより大きな貢献のこれらの発見は、アストロサイトが高次脳機能にとって重要であり、人間の精神障害において脆弱である可能性があることを示唆している。
結論
したがって、人間の脳は大解剖学、細胞構成要素、分子組成における違いを示し、系統発生学的に下位の種から区別する特徴を持っている。
人間の脳研究における限界
これらのタイプの種差は、齧歯類や非ヒト霊長類の研究を推論の基盤として使用する際に、人間の脳機能に関して行われる一般化の精度に限界があることを示している。しかし、人間の脳の組織の直接的な調査は、明らかに多数の要因によって制限され、複雑化されている。
前述のように、人間の脳の拡大は大脳皮質の追加領域の出現と関連している。例えば、人間の内側側頭葉の嗅内皮質は時として単一の皮質領域と見なされるが、この皮質の細胞構築と化学構築はその吻尾側範囲に沿って実質的に異なっている(図1.2-3参照)。これらの領域を他の構造との相対的位置によって同定することは魅力的であるが、人間の脳には十分な個体間変動があり、そのような位相学的定義を信頼できないものにしている。嗅内皮質の場合、扁桃体や海馬などの隣接構造に対するその異なる細分化の位置は、人間の脳全体で変動する。したがって、すべての研究、特に人間の脳を使用する研究では、関心領域は、研究者がすべての症例で同じ領域を正確に同定できるような方法(例:細胞構築、化学構築、髄鞘構築的特徴を使用)で定義されなければならない。
人間の脳の研究に対する追加の制限は、死亡時から脳標本の凍結または固定までの間隔中に生じる可能性のある形態学的および生化学的変化に関するものである。既知の死後間隔の影響に加えて、そのような変化は死に先立つ瀕死状態中に生じ始める可能性がある。人間の脳の組織を他の種のものと比較する際、研究者は死後遅延または瀕死状態の結果として人間の脳に生じた可能性のある変化を考慮しようとしなければならない。疾患状態の研究では、神経伝達物質含量またはその他の特性における症例間の違いが、組織調製方法などの疾患状態以外の要因の結果である可能性があるため、適切な対照が使用されなければならない。ポジトロン断層撮影(PET)、磁気共鳴画像法(MRI)、磁気共鳴分光法(MRS)、拡散テンソル画像法(DTI)などの画像化技術を使用した生体内での人間の脳の研究は、これらの問題の多くを回避するが、人間の脳組織の多くの側面の研究には不十分な解像度によって制限される。
構造成分
主要な脳構造
人間の脳発達の初期段階では、神経管内に3つの一次小胞を同定することができる:前脳、中脳、菱脳(図1.2-6)。その後、前脳は分裂して終脳と間脳になる。終脳は大脳皮質、海馬体、扁桃体、および大脳基底核の一部の成分を生じる。間脳は視床、視床下部、およびいくつかの他の関連構造になる。中脳は成人脳の中脳構造を生じる。菱脳は後脳と髄脳に分裂する。後脳は橋と小脳を生じ、延髄は髄脳の派生物である。
各半球の大脳皮質は4つの主要な領域に分けられる:前頭葉、頭頂葉、側頭葉、後頭葉(図1.2-5参照)。前頭葉は中心溝の前方に位置し、一次運動野、運動前野、前頭前野領域から構成される(図1.2-7)。前頭前皮質は背外側と腹外側領域に分けることができ、これらの領域はそれぞれ異なる機能特性を持つ。例えば、背外側前頭前皮質(DLPFC)は、作業記憶中の情報の純粋な維持により関与している腹外側前頭前皮質よりも、作業記憶課題中のデータの操作により関与しているようである。一次体性感覚野は前頭頂葉に位置し、さらに、複雑な視覚および体性感覚機能に関連する他の皮質領域が後頭頂葉に位置している。側頭葉の上部は一次聴覚野および他の聴覚領域を含み、下部は複雑な視覚機能に関与する領域を含む。さらに、上側頭溝内の一部の領域は、視覚、体性感覚、聴覚の感覚領域からの入力の収束を受ける。後頭葉は一次視覚野および他の視覚連合野から構成される。人間および他の類人猿の脳はまた、島皮質と呼ばれる主要な領域を含み、これは外側溝の深部に位置する皮質領域である(図1.2-8および1.2-9)。背側は前頭および頭頂弁蓋によって、腹側は側頭弁蓋によって覆われている。人間の島皮質は脳回を有し(回転状)、平滑である非ヒト霊長類の島皮質とは異なる。人間の島皮質は他の皮質領域、扁桃体、視床との広範な接続を持つ。
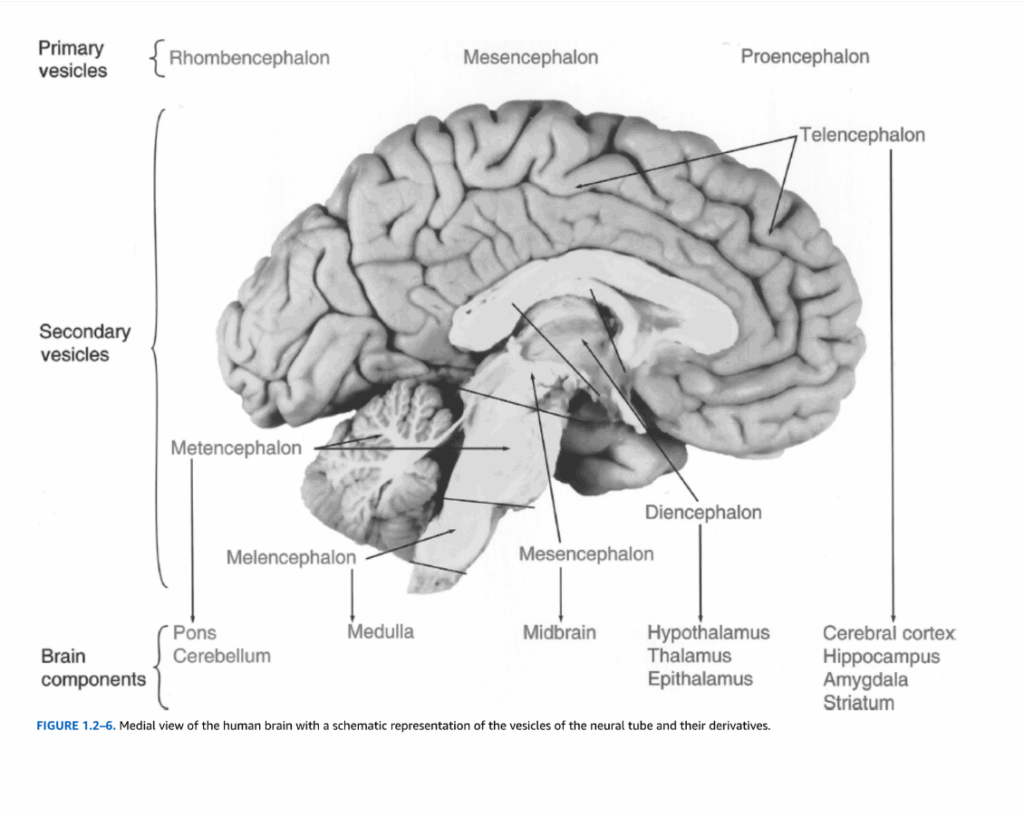
図1.2-6. 神経管の小胞とその派生物の模式的表現を伴う、ヒト脳の内側面図。

図1.2-7. ヒト脳の脳梁膝部直前の冠状断面図。下部の挿入図は断面のレベルを示す。IFG、下前頭回;MFG、中前頭回;PFC、前頭前皮質;SFG、上前頭回。(Nieuwenhuys R, Voogd J, Huijzen C. Brain slices. In: The Human Central Nervous System: A Synopsis and Atlas. 4th ed. Springer Nature, 2008:141.より許可を得て改変。)
大脳皮質の外側外套の下には、尾状核、被殻、淡蒼球など、多くの他の主要な脳構造がある(図1.2-8参照)。これらの構造は大脳基底核の構成要素であり、運動の制御と特定の認知過程に関与するシステムである。海馬と扁桃体は、辺縁系の構成要素で、内側側頭葉の深部に位置している(図1.2-9から1.2-11)。さらに、視床や視床下部などの間脳の派生物は、顕著な内部構造である。視床は大脳皮質との明確な接続パターンを持つ多数の核から構成される比較的大きな構造である(図1.2-9から1.2-11参照)。対照的に、視床下部は自律神経系および内分泌機能に関与するはるかに小さな構造である。
白質線維束
大脳半球は数十億の有髄軸索または線維を含み、白質にその特徴的な色を与え、大脳皮質への情報および大脳皮質からの情報を運ぶ。これらの軸索は投射線維、交連線維、連合線維を含む白質線維束に束ねられている。
投射線維 主要な投射線維系のうち2つは、大脳皮質に起源を持ち皮質下標的に投射する線維(皮質遠心性)と、終脳外に起源を持ち大脳皮質に投射する線維(皮質求心性)から構成される。これらの例は、それぞれ皮質視床投射と視床皮質投射である。これらの投射線維は内包を通り、視床とレンズ核(すなわち、被殻と淡蒼球を一つの構造として考えたもの)と構造的に関連した緻密な線維束である。各大脳半球において、内包は外側はレンズ核によって、内側は視床と尾状核頭部によって境界づけられている(図1.2-12)。皮質橋路、皮質脊髄路、皮質延髄路などの他の線維系は、皮質から内包と大脳脚を通って下降し、橋、脊髄、脳幹のそれぞれの目的地に到達する。内包を通るすべての線維は放線冠を形成し(図1.2-8参照)、内包の直上に位置する扇状の構造である。
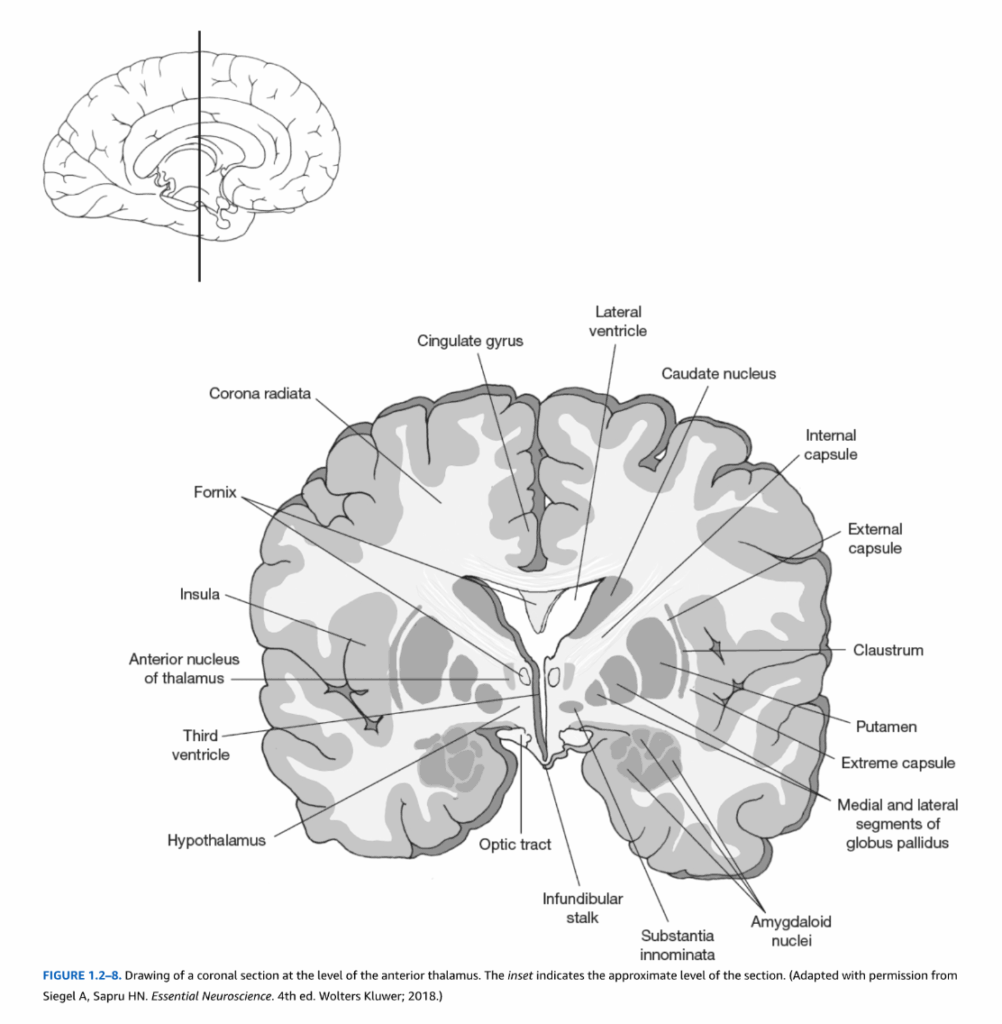
図1.2-8. 視床前部レベルでの冠状断面図。挿入図は断面のおおよそのレベルを示している。(Siegel A, Sapru HN. Essential Neuroscience. 4th ed. Wolters Kluwer; 2018より許可を得て改変)
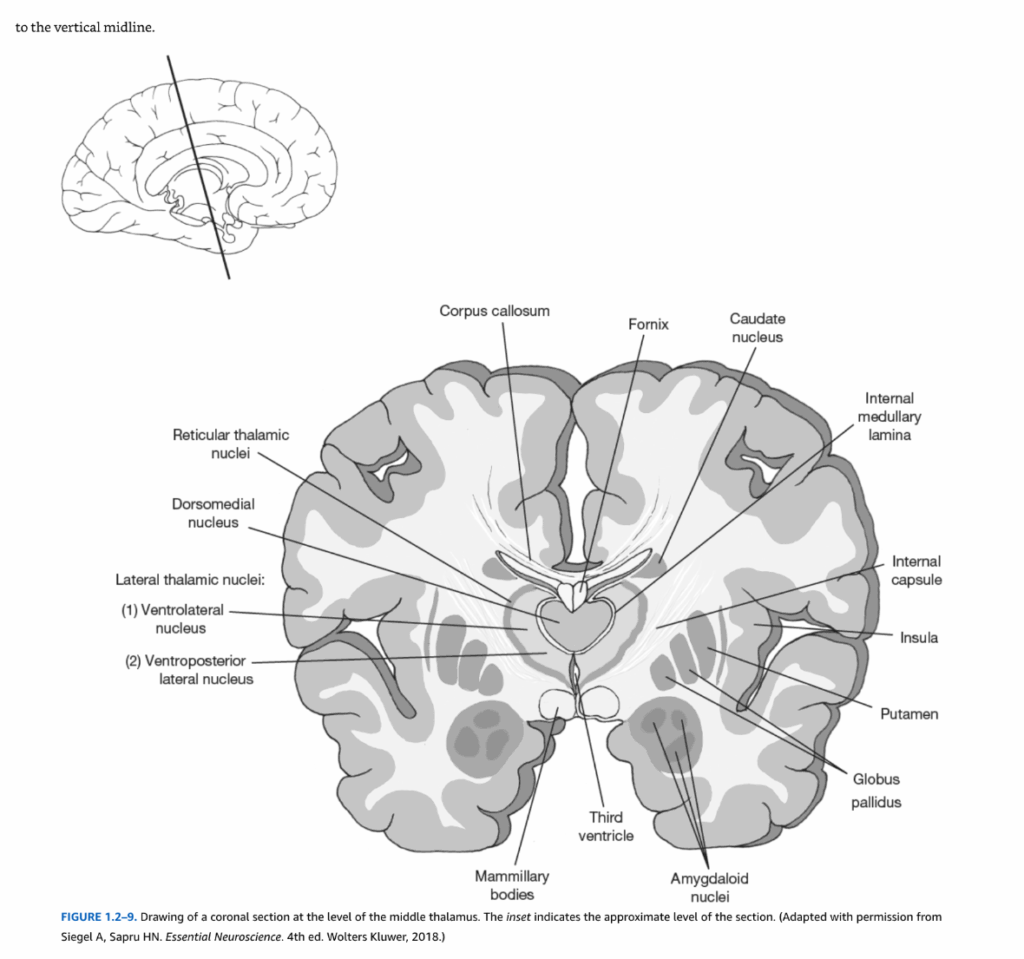
図1.2-9. 視床中部レベルでの冠状断面図。挿入図は断面のおおよそのレベルを示している。(Siegel A, Sapru HN. Essential Neuroscience. 4th ed. Wolters Kluwer, 2018より許可を得て改変)
脳梁は4つの部分から構成されている(図1.2-13)。脳梁は嘴部から始まり、前方および背方に湾曲して膝部を形成する。脳梁体部は最も大きな部分であり、拡大した丸みを帯びた後端部である膨大部へと続く。脳梁体部と膨大部の間の狭い部分は峡部と呼ばれることもある。前頭前野、側頭葉、頭頂葉皮質からの高次認知・感覚情報を運ぶ軸索は主に膝部と膨大部を通過するが、視覚、聴覚、体性感覚情報は主に脳梁体部と峡部を通って運ばれる。
前交連は脳梁の尾側にある繊維の密集した束で、脳弓の前方で正中線を横断している(図1.2-8参照)。前交連は両側の側頭葉領域と前嗅核からの繊維を相互に結んでいる。より小さな交連繊維束には、間脳の尾側部分を結ぶ後交連と、両側の海馬体を相互に結ぶ海馬交連がある。
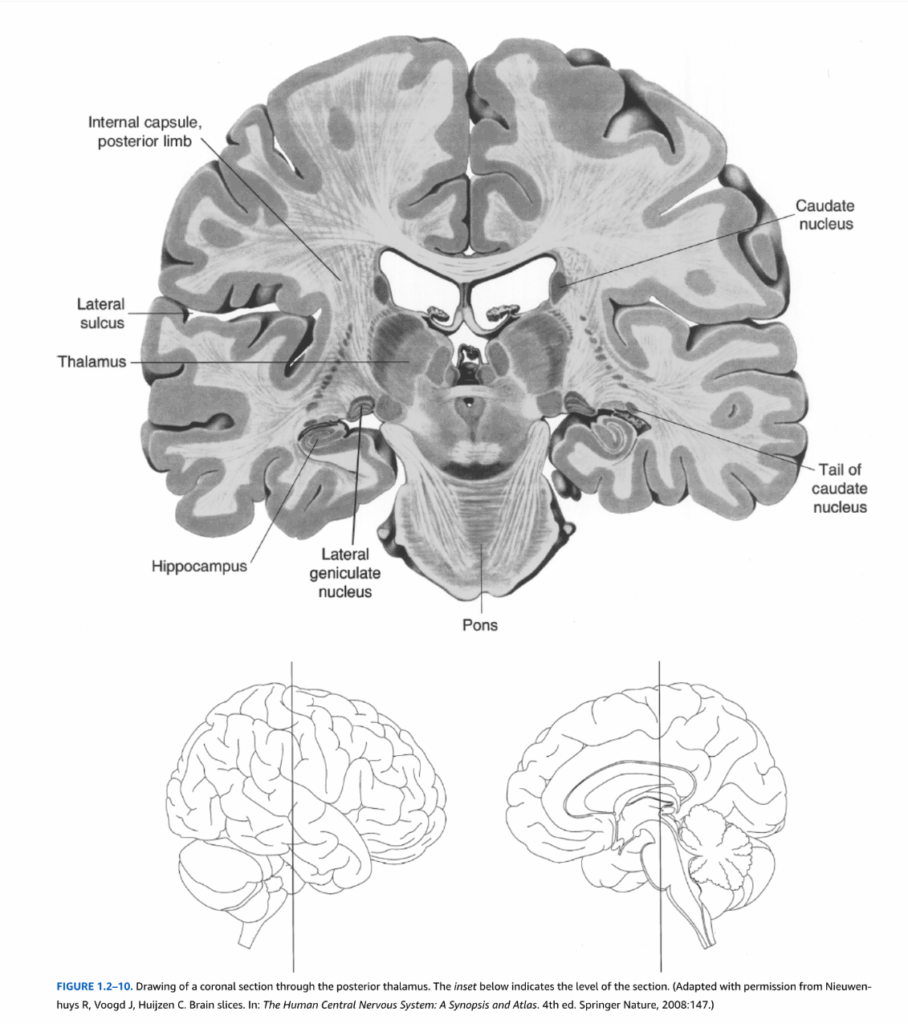
図1.2-10. 視床後部を通る冠状断面図。下の挿入図は断面のレベルを示している。(Nieuwenhuys R, Voogd J, Huijzen C. Brain slices. In: The Human Central Nervous System: A Synopsis and Atlas. 4th ed. Springer Nature, 2008:147より許可を得て改変)
連合線維 連合線維は同一半球内の皮質領域を結び、同一葉内の領域を結ぶ非常に短い線維から、異なる葉の領域を結ぶより長い線維まで、その大きさは様々である。短連合線維は隣接する脳回を結び、ある脳回から別の脳回へとUの字を形成することからU線維と呼ばれることが多い(図1.2-14)。同一半球内の遠隔皮質領域を結ぶ長連合線維の主要な線維束は5つある。上縦束は島の上方で半球の外側に位置し、前頭葉、頭頂葉、後頭葉皮質を結んでいる。弓状束は前頭葉と側頭葉を相互に結んでいる。鉤状束は湾曲した線維束で、前頭葉の眼窩部と側頭葉前部を結んでいる。その名の通り、下後頭前頭束は後頭葉と前頭葉を結び、半球内を腹側かつ外側に走行する線維束である。帯状束は帯状回の下の白質内にあり、この脳回を海馬傍回と結んでいる。下縦束は側頭葉と後頭葉を結んでいる。これらの線維束は皮質領域間の独立した点対点の経路ではなく、その経路全体にわたって線維が出入りする連続的な経路である。その他の連合線維束には、前障と被殻の間に挟まれた外包、および前障と島皮質の間にある最外包がある(図1.2-8参照)。
半球内および半球間の結合性の障害は、統合失調症の病態生理に関与していると考えられている。例えば、統合失調症患者のMRI研究では、脳梁、内包、前交連の白質密度が低いことが明らかになっている。さらに、組織の構造と微細構造に関する情報を提供するDTI(拡散テンソル画像)を用いた研究では、統合失調症患者において脳梁、内包、帯状束、鉤状束、脳弓を含むほぼすべての主要白質路に異常が認められ、また抑うつ患者では脳梁と鉤状束の微細構造の変化が報告されている。白質路の異常は他の神経精神疾患でも報告されている。例えば、MRI研究では、アルツハイマー病患者や自閉症児において脳梁の横断面積の減少が明らかになっている。
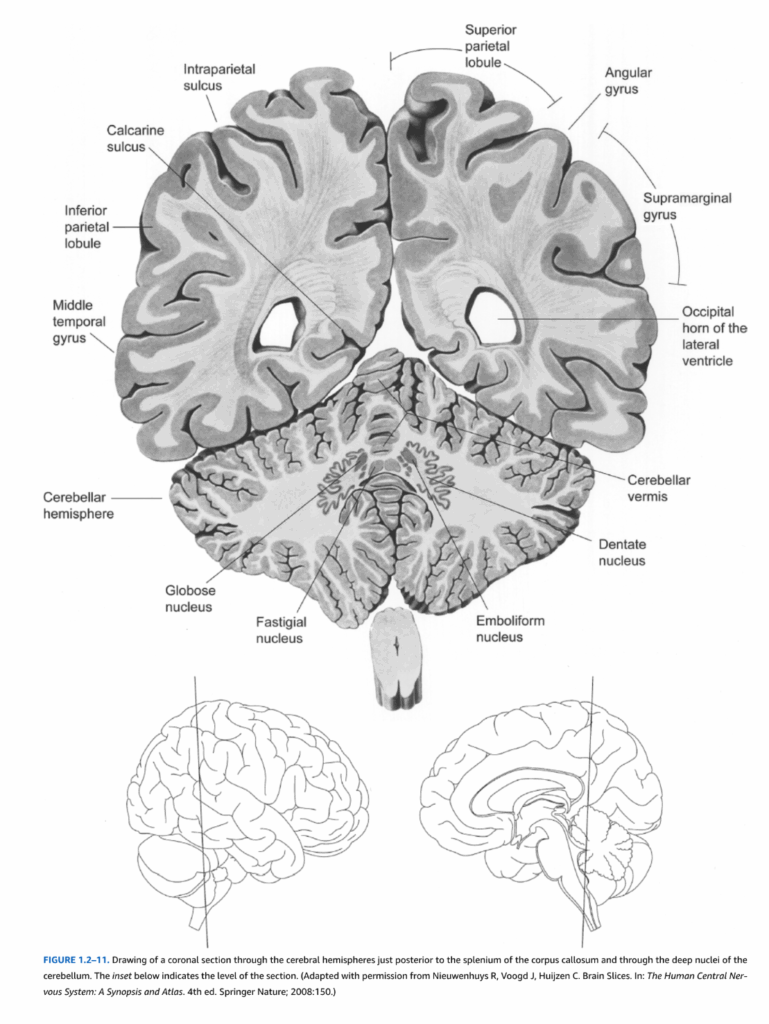
図1.2-11. 脳梁膨大部の直後および小脳深部核を通る大脳半球の冠状断面図。下の挿入図は断面のレベルを示している。(Nieuwenhuys R, Voogd J, Huijzen C. Brain Slices. In: The Human Central Nervous System: A Synopsis and Atlas. 4th ed. Springer Nature; 2008:150より許可を得て改変)
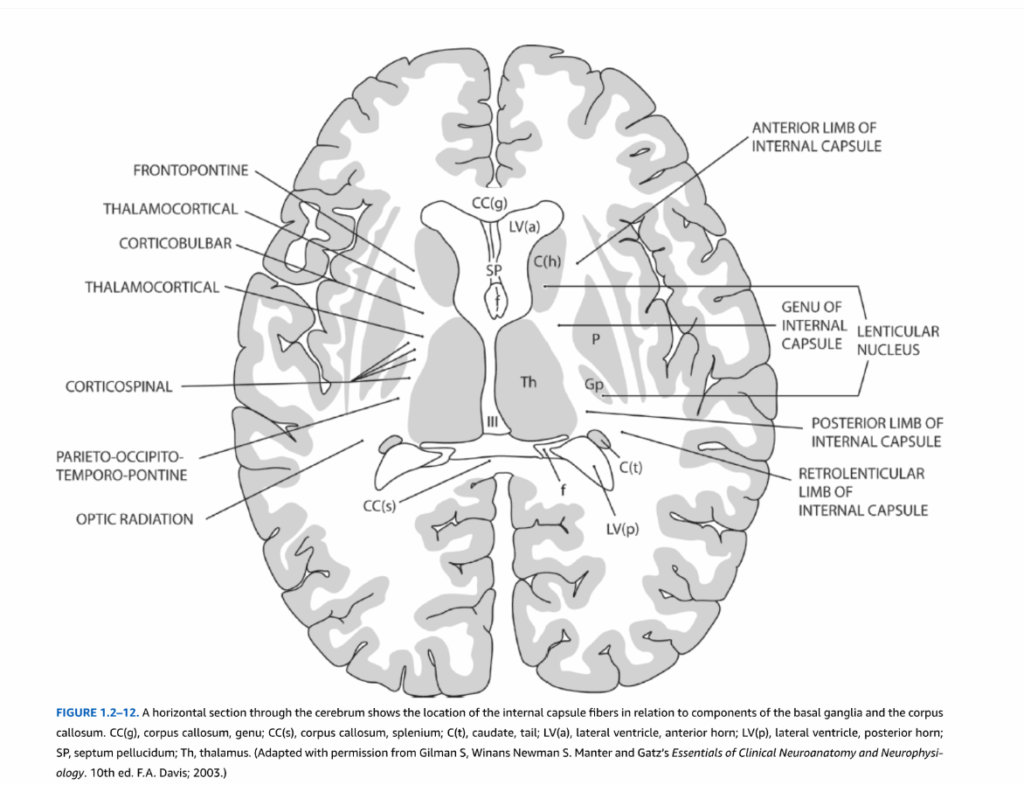
図1.2-12. 大脳の水平断面は、基底核および脳梁の構成要素との関係における内包線維の位置を示している。CC(g)、脳梁膝部;CC(s)、脳梁膨大部;C(t)、尾状核尾部;LV(a)、側脳室前角;LV(p)、側脳室後角;SP、透明中隔;Th、視床。(Gilman S, Winans Newman S. Manter and Gatz’s Essentials of Clinical Neuroanatomy and Neurophysiology. 10th ed. F.A. Davis; 2003より許可を得て改変)
脳室系
発生過程で神経管が癒合すると、神経管の空洞は脳の脳室系となる。脳室系は大脳半球内の2つのC字型の側脳室から構成され、さらに5つの部分に分けることができる:前角(前頭葉に位置)、脳室体部、下角または側頭角(側頭葉内)、後角または後頭角(後頭葉内)、そして心房である(図1.2-15)。モンロー孔(脳室間孔)は2つの側脳室を間脳の正中線上にある第三脳室と結ぶ2つの開口部である。中脳水道は第三脳室を橋および延髄の第四脳室と結んでいる。
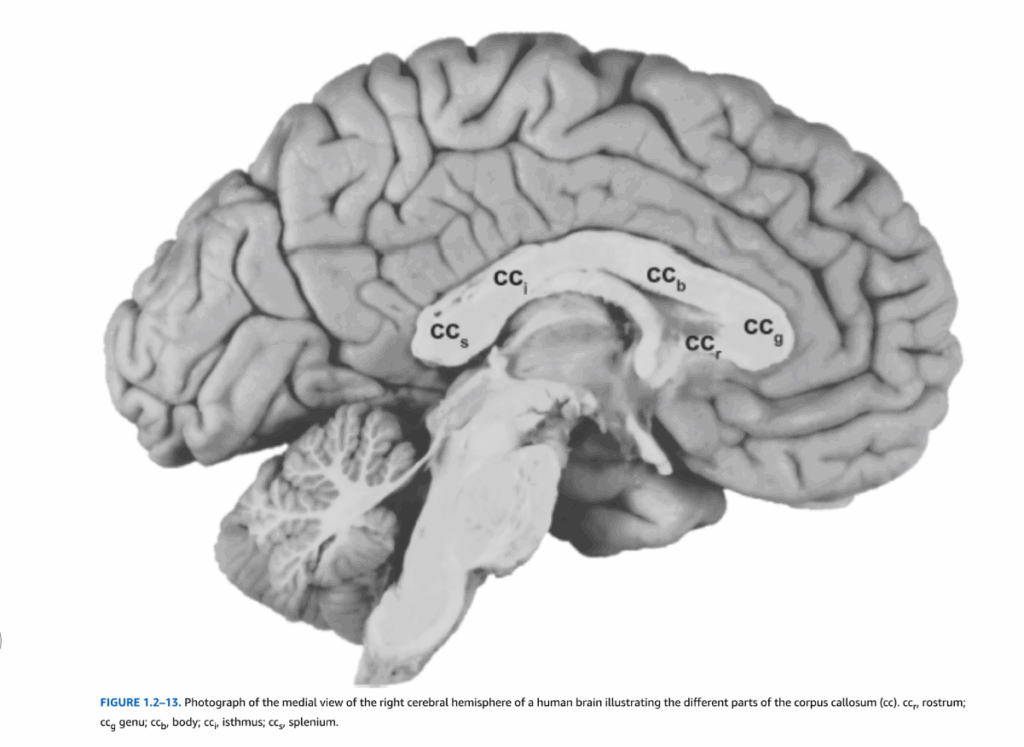
図1.2-13. 脳梁(cc)の異なる部分を示すヒト脳の右大脳半球内側面の写真。ccp、嘴部;ccg、膝部;ccb、体部;ccj、峡部;ccs、膨大部。
脳室系は脳脊髄液(CSF)で満たされている。CSFは無色の液体で、タンパク質、グルコース、カリウムの濃度は低く、ナトリウムと塩化物の濃度は比較的高い。CSFの大部分(70%)は、側脳室壁および第三脳室と第四脳室の天井に位置する脈絡叢で産生される。脈絡叢は上衣、軟膜、毛細血管の複合体で、脳室内に陥入している。脳の他の部分とは対照的に、脈絡叢の毛細血管は有窓性であり、物質が毛細血管から出て軟膜を通過することを可能にしている。しかし、上衣細胞または脈絡上皮細胞は細胞間に密着結合を有し、物質のCSFへの漏出を防いでいる。これは血液-CSF関門と呼ばれることもある。脳の他の部分では、毛細血管の内皮細胞が密着結合を示し、血液から脳への物質の移動を防いでいる。これは血液-脳関門と呼ばれる。
CSFは絶えず産生され、側脳室から第三脳室、そして第四脳室へと循環する。その後CSFは正中孔と外側孔を通って大槽と橋槽に流れ、最終的に大脳半球上を移動してくも膜顆粒に吸収され、上矢状静脈洞に放出される。CSFの流れの障害は通常何らかの形の水頭症を引き起こす。例えば、脳室間孔が閉塞すると、関連する側脳室は拡大するが、脳室系の残りの構成要素は正常のままである。
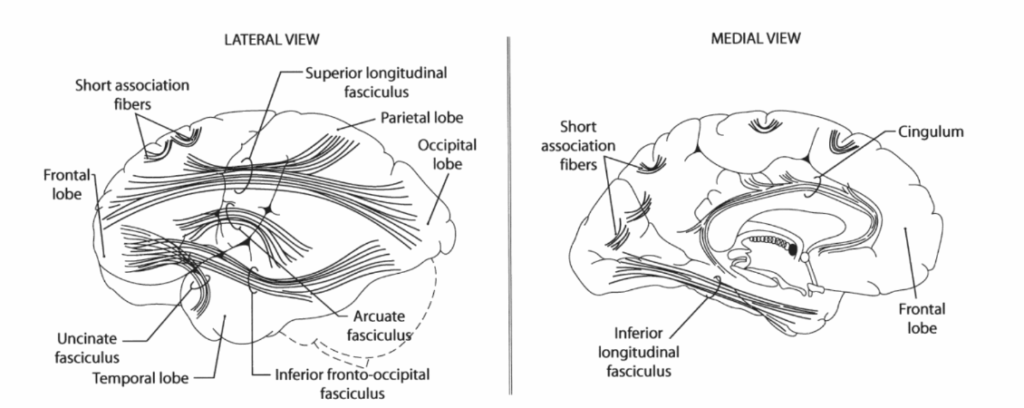
図1.2-14. 左半球の外側面(左パネル)と内側面(右パネル)から見た主要な連合線維路を示す図。(Haines DE. Fundamental Neuroscience for Basic and Clinical Applications. 3rd ed. Churchill Livingstone; 2006:253より改変)
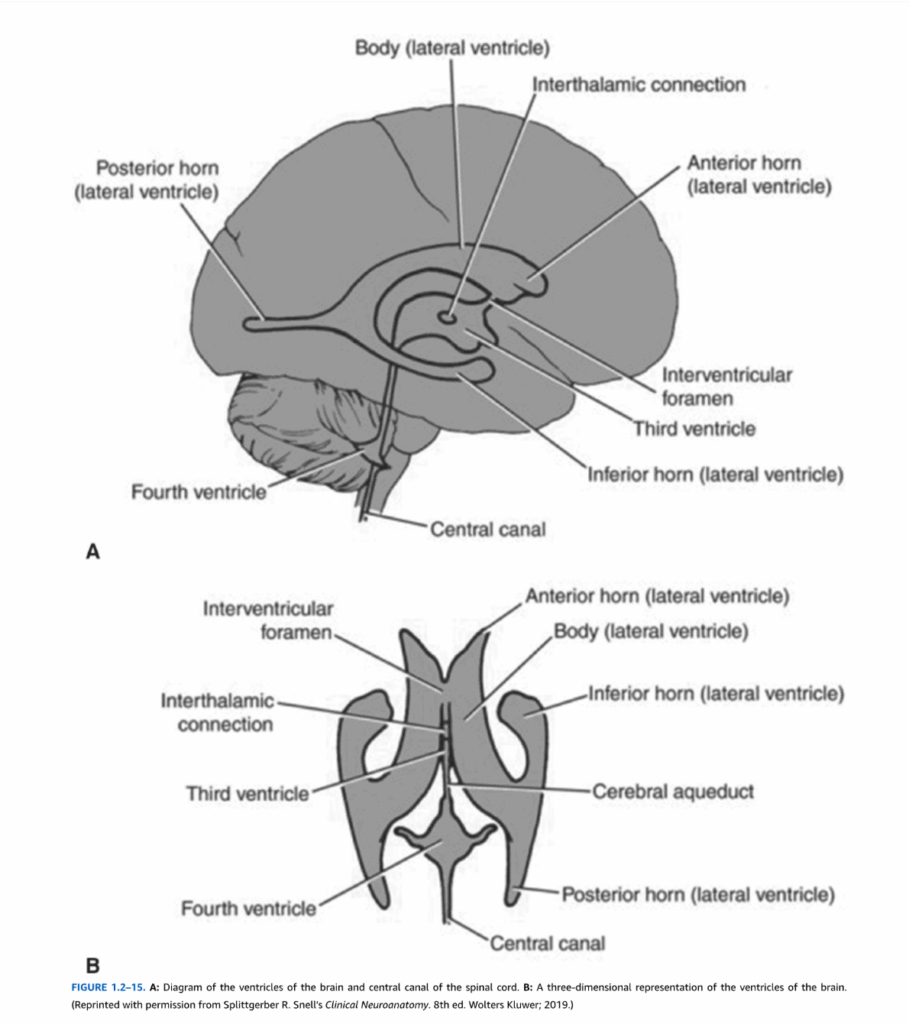
図1.2-15. A:脳の脳室と脊髄中心管の図。B:脳の脳室の三次元表示。(Splittgerber R. Snell’s Clinical Neuroanatomy. 8th ed. Wolters Kluwer; 2019より許可を得て転載)
CSFにはいくつかの機能がある:外傷に対する脳のクッション、細胞外環境の維持と制御、内分泌ホルモンの拡散である。CSFは脳を浸し、細胞外液と直接交通しているため、CSF中の特定の化合物を測定することで、脳内のその物質の量の相関を調べることが可能である。例えば、CSF中のアミロイドベータ1-42と総タウのレベルは、それぞれアミロイド代謝の変化と軸索損傷のマーカーと考えられている。腰椎穿刺で採取したCSFサンプル中のアミロイドベータ1-42と総タウの濃度は、軽度認知障害からアルツハイマー病への進行を調べる研究で使用されている。さらに、CSF中のアミロイドベータ1-42と総タウの測定値は、軽度認知障害患者におけるアルツハイマー認知症の確立された予測バイオマーカーである。
機能的脳システム
ヒト脳の組織原理と構造的構成要素との関係は、視床皮質系、基底核系、辺縁系の3つの機能的システムで説明される。
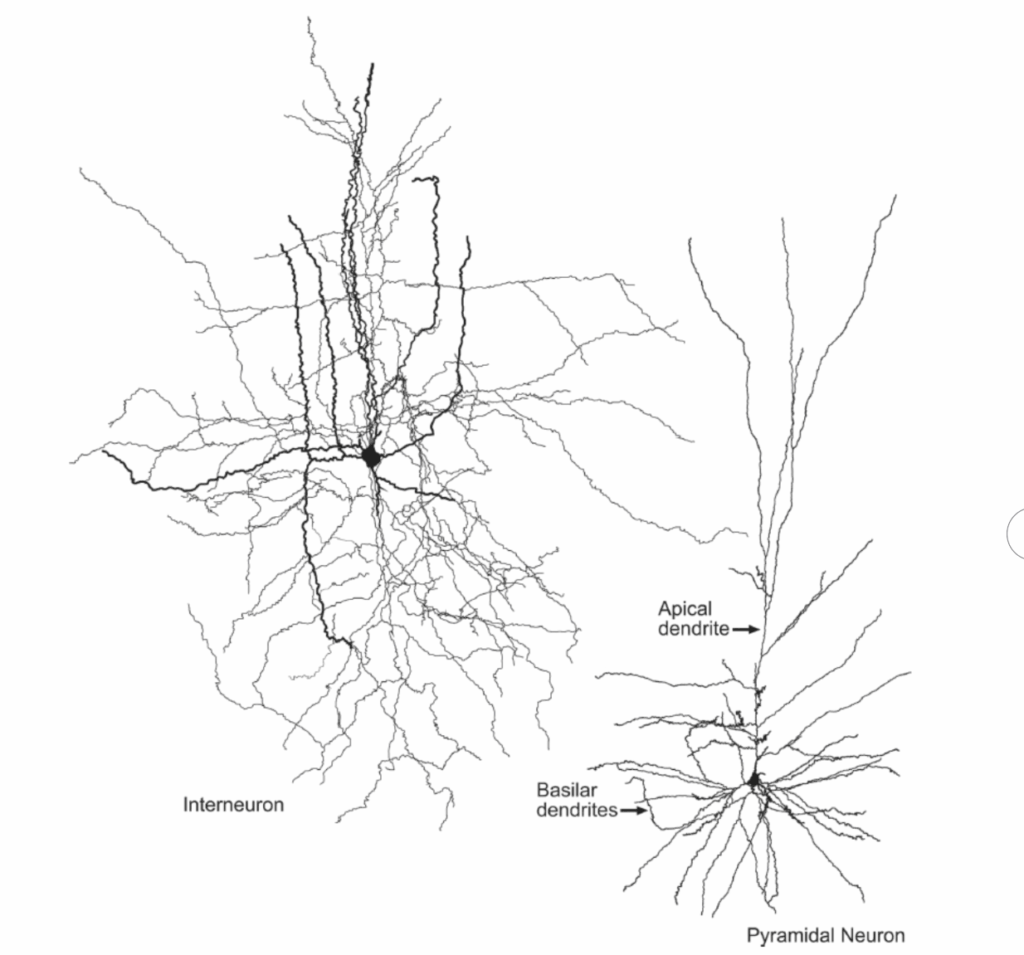
図1.2-16. 介在ニューロン(左)と錐体ニューロン(右)の図。これら2つのタイプのニューロンの形態の違いに注目。介在ニューロンの細胞体は円形または卵円形である傾向があるが、錐体ニューロンの細胞体は二次元的視点から一般的に三角形に見える。また、2つの細胞間における樹状突起と軸索樹状分岐の違いにも注目。介在ニューロンから生じる突起は多方向に分岐するように見えるが、錐体ニューロンは明瞭でよく定義された頂端および基底樹状突起を有している。(Permission from: Neuromorpho.org; Ascoli GA, Donohue DE, Halavi M. NeuroMorpho.Org: a central resource for neuronal morphologies. J Neurosci. 2007;27(35):9247-9251(NM)-01861; Krimer LS, Zaitsev AV, Czarner G, et al. Cluster analysis-based physiological classification and morphological properties of inhibitory neurons in layer 2-3 of monkey dorsolateral prefrontal cortex. J Neurophysiol. 2005;94(5): 3009-3022.)
大脳皮質
大脳皮質は大脳半球を覆う数ミリメートルの厚さの層状ニューロンシートである。約220億個のニューロンが約165兆個のシナプスを介して交流している。これらのニューロンは約1,200万キロメートルの樹状突起を有し、大脳皮質と皮質下領域は約10万キロメートルの軸索によって相互に結ばれている。全皮質面積の90%以上は新皮質から構成され、6層構造を有している(少なくとも発生過程のある時点で)。大脳皮質の残りは異皮質(他の層構造を有する皮質領域)と呼ばれ、古皮質と原皮質から構成され、それぞれ終脳の基底部と海馬体に限られている。
新皮質内では、主要な2つのニューロン細胞型は錐体ニューロンと星状または非錐体ニューロンである(図1.2-16)。新皮質ニューロンの約75%を占める錐体ニューロンは、通常、皮質表面に向かって垂直に上昇する単一の頂端樹状突起を生じる特徴的な形の細胞体を有している。さらに、これらのニューロンは細胞の基底部から外側に広がる一連の短い樹状突起を有している。錐体ニューロンの樹状突起は樹状突起棘と呼ばれる短い突起で覆われており、これらのニューロンへの興奮性シナプスの大部分の部位となっている(図1.2-17)。ほとんどの錐体細胞は、グルタミン酸を神経伝達物質として使用する興奮性投射ニューロンである。興味深いことに、死後研究では、統合失調症患者は背外側前頭前野の第3層深部の錐体ニューロン基底部に位置する樹状突起の棘が少ないように見える。この樹状突起棘の欠損は、アクチン動態の調節における遺伝的脆弱性によるものかもしれず、構造的不安定性および/または棘の過剰なミクログリア介在性刈り込みによる樹状突起棘の喪失につながる。棘の減少、そしておそらくそれらが受ける興奮性シナプスの減少は、錐体ニューロンの興奮の減少をもたらし、最終的にこれらの錐体ニューロンの出力の減少、そして最終的に皮質活動指数の低下につながる。
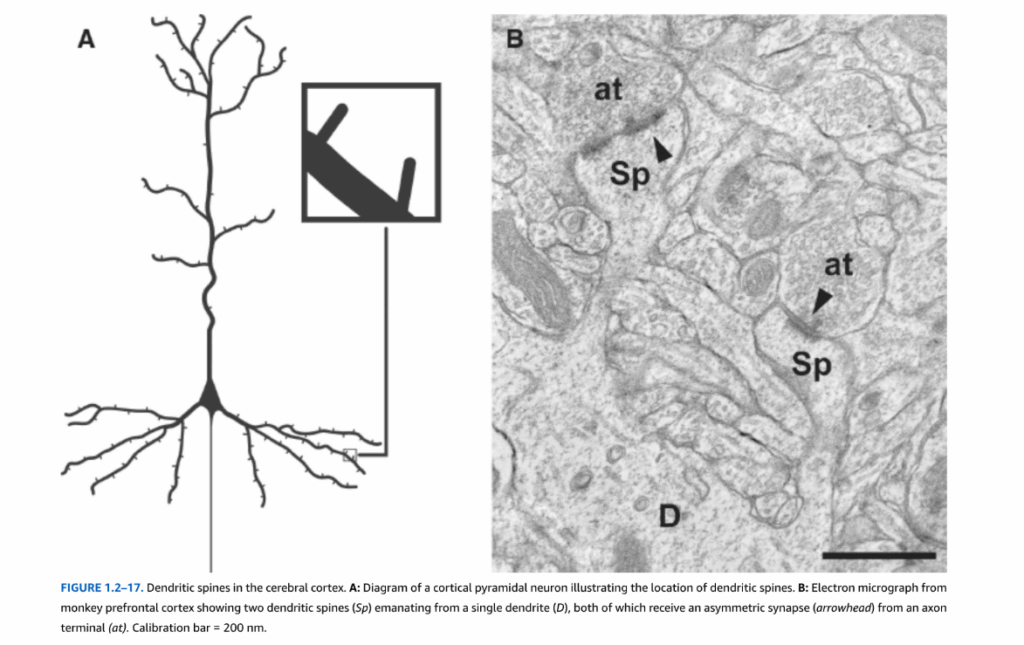
図1.2-17. 大脳皮質の樹状突起棘。A:樹状突起棘の位置を示す皮質錐体ニューロンの図。B:単一の樹状突起(D)から出現する2つの樹状突起棘(Sp)を示すサル前頭前野の電子顕微鏡写真。両方とも軸索終末(at)から非対称シナプス(矢頭)を受けている。較正バー = 200 nm。
対照的に、非錐体細胞は一般的に小さな局所回路ニューロンで、その多くは抑制性神経伝達物質GABAを使用している(図1.2-16)。介在ニューロンとしても知られる皮質GABA細胞の軸索は灰白質内で樹状分岐し、ほとんどは存在する皮質領域から外に投射しない。大脳皮質には複数の異なるGABAニューロンのサブタイプが存在し、これらは生化学的、電気生理学的、形態学的に区別できる。例えば、GABA細胞の亜集団は特定の神経ペプチドやカルシウム結合タンパク質の存在によって区別できる。さらに、軸索樹状分岐の構成と軸索終末のシナプス標的は、これらの異なるサブタイプ間で大きく異なる。
GABAシャンデリア細胞はカルシウム結合タンパク質パルブアルブミン(PV)を発現し、カートリッジと呼ばれる明確な垂直構造として配列された軸索終末を有している(図1.2-18)。これらの軸索終末は、錐体細胞の軸索起始部のみと抑制性または対称シナプスを形成する。PV含有バスケット細胞は錐体ニューロンの細胞体と樹状突起に対称シナプスを形成する。PV含有ニューロンは主に第3層と第4層に位置している。
対照的に、GABAマルティノッティ細胞は神経ペプチドソマトスタチンを含有し、錐体ニューロンの房状樹状突起に対称シナプスを形成する。一部のダブルブーケGABAニューロンは放射状に配向した軸索樹状分岐を有し、ソマトスタチンとカルシウム結合タンパク質カルビンディンを含有し、錐体ニューロンの遠位樹状突起幹と棘に対称シナプスを形成する。神経ペプチドコレシストキニン(CCK)は、脳カンナビノイド受容体1(CB1)も発現する別のバスケット細胞集団に存在する。これらのCCK/CB1バスケット細胞は主に錐体ニューロンの樹状突起幹と細胞体にシナプスを形成する。対照的に、カルレチニン含有ダブルブーケ細胞は主に他のGABAニューロンの樹状突起幹に対称シナプスを形成する。カルレチニン含有カハール・レチウス細胞は第1層のみに存在し、錐体ニューロンの房状樹状突起を標的とする。
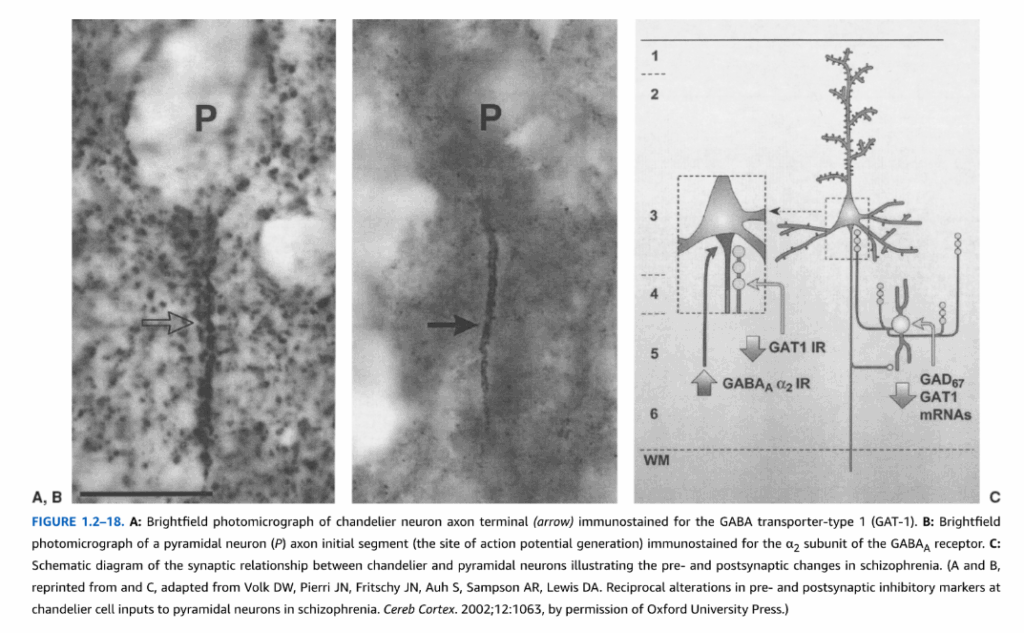
図1.2-18. A:GABA輸送体タイプ1(GAT-1)に対して免疫染色されたシャンデリアニューロン軸索終末(矢印)の明視野顕微鏡写真。B:GABAA受容体のα2サブユニットに対して免疫染色された錐体ニューロン(P)軸索起始部(活動電位発生部位)の明視野顕微鏡写真。C:統合失調症におけるシナプス前およびシナプス後の変化を示すシャンデリア細胞と錐体ニューロン間のシナプス関係の模式図。(AとBは転載、Cは改変:Volk DW, Pierri JN, Fritschy JM, Auh S, Sampson AR, Lewis DA. Reciprocal alterations in pre- and postsynaptic inhibitory markers at chandelier cell inputs to pyramidal neurons in schizophrenia. Cereb Cortex. 2002;12:1063, Oxford University Pressの許可による)
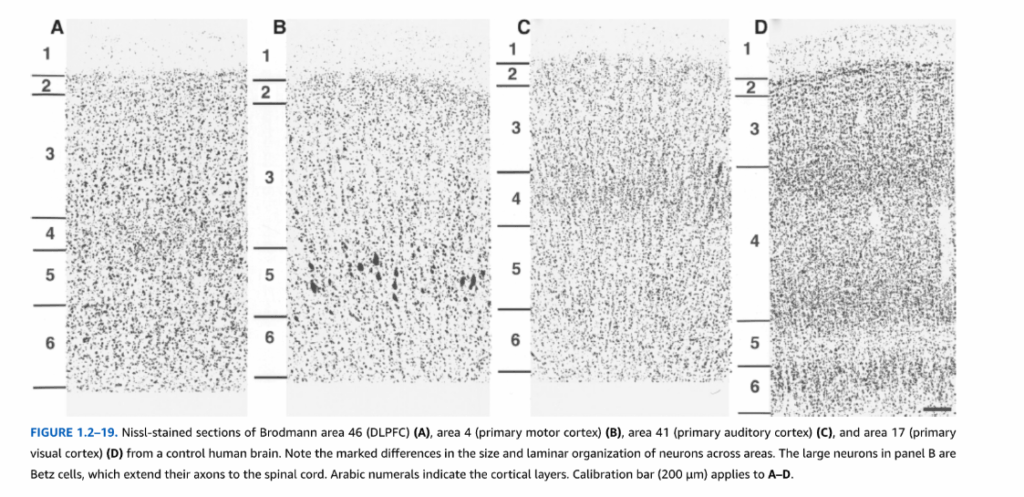
図1.2-19. ヒト対照脳からのブロードマン野46(DLPFC)(A)、野4(一次運動野)(B)、野41(一次聴覚野)(C)、野17(一次視覚野)(D)のニッスル染色切片。各領域におけるニューロンのサイズと層構成の顕著な違いに注目。パネルBの大型ニューロンはベッツ細胞で、軸索を脊髄まで伸ばしている。アラビア数字は皮質層を示す。較正バー(200 μm)はA-Dに適用。
多くの研究により、様々な神経疾患および精神疾患において異なる非錐体ニューロン集団の異常が明らかになっている。例えば、てんかん組織ではカルレチニンニューロンの数の減少とPVニューロンの軸索樹状分岐の形態学的変化が報告されている。統合失調症では、バスケット細胞はPVと生合成酵素グルタミン酸脱炭酸酵素の67キロダルトン型(GAD67)のレベルが低い。シャンデリアニューロンの軸索終末では膜GABA輸送体タイプ1(GAT-1)のレベルが減少し、シナプス後錐体ニューロンではGABAA α2受容体のレベルが増加している(図1.2-18C参照)。これらの所見は総合的に前頭前野皮質回路の機能不全を反映しており、統合失調症で観察される認知症状に寄与している可能性がある。
新皮質ニューロンは新皮質の6層にわたって分布している。これらの層は構成ニューロンの相対的サイズと密度によって区別される(図1.2-19)。各皮質層は特定のタイプの入力を受け、特徴的な投射を提供する傾向がある。例えば、視床中継核からの求心線維は主に第3層深部と第4層に終止し、他の皮質領域からの求心線維は主に第1-3層に終止する。対照的に、皮質線条体投射は第5層錐体細胞から生じ、皮質視床投射は主に第6層錐体ニューロンから発生する(図1.2-20)。これらの層の区別は、精神疾患における病態生理学的機序の可能性を解明する重要な手がかりを提供する。統合失調症患者の前頭前野第3層深部錐体ニューロンの細胞体サイズの縮小と棘密度の低下の報告は、これらの変化が前頭前野への主要な視床投射核である内側背側視床核からの求心投射の変化と関連している可能性を示唆している。
水平な層構造に加えて、皮質構成の多くの側面は垂直または円柱状の特徴を有している。例えば、錐体ニューロンの頂端樹状突起と一部の局所回路ニューロンの軸索は顕著な垂直配向を有しており、これらの神経要素がそれぞれ複数の層への入力をサンプリングしたり、複数の層のニューロンの機能を調節したりする可能性を示している。他の皮質領域から新皮質への求心入力も、円柱様式で皮質層にわたって分布する傾向がある。最後に、体性感覚野と視覚野での生理学的研究により、特定の円柱内のニューロンは特定の特徴を持つ刺激に応答するが、隣接する円柱のニューロンは異なる特徴を持つ刺激に応答することが示されている。
感覚野で最もよく研究されているが、この組織パターンは連合野にも存在する。サルでの経路追跡技術を用いたより最近の研究により、前頭前野皮質ニューロンのクラスターが相互に結合した離散的なモジュラーストライプに組織化されており、これが視覚野で同定された円柱の類似体であることが示されている(図1.2-4参照)。この組織化は前頭前野のワーキングメモリーと実行機能に寄与している可能性があると仮説が立てられている。
新皮質は2つの一般的なタイプの領域に分けることができる。容易に識別可能な6層構造を有する領域は同型皮質として知られ、前頭葉、側頭葉、頭頂葉の連合領域に見られる。対照的に、新皮質の一部の領域は6層構造を有していない。異型皮質と呼ばれるこれらの領域には、明確な第4層を欠く一次運動野(図1.2-19B参照)と、拡張した第5層を示す一次感覚領域(図1.2-19D参照)が含まれる。新皮質はさらに個別の領域に分けることができ、各領域は独特の構築、特定の結合セット、特定の脳機能における役割を有している。ヒト新皮質の細分化の大部分は細胞構築学的特徴に基づいている。すなわち、細分化は層間でのニューロンのサイズ、密度、配列が異なる(図1.2-19参照)。最も広く使用されているシステムはコルビニアン・ブロードマンのものであり(図1.2-21)、彼は各半球の皮質を44の番号付き領域に分割した。これらの番号付き領域の一部は、野4(中心前回の一次運動野)や野17(後頭葉の一次視覚野)など、機能的に明確な領域と密接に対応している。対照的に、他のブロードマン野は機能的属性が異なるいくつかの皮質帯域を包含しているように見える。ブロードマンの脳地図は精神疾患の死後研究で広範に使用されているが、領域間の区別の多くは微妙であり、領域間の境界の位置は個体間で変動する。
