
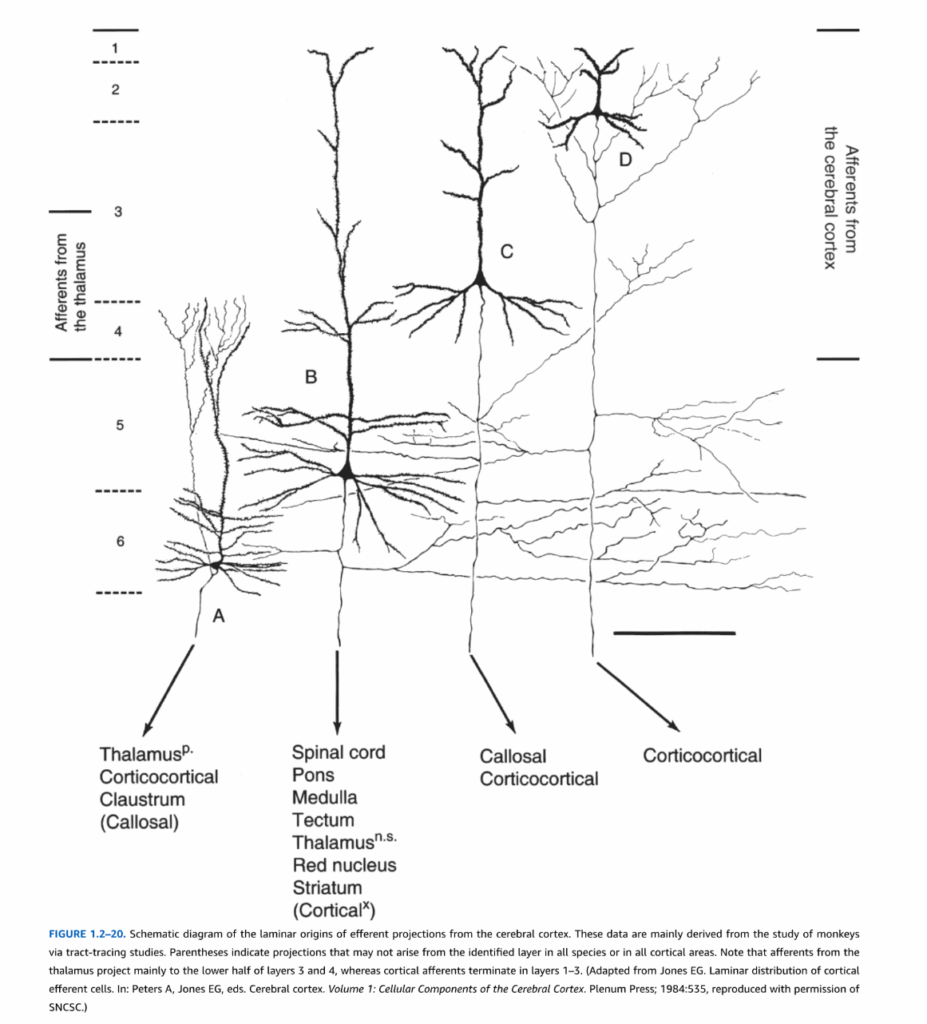
図1.2-20. 大脳皮質からの遠心性投射の層起源の模式図。これらのデータは主に経路追跡研究によるサルの研究から得られたものである。括弧は、すべての種またはすべての皮質領域において同定された層から生じない可能性のある投射を示す。視床からの求心線維は主に第3層と第4層の下半分に投射するが、皮質求心線維は第1-3層に終止することに注目。(Jones EG. Laminar distribution of cortical efferent cells. In: Peters A, Jones EG, eds. Cerebral cortex. Volume 1: Cellular Components of the Cerebral Cortex. Plenum Press; 1984:535より改変、SNCScの許可を得て転載)
特定の皮質領域は他の入力を受ける可能性があるが、特定の視床核からの投射と、同一半球内(連合線維)または対側半球(交連線維)の特定の他の皮質領域からの投射により強く神経支配されている。結合パターンにより、皮質領域を異なるタイプに分類することが可能である。一次感覚領域は特定の視床中継核からの入力が優勢で、それぞれ一次視覚野、一次体性感覚野、一次聴覚野の皮質表面における視覚空間、体表面、または可聴周波数範囲の地形表現によって特徴づけられる。これらの領域は近隣の単一様式連合領域に投射し、これらの領域も特定の感覚様式からの情報処理に専念している。これらの領域からの出力は、前頭前野や側頭頭頂皮質領域などの多様式連合領域で収束する。これらの領域のニューロンは複雑な刺激に応答し、高次認知機能の仲介者と考えられている。最後に、これらの領域は行動反応を制御する大脳皮質の運動領域の活動に影響を与える。
この皮質領域の分類スキームは多くの点で正確であるが、皮質情報処理の既知の複雑さの一部を説明できない。例えば、視床からの体性感覚入力は大脳皮質のいくつかの異なる地形的に組織化された地図に投射する。さらに、皮質内の情報の流れは分類スキームで示唆される直列処理経路に限定されず、視床から一次および連合領域への感覚入力などの並列処理ストリームも含む。
この議論では大脳半球間を区別していないが、言語などの特定の脳機能は一方の半球に局在している。機能の偏側性の構造的基盤は決定されていないが、大脳半球間でいくつかの解剖学的違いが観察されている。例えば、側頭平面と呼ばれる上側頭皮質の一部は、一般的に左半球の方が右半球より大きい。一次聴覚野に近く、ウェルニッケ野として知られる領域を含むその皮質領域(図1.2-5参照)は、左半球に局在する受容性言語機能に関与しているようである。さらに、左下前頭皮質のブロードマン野44(ブローカ野)(図1.2-5参照)は右半球の同所的領域よりも大きな錐体ニューロンを含んでおり、この違いが運動性言語機能に対するブローカ野の専門化に寄与している可能性がある。ブローカ野の病変は途切れた発話を引き起こし、ウェルニッケ野の病変は意味をなさない多弁な発話を引き起こす。
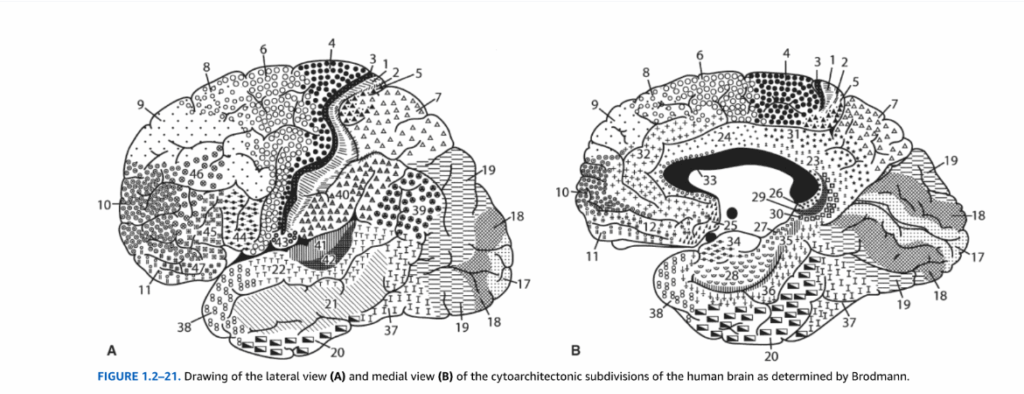
図1.2-21. ブロードマンによって決定された人間の脳の細胞構築学的区分の外側面図(A)と内側面図(B)の描画。
ブロードマンの脳地図は、大脳皮質を細胞の組織構造に基づいて番号付きの領域に分割した古典的な神経解剖学的参考資料です。1から52までの番号が付けられた領域(すべての番号が使用されているわけではありません)は、人間の皮質を区分する最も永続的で広く使用されているシステムの一つです。
よく知られている領域には以下があります:
- 領域3、1、2:一次体性感覚野
- 領域4:一次運動野
- 領域17:一次視覚野
- 領域41、42:一次聴覚野
- 領域44、45:ブローカ野(優位半球)
- 領域22:ウェルニッケ野の一部
外側面図では通常、運動制御、体性感覚、言語処理、高次連合機能に関わる領域が示され、内側面図では辺縁系機能、視覚処理、運動制御に関連する領域が表示されます。
機能的回路
皮質皮質結合 皮質皮質結合は大脳皮質内の結合の大部分を占め、3つの基本カテゴリーに分けることができる:(1)内在性-一つの皮質領域内の結合、(2)連合性-同一半球内の皮質領域間の結合、(3)脳梁性-異なる半球の皮質領域間の結合。内在性結合は錐体ニューロンの軸索側枝によって形成され、連合性および脳梁性結合は錐体ニューロンの主軸索によって形成される。
皮質皮質結合は、皮質領域内および皮質領域間に分布する大きなニューロン群の協調活動に責任を負っている。この活動の同期化は、大脳皮質に帰属する知覚および認知機能の大部分に関与しているようである。
これらの結合されたニューロン群の同期神経活動または振動は、脳波(EEG)で記録できる特徴的な電気活動パターンを生成する。特定の周波数で一緒に振動するニューロンの数が多いほど、ノイズを超えるEEG信号の振幅が大きくなる。これらの振動は、シータ(4-8 Hz)、アルファ(8-12 Hz)、ガンマ(30-80 Hz)などの異なる周波数帯域にグループ化される。すべての周波数帯域の振動活動は、抑制性GABAニューロンのネットワークから生じ、その出力が錐体ニューロンの活動を調節する。
高周波振動は、知覚、注意、認知などの様々な精神過程に関与する分散ニューロンネットワークの統合のための可能なメカニズムとして提案されている。DLPFCにおけるガンマ帯域振動は認知の重要な神経基盤と考えられている。さらに、DLPFCの顆粒上層内でのPVバスケットサブクラスのGABAニューロンと錐体ニューロン間のシナプス結合がこの振動活動を生成するようである。興味深いことに、統合失調症患者は認知課題遂行時に前頭前野ガンマ振動のパワーが低く、これはおそらく統合失調症患者の死後DLPFCで観察されるPVニューロンを含むGABA系の異常の結果である。
皮質皮質回路のネットワーク 皮質皮質結合の研究は、かつては経路追跡法を用いた動物でのみ可能であった。これらの方法の改良により、異なる皮質領域間の投射に関する知識が洗練され深化したが、この方法論で明らかになった結合がヒトでも同じであると推測することしかできない。過去20年間で、神経画像研究により、ヒトの皮質皮質結合性に関する知識が大幅に拡大した。しかし、これらの神経画像研究の大部分は機能的結合性(すなわち、特定のタスク中に共活性化する脳領域)を同定しており、領域間の物理的結合ではないことに注意することが重要である。ヒト脳における直接的な解剖学的皮質皮質結合の証拠は乏しいが、DTIなどの新しい画像技術は構造的詳細を提供する。注意や記憶などの特定の機能に寄与する分散領域を結ぶ大規模皮質ネットワークは長い間提案されており、現在は前述の神経画像技術でより容易に研究されている。数多くのネットワークが記述されているが、3つの特定のネットワークが最も注目を集めている。
表1.2-2. 大脳皮質の機能的ネットワーク
| ネットワーク | 関与する脳領域 | 機能 |
|---|---|---|
| 中央実行系 | 外側前頭前野背側部 | 注意 |
| 外側後頭頂皮質 | ワーキングメモリ | |
| 意思決定 | ||
| 顕著性 | 前帯状皮質 | 関連する入力刺激の検出 |
| 腹内側前頭前野 | ||
| 島皮質 | ||
| デフォルト | 内側前頭前野 | 内的認知 |
| 後帯状皮質 | 心の理論 | |
| 内側後頭頂皮質 |
機能的ネットワークの詳細
中央実行系ネットワークは外側前頭前野背側部(DLPFC)と外側後頭頂皮質の複数領域を含み、注意、ワーキングメモリ、意思決定などの認知機能に関与しています。
顕著性ネットワークは前帯状皮質、腹内側前頭前野、島皮質で構成され、認知的、恒常性維持的、感情的な入力情報の重要度に応答します。島皮質は内受容感覚(体内の臓器や組織の生理学的状態の表現と統合)に主に関連していますが、最近の研究では身体認識、自己認識、感情認識、内受容感覚情報と感情の統合など、より広範な機能があることが示されています。
デフォルトモードネットワークは内側前頭前野、後帯状皮質、後頭頂皮質の内側領域で形成され、安静時や外的課題に従事していない時に活動します。このネットワークは個人的経験の想起、将来の想像、心の理論など、内的認知様式に関与しています。
ネットワーク間の相互作用
最近の研究では、これら3つのネットワークが相互作用し、顕著性ネットワークが課題陽性の中央実行系ネットワークと課題陰性のデフォルトモードネットワーク間のスイッチとして機能することが示されています。フォン・エコノモニューロンという人間特有の神経細胞が前島皮質にのみ存在し、この切り替え機能を担っていると考えられています。
精神疾患との関連
これらのネットワークの機能不全は様々な神経精神疾患に関連しています:
- うつ病:デフォルトモードネットワーク内の機能的結合が異常に高く、中央実行系ネットワーク間の結合が低下
- 統合失調症:3つのネットワーク間の機能的結合が整理されておらず、特に顕著性ネットワークの機能不全により、外界と内的思考・感情を区別する能力が破綻し、妄想や幻覚などの精神病症状が現れる
前障(クラウストルム)
前障は島皮質と最外包の間にある薄い灰白質で、脳全体に広範な結合を持ちます。感覚皮質領域と密に結合し、意識体験への情報統合や顕著性検出器として機能すると考えられています。セロトニン2A受容体の発現が脳内で最も高く、サイケデリック物質の作用機序との関連が注目されています。
視床の解剖学的構造
視床は間脳の最大部分で、大脳皮質への主要な中継基地として機能します。6つの核群に分類されます:前核群、内側核群、外側核群、網様体核群、髄板内核群、正中核群。これらの核群は内髄板と呼ばれるY字状の有髄線維によって区分されています。
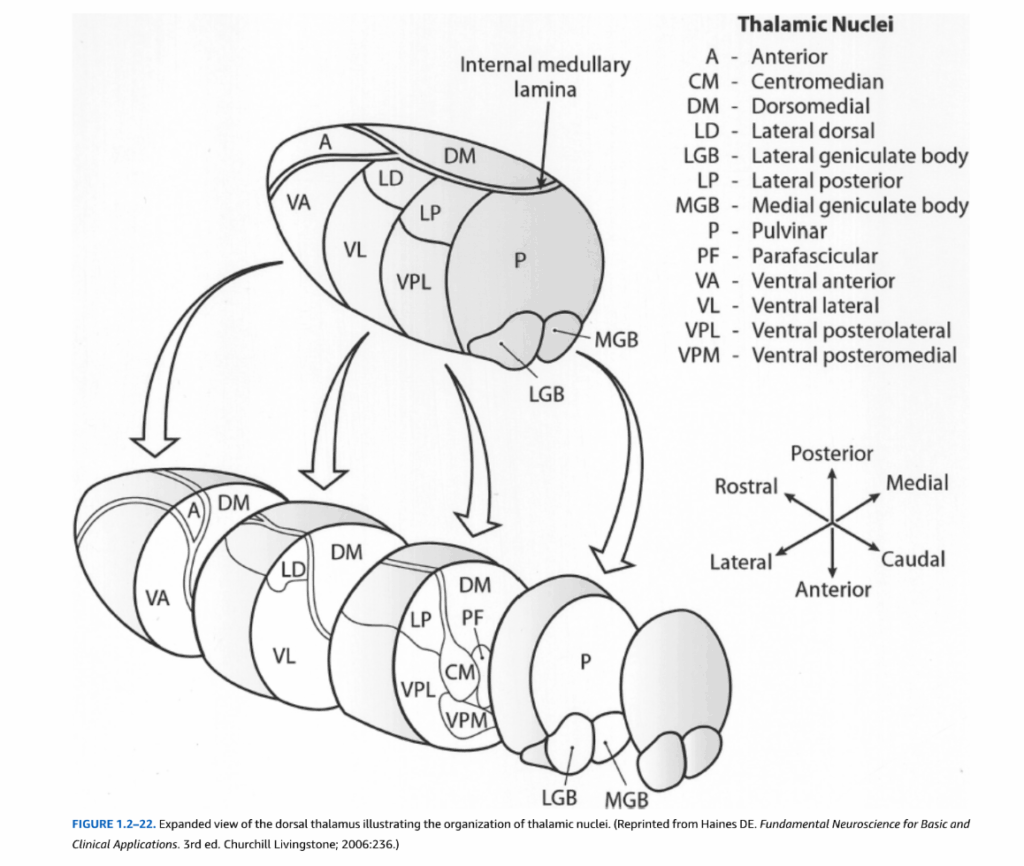
図1.2-22. 視床核の組織化を説明する背側視床の拡大図
(Haines DE. Fundamental Neuroscience for Basic and Clinical Applications. 3rd ed. Churchill Livingstone; 2006:236より転載)
視床核はまた、それらの結合のパターンと情報内容に基づいて複数のグループに分類することができる(表1.2-3)。中継核は大脳皮質の特定の領域に投射し、そこから入力を受ける。これらの相互結合は、明らかに大脳皮質がそれが受ける視床入力を調節することを可能にする。特異的中継核は単一の感覚モダリティからまたは運動系の明確な部分からの入力を処理する。例えば、外側膝状体核は視索から視覚入力を受け、後頭皮質の一次視覚野に投射する。図1.2-23に要約されているように、視床中継核のニューロンは大脳皮質の特定の領域に地形的に組織化された投射を提供するが、いくつかの皮質領域は複数の核から入力を受ける。
対照的に、連合中継核は複数のソースから高度に処理された入力を受け、連合皮質のより大きな領域に投射する。例えば、内側背側視床核は視床下部と扁桃体から入力を受け、前頭前野および特定の前運動皮質と側頭皮質領域と相互に相互結合している(図1.2-24)。中継核とは対照的に、びまん性投射核は多様なソースから入力を受け、大脳皮質の広範囲な領域と視床に投射する。これらの核の皮質結合の発散的な性質は、それらが皮質興奮性と覚醒のレベルを調節することに関与している可能性があることを示している。最後に、網様体核は、他の視床核と大脳皮質を相互に結合する軸索の側枝から入力を受ける抑制性ニューロンを含むという点で独特である。網様体核の各部分は、そこから入力を受ける視床核に投射する。結合のパターンは、網様体核が皮質求心性および遠心性活動をサンプリングし、その後その情報を使って視床機能を調節することを示している。
視床核を異なるカテゴリーに分類することは、視床回路についてのより大きな理解を反映するために過去20年間にわたって洗練されてきた。異なる核への分類は有用であるが、焦点は様々な核への入力のタイプに移ってきた。例えば、視床求心線維はドライバーまたはモジュレーターのいずれかとして定義することができる。ドライバーは感覚と末梢からの情報を伝達し、その後大脳皮質に中継される。これらの入力は視床中継の性質を決定する。例えば、外側膝状体核は網膜入力を一次視覚皮質に伝達するので視覚中継である。連合およびびまん性投射核に対するドライバー入力はより決定するのが困難であるが、これらの入力は最も可能性が高いのは大脳皮質の第5層錐体ニューロンから生じる。モジュレーターは視床への他のすべての入力を含み、脳幹核からのモノアミン作動性およびコリン作動性投射や大脳皮質の第6層錐体ニューロンからのフィードバック投射などである。視床回路はその後、ドライバー入力のソースに基づいて区別することができる。一次中継は上行性経路から生じるドライバーを持ち、まだ皮質で処理されていない情報を伝達する。対照的に、高次中継は第5層錐体ニューロンから生じるドライバーを持ち、視床を介して一つの皮質領域から別の皮質領域への情報の伝達を可能にする。したがって、与えられた視床核が一次と高次の両方の中継を含むことが可能である。興味深いことに、霊長類の視床核における回路の半分以上が高次と考えられており、皮質視床皮質機能における視床の複雑な役割を示唆している。
視床核の機能的分類と回路システム
視床核の機能的分類
視床核は結合パターンと情報内容に基づいて以下のグループに分類されます:
1. 中継核(Relay Nuclei)
- 特徴: 大脳皮質の特定領域に投射し、そこから入力を受ける相互結合
- 機能: 単一の感覚様式または運動系の特定部分からの入力を処理
- 例: 外側膝状体核
- 視索から視覚入力を受け取る
- 後頭皮質の一次視覚野に投射
- 地形的に組織化された投射を皮質特定領域に送る
2. 連合中継核(Association Relay Nuclei)
- 特徴: 複数のソースから高度に処理された入力を受ける
- 機能: より広範な連合皮質領域に投射
- 例: 内側背側視床核
- 視床下部と扁桃体から入力
- 前頭前野および特定の前運動皮質・側頭皮質領域と相互結合
3. びまん性投射核(Diffuse-Projection Nuclei)
- 特徴: 多様なソースから入力を受け、大脳皮質と視床の広範囲に投射
- 機能: 皮質興奮性と覚醒レベルの調節に関与
4. 網様体核(Reticular Nucleus)
- 特徴: 抑制性ニューロンを含む
- 機能:
- 他の視床核と大脳皮質を相互結合する軸索の側枝から入力
- 入力を受けた視床核に逆投射
- 皮質求心性・遠心性活動をサンプリングし、視床機能を調節
新しい分類システム:ドライバーとモジュレーター
ドライバー(Drivers)
- 定義: 感覚・末梢からの情報を伝達し、大脳皮質に中継
- 機能: 視床中継の性質を決定
- 例: 外側膝状体核では網膜入力がドライバー
- 連合核・びまん性投射核: 大脳皮質第5層錐体ニューロンがドライバーの可能性
モジュレーター(Modulators)
- 定義: その他すべての視床への入力
- 含まれるもの:
- 脳幹核からのモノアミン作動性・コリン作動性投射
- 大脳皮質第6層錐体ニューロンからのフィードバック投射
視床回路の分類
一次中継(First-Order Relays)
- ドライバーの起源: 上行性経路
- 機能: まだ皮質で処理されていない情報を伝達
高次中継(Higher-Order Relays)
- ドライバーの起源: 第5層錐体ニューロン
- 機能: 視床を介して一つの皮質領域から別の皮質領域への情報伝達
- 重要性: 霊長類視床核回路の半数以上が高次中継とされ、皮質-視床-皮質機能における視床の複雑な役割を示唆
視床皮質システム
視床、皮質、関連脳構造間の結合により3つの視床皮質システムが構成されます:
- 感覚系
- 運動系
- 連合系
これらのシステムは個別に記述されるが、密接に相互結合している。
注:この分類は過去20年間で視床回路の理解の深化に伴い洗練されてきました。
機能回路
視床、皮質、および特定の関連する脳構造間の結合は、それぞれが機能回路の異なるパターンを持つ3つのタイプの視床皮質システムを構成する。これらの3つのシステム―感覚、運動、および連合システム―はここでは個別に記述されるが、密接に相互結合している。
表1.2-3. 視床核の結合
特異的中継核
| 核 | 求心性線維 | 遠心性線維 | 機能 |
|---|---|---|---|
| 前核 | 視床下部乳頭体<br>海馬体 | 帯状皮質、海馬傍回 | 感情の表出<br>学習、記憶 |
| 腹側前核 | 淡蒼球<br>黒質 | 運動前皮質<br>前頭眼野 | 四肢運動の制御 |
| 腹側外側核 | 小脳歯状核 | 運動皮質、運動前皮質 | 眼、顔面、頭部運動の制御 |
| 腹側後外側核 | 内側毛帯および脊髄視床路 | 体性感覚皮質 | 頭部および四肢運動の制御 |
| 腹側後内側核 | 三叉神経感覚核 | 体性感覚皮質 | 疼痛、温度、触覚、圧覚、固有感覚の処理 |
| 内側膝状体核 | 下丘 | 聴覚皮質 | 聴覚 |
| 外側膝状体核 | 視索 | 視覚皮質 | 視覚 |
連合中継核
| 核 | 求心性線維 | 遠心性線維 | 機能 |
|---|---|---|---|
| 外側背側核 | 被蓋前野 | 帯状皮質、楔前部後皮質、海馬台 | 感情の表出 |
| 外側後核 | 上丘<br>被蓋前野 | 頭頂皮質<br>楔前部 | 感覚統合 |
| 視床枕 | 上丘<br>小脳 | 側頭皮質、頭頂皮質、後頭皮質 | 視覚、聴覚、体性感覚情報の統合 |
| 内側背側核 | 扁桃体、視床下部、脳幹 | 前頭前皮質 | 感情の表出、感覚情報の統合 |
びまん性投射核
| 核 | 求心性線維 | 遠心性線維 | 機能 |
|---|---|---|---|
| 正中核 | 網様体、視床下部、線条体、海馬 | 線条体、海馬、辺縁皮質 | 皮質興奮性の調節 |
| 髄板内核 | 網様体、脊髄視床路、淡蒼球 | 基底核、大脳皮質 | 皮質活動の制御 |
| 網様核 | 大脳皮質 | 他の視床核 | 視床活動の統合と制御 |
注釈:
- この表には各視床核への皮質入力は含まれていない。
- Patestas MA, Gartner LP. A Textbook of Neuroanatomy. 2nd ed. Wiley 2016より許可を得て改変。Copyright 2016 by John Wiley & Sons, Inc.
視床皮質感覚システム
視床皮質感覚システムの構成にはいくつかの一般的な原理が存在する。
第一原理:感覚受容器から視床への伝達
感覚受容器は外部環境の特定の刺激を神経インパルスに変換する。このインパルスは、しばしば脊髄と延髄の介在核を経由して上行し、最終的に視床の特異的中継核でシナプスを形成する。
第二原理:地形的構成(トポグラフィー)
末梢感覚受容器から視床および皮質への投射は地形的構成を示す。すなわち、外界の特定の部分が脳の特定の領域にマッピングされている。
例えば、体性感覚システムでは、身体の特定の部位に関する情報を伝える軸索が、視床腹側後核の離散的な部分でシナプスを形成する。具体的には:
- 腹側後内側核:頭部に関する入力を受ける
- 腹側後外側核:身体の残りの部分に関する入力を受ける
これらの視床核は一次体性感覚皮質に地形的に投射し、そこで身体の対側半分の複数の表現が見つけられる。これらの表現は歪んでおり、指などの感覚受容器が密に分布する領域は、一次体性感覚皮質において不釣り合いに大きく表現されている。
第三原理:サブモダリティによる分離処理
場合によっては、感覚入力は伝達される情報のサブモダリティに応じて分離された方式で視床に到達する。入力は並列的に処理され、特定の経路がサブモダリティの処理に専念している場合がある。
例えば、体性感覚システム(図1.2-25)では:
- 内側毛帯:触覚および固有感覚情報を伝える線維の大部分
- 脊髄視床路:疼痛と温度情報を伝える線維
両経路とも腹側後視床核で終止する。一部の触覚情報は脊髄視床路で伝達されるものの、疼痛と温度のサブモダリティは、視床に上行する際に触覚および固有感覚入力から大部分が分離されている。
第四原理:収束
感覚経路は収束を示す。すなわち、一次感覚野は感覚情報を処理し、単一モダリティ連合野に投射する。その後、単一モダリティ野は多モダリティ連合野に投射し収束する。
体性感覚システムにおける収束の例
前頭頂葉に位置する一次体性感覚皮質は、細胞構築学に基づいて4つの領域に分けられている。ブロードマンによって1、2、3a、3bと番号付けされた各細胞構築学的領域は、身体の地形的表現を含んでいる。
これらの領域は密に相互接続され、すべてがS-II野(ブロードマン野40および43)における体性感覚処理の次のレベルに投射する。
フィードフォワードとフィードバック投射
- フィードフォワード投射:一つの処理レベルからより高次の処理レベルへの投射
- 皮質層2および3から起始し、第4層で終止
- フィードバック投射:より高次の処理レベルから単純なレベルへの相互接続
- 主に第5層および第6層から起始し、第4層以外で終止
高次処理領域
体性感覚情報のさらなる処理は、S-II野からフィードフォワード投射を受ける後頭頂皮質の7b野などの高次体性感覚野で行われる。
後頭頂皮質病変の影響
後頭頂皮質の病変は、そこで処理される情報の複雑さを反映している。後頭頂病変を受けた人では:
- 感覚刺激の意義を理解する能力が障害される
- 極端な場合、対側感覚無視および不注意が生じる
