
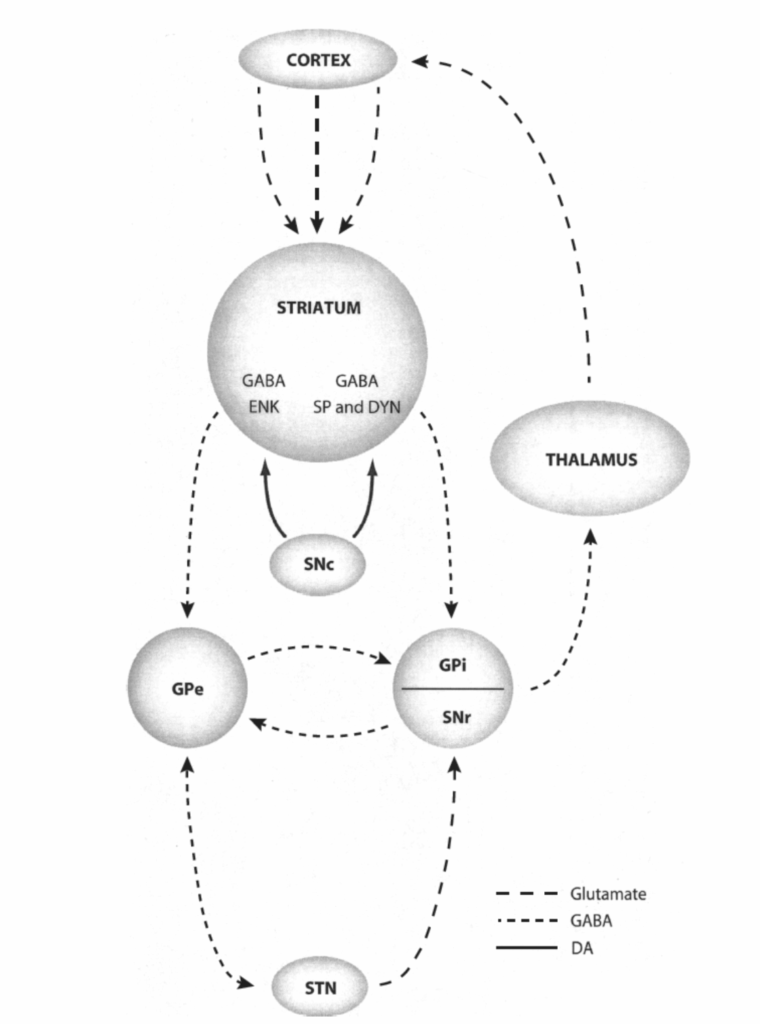
大脳基底核の内在回路の翻訳
図1.2-31. 大脳基底核の内在回路の模式図
皮質ニューロンは線条体に投射する。サブスタンスP(SP)を含有する線条体ニューロンは、淡蒼球内節(GPi)に直接抑制性投射を送る一方、エンケファリンを含有するニューロンは、淡蒼球外節(GPe)のGABAニューロンに抑制性投射を行い、これらのGABAニューロンはGPiに投射する。視床下核(STN)はGPeからの投射を受け、両方の節に投射し返す。STNとGPeは続いて黒質網様部(SNr)に投射し、SNrは視床に投射する。最終的に、視床は大脳皮質に投射し返し、ループを完成させる。
大脳基底核の出力
淡蒼球内節は大脳基底核の出力の大部分を担う(図1.2-32)。淡蒼球のこの部分は、視床の腹外側核および腹前核、ならびに髄板内視床核、特に正中中心核に投射を提供する。黒質網様部もまた、腹前核および腹外側視床核に投射を提供する。
腹外側核および腹前視床核のこれらの部分は、運動前野および前頭前野に投射する。運動前野および前頭前野から一次運動野への投射の結果として、大脳基底核は一次運動野の出力を間接的に影響することができる。さらに、大脳基底核の皮質出力は著明な収束を示す。線条体は新皮質の全領域から求心性線維を受けるが、淡蒼球および網様部の最終的な出力は、主に視床を通じて新皮質のはるかに小さな部分である運動前野および前頭前野領域に伝達される。
大脳基底核の神経回路の機能的帰結は、使用される神経伝達物質の文脈でも考慮することができる(図1.2-31および1.2-32参照)。皮質からの求心性線維は興奮性神経伝達物質であるグルタミン酸を使用すると考えられているため、皮質求心性線維はおそらく、それらが終止する大脳基底核の構造を興奮させる。大脳基底核内の多くの処理経路は、抑制性神経伝達物質GABAを使用する。最後に、大脳基底核の出力経路である淡蒼球および黒質網様部もまたGABAを使用する。皮質求心性線維からの興奮は、大脳基底核の連続した抑制経路のために、最終的に大脳基底核の標的構造を脱抑制する。
歴史的に、運動系は錐体路系(皮質脊髄路)と錐体外路系(大脳基底核)成分に分けられてきた。これらのシステムの名称は、脳幹の延髄腹側面を特徴づける両側性白質構造である錐体における前者の存在に由来する。この錐体路系と錐体外路系への分類は、各システムの病変が異なる運動症候群をもたらすという臨床所見によって支持されている。例えば、錐体外路系の病変は不随意運動、筋緊張の変化、および運動の緩慢化をもたらし、錐体路系の病変は痙縮と麻痺をもたらす。これらの所見のために、錐体路系と錐体外路系は随意運動と不随意運動を独立して制御すると考えられていた。
しかし、この分類はいくつかの理由でもはや正確ではない。第一に、小脳など、従来の錐体路系および錐体外路系以外の脳の他の構造が運動の制御に関与している。第二に、錐体路系と錐体外路系は独立していない。これらのシステムの神経回路は相互接続されている。例えば、大脳基底核は大脳皮質の特定の領域を通じて運動行動に影響を与え、これらの領域は直接的に(皮質脊髄路を通じて)または間接的に(特定の脳幹核を通じて)運動活動を産生する。
最後に、大脳基底核は運動の制御において重要であるが、この神経システムは脳の他の機能にも関与しているようである。非ヒト霊長類における大脳基底核の接続に関するより最近の研究も、これらの構造が認知機能において役割を果たすことを支持している。背外側前頭前野(DLPFC)は、淡蒼球内節内の特定の場所からの投射の標的である視床の部分から入力を受けることが示されており、明確な淡蒼球視床皮質経路の証拠を提供している。前頭前野および後頭頂領域などの大脳皮質の連合領域を一次運動野における運動活動の制御とリンクすることに加えて、大脳基底核の出力の一部は前頭前野の領域に向けて戻されているようである。これらの所見は、前頭前野と大脳基底核の間に「閉じた」ループが存在し、これらはおそらく運動機能ではなく認知機能を持つことを示唆している。
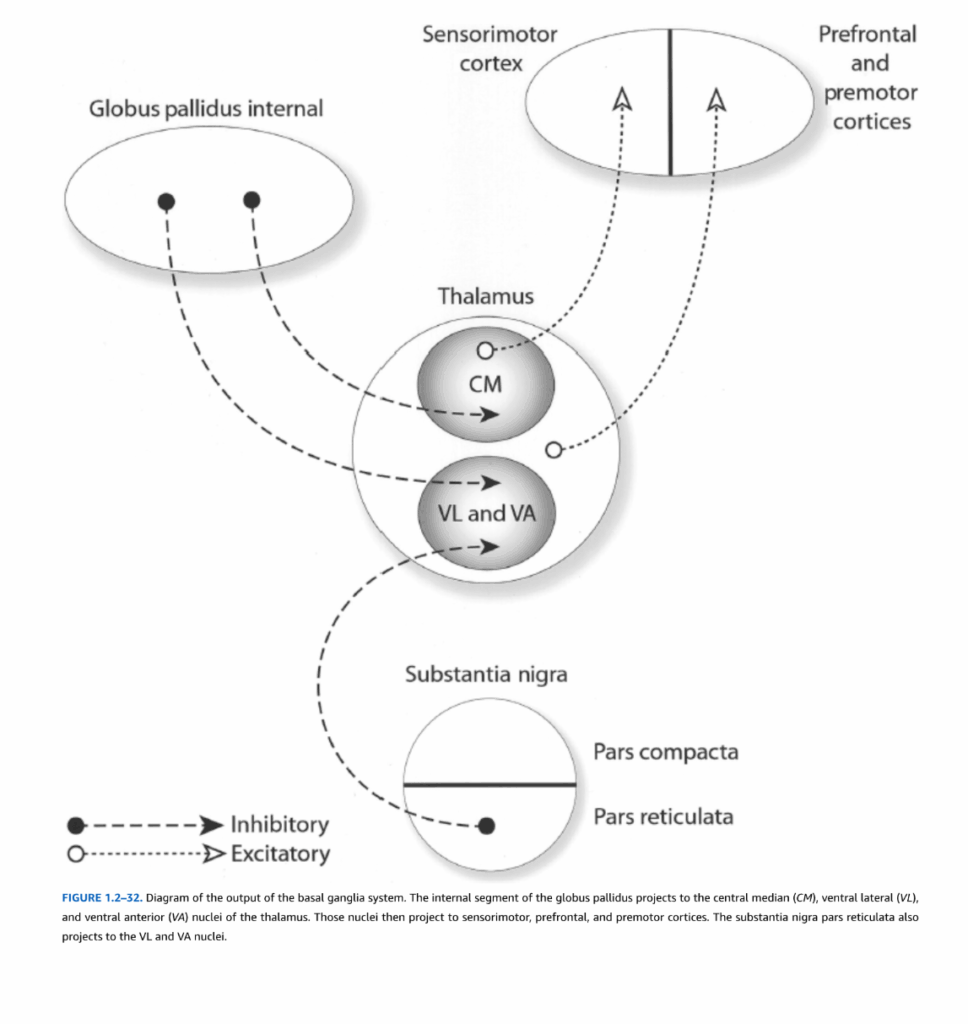
図1.2-32. 大脳基底核系の出力の模式図
淡蒼球内節は、視床の正中中心核(CM)、腹外側核(VL)、および腹前核(VA)に投射する。これらの核は続いて感覚運動野、前頭前野、および運動前野に投射する。黒質網様部もまたVL核およびVA核に投射する。
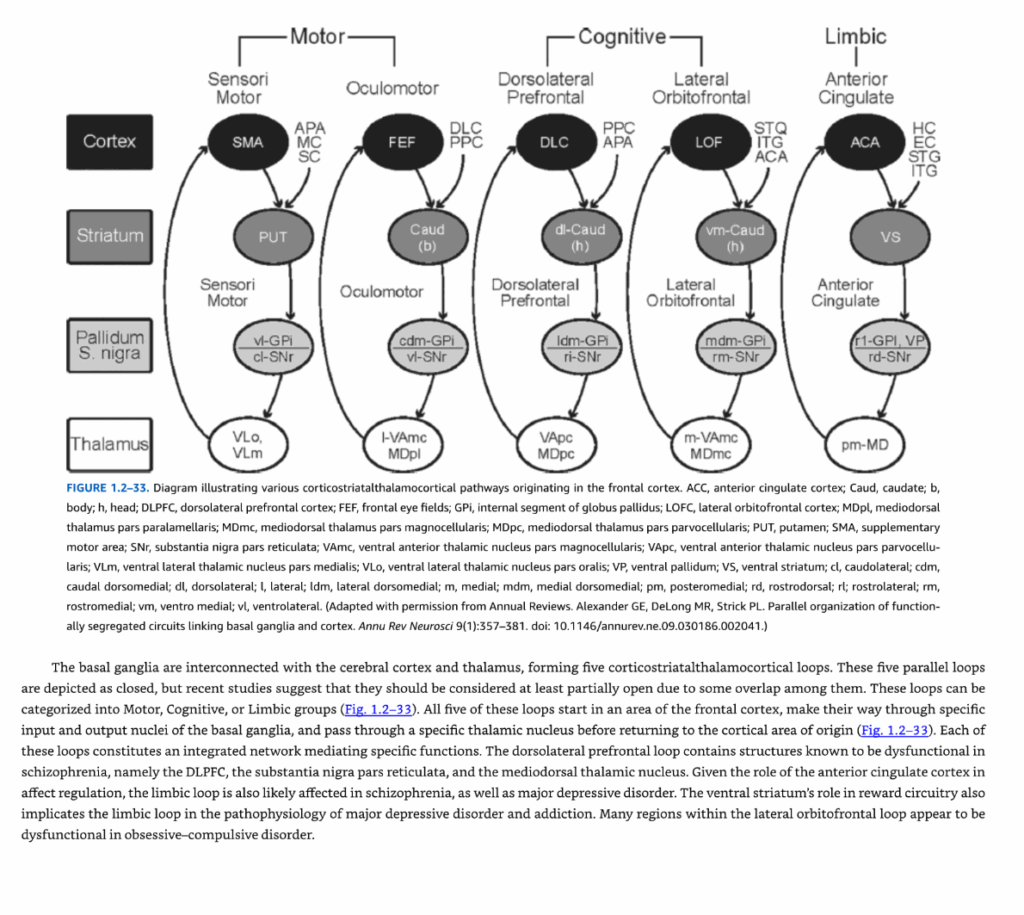
図1.2-33. 前頭皮質に起源を持つ様々な皮質線条体視床皮質経路を示す模式図
略語の説明:
- ACC:前帯状皮質
- Caud:尾状核(b:体部、h:頭部)
- DLPFC:背外側前頭前野
- FEF:前頭眼野
- GPi:淡蒼球内節
- LOFC:外側眼窩前頭皮質
- MDpl:視床背内側核傍層板部
- MDmc:視床背内側核大細胞部
- MDpc:視床背内側核小細胞部
- PUT:被殻
- SMA:補足運動野
- SNr:黒質網様部
- VAmc:腹前視床核大細胞部
- VApc:腹前視床核小細胞部
- VLm:腹外側視床核内側部
- VLo:腹外側視床核口側部
- VP:腹側淡蒼球
- VS:腹側線条体
方向の略語: cl:尾外側、cdm:尾側背内側、dl:背外側、l:外側、ldm:外側背内側、m:内側、mdm:内側背内側、pm:後内側、rd:吻背側、rl:吻外側、rm:吻内側、vm:腹内側、vl:腹外側
(Annual Reviews許可の下で改変。Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 9(1):357-381. doi: 10.1146/annurev.ne.09.030186.002041.)
皮質線条体視床皮質ループ
大脳基底核は大脳皮質および視床と相互接続し、5つの皮質線条体視床皮質ループを形成している。これらの5つの並列ループは閉じたものとして描かれているが、最近の研究では、それらの間にある程度の重複があるため、少なくとも部分的に開いたものと考えるべきであることが示唆されている。これらのループは運動系、認知系、または辺縁系のグループに分類することができる(図1.2-33)。
これら5つのループはすべて前頭皮質の領域で始まり、大脳基底核の特定の入力核および出力核を通り、特定の視床核を通過してから皮質の起始領域に戻る(図1.2-33)。これらの各ループは、特定の機能を媒介する統合されたネットワークを構成している。
背外側前頭前野ループは、統合失調症において機能不全であることが知られている構造、すなわち背外側前頭前野(DLPFC)、黒質網様部、および視床背内側核を含んでいる。前帯状皮質の感情調節における役割を考えると、辺縁系ループも統合失調症および大うつ病性障害において影響を受ける可能性が高い。腹側線条体の報酬回路における役割は、辺縁系ループが大うつ病性障害および依存症の病態生理学に関与していることも示唆している。外側眼窩前頭ループ内の多くの領域は、強迫性障害において機能不全であるように見える。
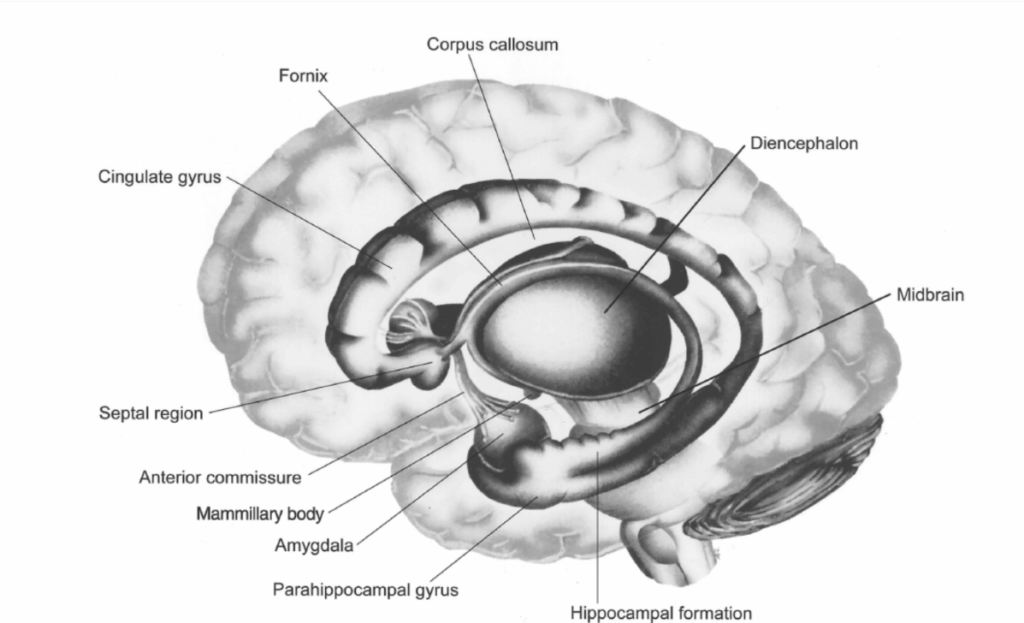
図1.2-34. 辺縁系の主要な解剖学的構造の模式図
帯状回と海馬傍回は「辺縁葉」を形成し、間脳と大脳半球の境界部に位置する組織の縁を構成している。(Hendelman WJ. Student’s Atlas of Neuroanatomy. WB Saunders; 1994:179より改変。Copyright c 1994 Elsevier.)
辺縁系
情動体験と表出の神経基盤としての辺縁系の概念は、豊富だが論争の多い歴史を持っている。100年以上前、Paul Brocaは辺縁(ラテン語のlimbus、「境界」を意味する)という用語を、間脳と大脳半球の境界部に位置する帯状回と海馬傍回を含む大脳皮質の湾曲した縁に適用した(図1.2-34)。1937年、James Papezは主に解剖学的データに基づいて、これらの皮質領域が海馬、乳頭体、前視床と回路でつながり、情動行動を媒介すると仮定した(図1.2-35)。この概念は、Heinrich KluverとPaul Bucyの研究によって支持された。彼らは、回路の構成要素を破壊する側頭葉病変が、非ヒト霊長類の情動反応を変化させることを示した。1952年、Paul MacLeanは、Brocaの辺縁葉と関連する皮質下核を情動の神経基盤として記述するために、辺縁系という用語を作った。
しかし、過去40年間で、一部の辺縁系構造(例えば海馬)が記憶などの他の複雑な脳過程にも関与していることが明らかになった。さらに、伝統的な辺縁系構造の結合性に関する知識の拡大により、辺縁系の境界を定義することがますます困難になっている。これらの制限にもかかわらず、辺縁系の概念は、特定の終脳構造とその認知過程を、自律神経系、体性、および内分泌機能を制御する視床下部とその出力経路に関連づける回路を記述する有用な方法である可能性がある。
主要構造
上述のように、辺縁系を構成する脳構造について一致した見解は存在しない。本節では、辺縁系の構成要素として最も一般的に挙げられる脳領域を含む:帯状回と海馬傍回(辺縁皮質)、海馬体、扁桃体、中隔野、視床下部、および関連する視床と皮質領域。
辺縁皮質
辺縁皮質は2つの一般的な領域、帯状回と海馬傍回から構成される(図1.2-34参照)。帯状回は、脳梁の背側にあり脳梁に沿って走るC字型の皮質の伸展で、脳梁との関係によって定義されるいくつかの皮質領域を含む(図1.2-36)。
例えば、膝下前帯状皮質は脳梁の腹側にあり、膝の前境界から嘴部まで伸びている(図1.2-36)。膝下前帯状皮質は、Brodmann野25と、Brodmann野32および24の脳梁下部分から構成される。膝下前帯状皮質は、扁桃体や視床下部など、うつ病にも関連することが多い脳領域である他の辺縁系構造と接続している。
最近の画像研究により、膝下前帯状皮質はうつ病の人において過活動であり、抗うつ薬、電気けいれん療法、反復経頭蓋磁気刺激などの様々な治療により、この領域の活動が減少することが明らかになった。さらに、治療抵抗性うつ病患者の予備研究において、膝下前帯状皮質とその下層白質の深部脳刺激(DBS)が症状の軽減に有効であることが証明されている。
DBS刺激後に記録された反応は、膝下前帯状皮質の直接的な活性化だけでなく、扁桃体、視床下部、海馬を含む、それに直接接続した脳領域の活性化からも生じる可能性があることに注意することが重要である。これらの領域は、脳梁束、鉤状束、皮質下束を介して膝下前帯状皮質と接続しており、これらの白質路の刺激がDBSの抗うつ反応に必要である。さらに、背外側前頭前野(DLPFC)や島皮質など、膝下前帯状皮質に直接接続していない領域のシナプス間活性化も、DBSの抗うつ効果において役割を果たす可能性がある。
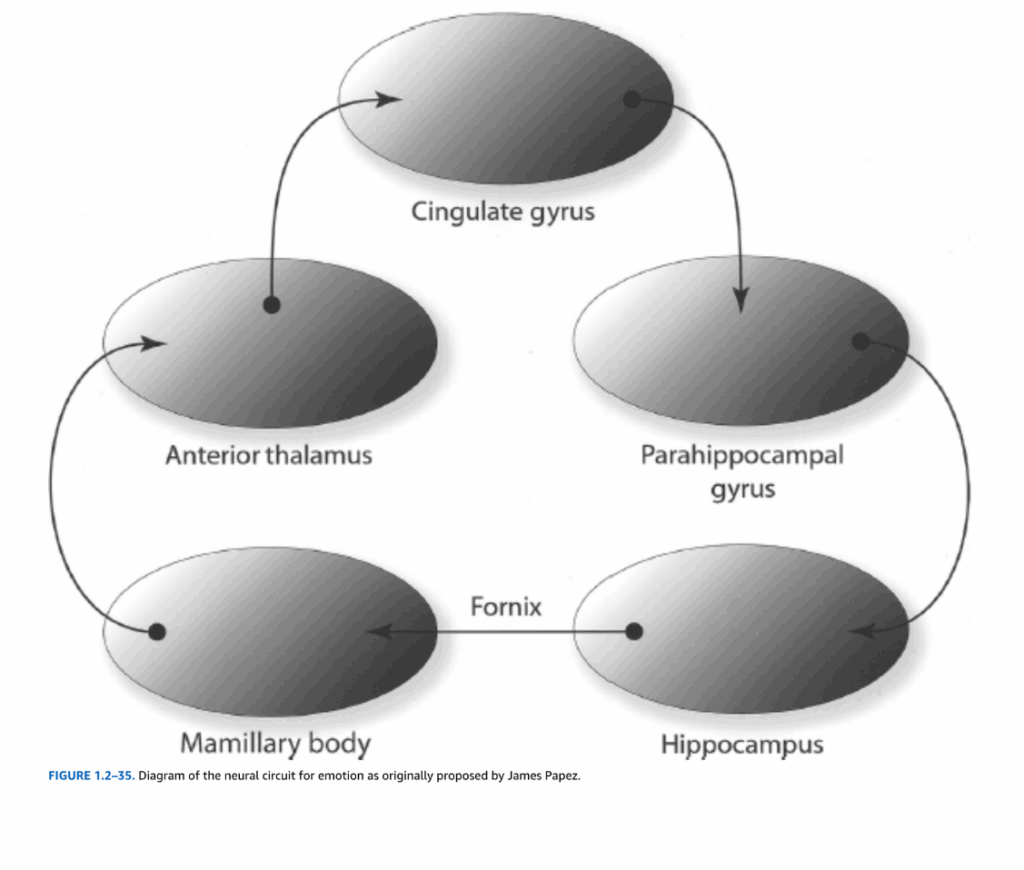
図1.2-35. James Papezによって最初に提唱された情動の神経回路の模式図
