
構築
ニューロンとその突起は多くの異なる方法でグループを形成し、これらの組織化パターン、または構築は、いくつかのアプローチによって評価することができます。
神経細胞体の分布パターンは細胞構築と呼ばれ、神経細胞体の核と細胞質中のリボヌクレオチドを染色するニッスル染色と呼ばれるアニリン染料によって明らかになります。ニッスル染色は、ニューロンの相対的な大きさと充填密度を示し、その結果、大脳皮質の異なる層へのニューロンの組織化を明らかにします。アルツハイマー病(『精神障害の診断と統計マニュアル』第5版[DSM-5]では「アルツハイマー型による主要神経認知障害」と呼ばれる)などの特定の病理学的状態では、ニューロンの変性と喪失により、一部の脳領域の細胞構築に顕著な変化が生じます(図1.2-2)。
銀染色などの他の組織学的技術は、軸索のミエリン被覆を選択的に標識し、その結果、脳のミエリン構築を明らかにします。例えば、視覚情報の処理に関与する側頭皮質の一部である領域MTなどの大脳皮質の特定の領域は、深層皮質層における重厚なミエリン化の特徴的なパターンによって同定することができます。ミエリン化の進行は高度に領域特異的で、出生後数年間完了しない場合があり、脳領域の機能的成熟の有用な解剖学的指標となる可能性があります。
ニューロン内の神経伝達物質、その合成酵素、または他の分子の位置を同定する免疫組織化学およびその他の関連技術は、脳の化学構築を決定するために使用できます(図1.2-3B)。場合によっては、これらの技術は、細胞構築では検出が困難な脳の化学構築における顕著な領域差を明らかにします。
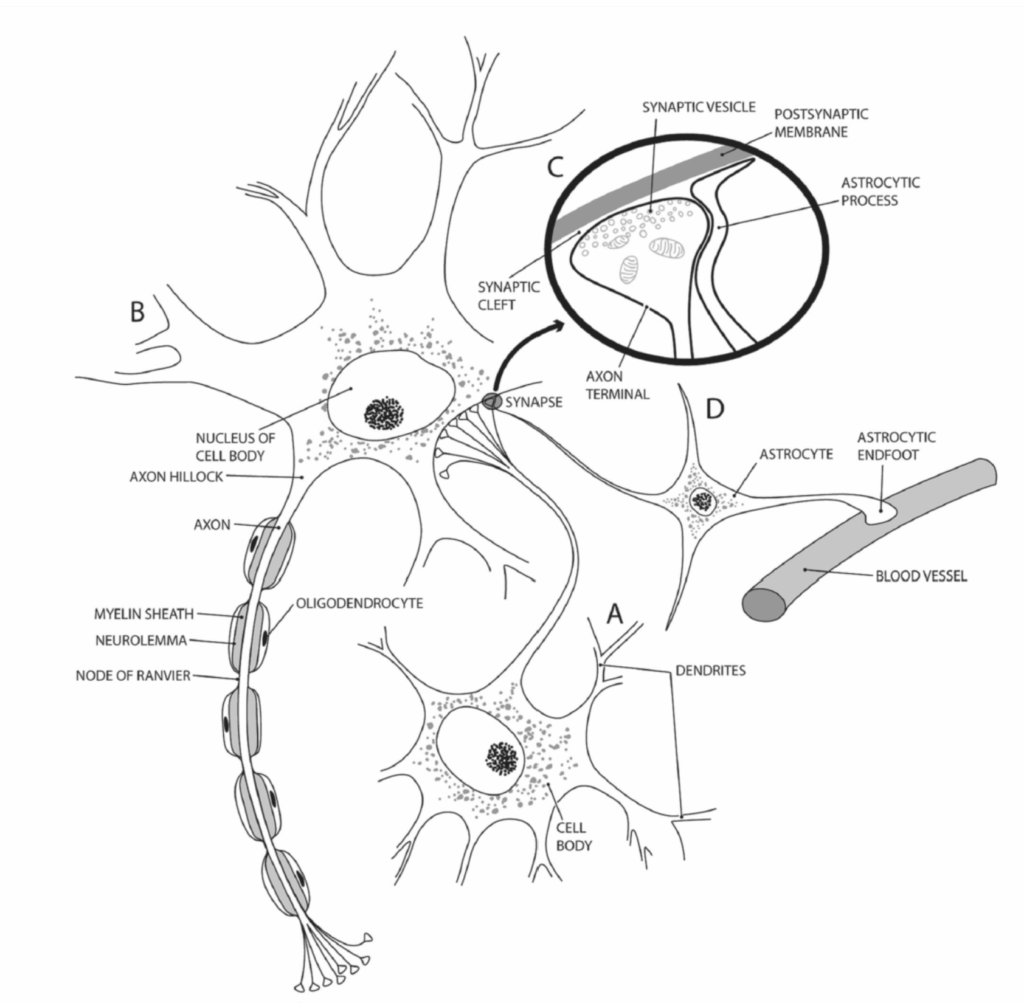
図1.2-1. 典型的なニューロン、オリゴデンドロサイト、およびアストロサイトの主要な特徴の図解。A、B:ニューロンの主要な特徴には、細胞体、樹状突起、主軸索、および軸索終末が含まれる。主軸索は隣接するオリゴデンドロサイトによってミエリン化されている。C:軸索終末は軸索から分岐し、シナプス小胞に貯蔵された神経伝達物質の放出を介してシナプス後ニューロンとシナプスを形成する。D:細いアストロサイト突起がシナプスを取り囲んで神経伝達を調節し、アストロサイトの血管周囲足突起が血管に接触して血流を調節する。(Gilman S, Winans-Newman S. Manter and Gatz’s Essentials of Clinical Neuroanatomy and Neurophysiology. 10th ed. FA Davis Co; 2003:2.より許可を得て改変)
表1.2-1 グリア細胞
| グリア細胞 | 位置 | 機能 |
|---|---|---|
| 原形質性アストロサイト | 灰白質において神経細胞体、樹状突起、軸索と接触;中枢神経系の外表面および中枢神経系血管の周囲に完全な裏打ちを形成 | 細胞外イオン環境の維持;グリア伝達物質と成長因子の分泌;ニューロンの構造的・代謝的支持;血流の調節 |
| 線維性アストロサイト | 白質路に沿って組織化;終足が血管、上衣、軸索、ランヴィエ絞輪と接触 | 白質における軸索の支持 |
| オリゴデンドロサイト | ミエリン化オリゴデンドロサイトは中枢神経系軸索の周囲にミエリン鞘を形成;衛星オリゴデンドロサイトは中枢神経系神経細胞体を取り囲む | ミエリン化オリゴデンドロサイト-ミエリン化;衛星オリゴデンドロサイト-不明 |
| シュワン細胞 | ミエリン化軸索の周囲にミエリン鞘を形成し、無髄軸索を包む | ミエリン化;ミエリン化軸索および無髄軸索の生化学的・構造的支持 |
| ミクログリア | 中枢神経系の灰白質および白質 | 細胞傷害・死後の残骸の清掃と貪食;出生後発達期における樹状突起棘のシナプス刈り込み;シナプス結合の監視 |
Mihailoff GA, Haines DE. The cell biology of neurons and gilia. In: Haines DE, Mihailoff GA. Fundamental Neuroscience for Basic and Clinical Applications. 5th ed. Elsevier; 2018:25.より改変

図1.2-2. ヒト嗅内皮質中間部の表層のニッスル染色切片。A:対照脳では、第2層に大型で濃染された神経細胞の集塊または島が含まれている。B:アルツハイマー病では、これらの第2層神経細胞が変性に対して特に脆弱であり、その消失により同領域の細胞構築に著明な変化が生じる。アラビア数字は皮質層の位置を示す。較正バー(200 μm)はAおよびBに適用される。
接続
人間の脳のあらゆる機能は、特定の神経回路の活動の結果である。これらの回路は、いくつかの発達プロセスの結果として形成される。まず、各ニューロンは軸索を伸長させる。これは最終的な位置に移動した後、または場合によっては移動前に起こる。軸索が特定の経路に沿って成長することは、環境からの分子的手がかりによって導かれ、最終的に特定の標的ニューロンとのシナプスの形成につながる。軸索の投射は非常に精密であるが、一部の軸索は初期段階で過剰な数の軸索分枝(側枝)を作り出し、成人の脳に存在するよりも広範囲の標的と接触する。思春期後期になると、特定のニューロンの接続は、不適切な標的への軸索投射の剪定または除去によって焦点化される。シナプスと軸索の除去の発達タイミングは、脳の領域間で高度に特異的であるようである。
成人の脳内では、ニューロン間または神経回路間の接続は、組織化のいくつかの重要な原則に従っている。第一に、脳領域間の多くの接続は相互的である。つまり、各領域は軸索投射を送る領域から入力を受ける傾向がある。場合によっては、ある領域から生じる軸索が別の領域の相互投射ニューロンを直接神経支配することがある。他の場合では、局所回路介在ニューロンが、入力軸索と相互接続を提供する投射ニューロンの間に介在している。一部の投射では、相互接続は間接的であり、初期の脳領域を神経支配する前に、一つまたは複数の追加の脳領域とシナプスを通過する。さらに、脳領域内の接続も相互性を示す。例えば、サルの前頭前皮質では、軸索追跡研究により、第2層と第3層の錐体ニューロンの軸索と細胞体が一連の離散した縞状に配列されていることが示されている(図1.2-4)。このシステムにおける相互性は、個々の内在性および連合縞内での順行性標識軸索と逆行性標識ニューロンの共登録によって表される。さらに、順行性標識軸索終末は、個々の縞内で逆行性標識樹状突起棘上に非対称シナプスを形成し、これらの接続における相互性のさらなる証拠を提供している。
