
1.13 精神医学における免疫と脳の相互作用
EBRAHIM HAROON, M.D., ANDREW H. MILLER, M.D., AND CHARLES L. RAISON, M.D.
免疫能力は、すべての多細胞生物にとって不可欠です。免疫系と中枢神経系(CNS)との関係は、環境からの挑戦に対する成功した適応の根底にある可能性があります。ますます、免疫と脳の間のコミュニケーションが多くの精神疾患および身体疾患の原因となり得ることが分かってきており、これは複数のライフスタイルや個人の危険因子の結果です。図1.13-1は、これらの危険因子の概要を示しています。しかし、長らく脳は、身体の免疫系の影響を受けない「免疫特権器官」であると考えられてきました。実際、血液脳関門(BBB)と血液脳脊髄液(CSF)関門は脳を身体の他の部分から隔てており、ごく少数の必須分子や細胞しかその強固な壁を通過できませんでした。結果として、免疫分子や細胞のCNSへのアクセスは限られていました。
代わりに、脳は、髄膜や脈絡叢にリンパ管やリンパ球、境界領域や脳実質に見られる様々なマクロファージ様細胞のサブセットなど、独自の免疫システムを採用しています。しかし、強固なBBBによって提供される保護にもかかわらず、脳の免疫システムは末梢免疫システムから自律的に機能しているわけではありません。身体の免疫システムの混乱は脳に影響を与え、その機能を変更します。脳は、その一方で、現在のおよび予測される危険に対して身体の免疫システムを形成し、準備します。脳は、その免疫システムの細胞および分子要素を、その発達を調節し、シナプス可塑性を変調するために利用してきました。本章では、この免疫と脳のコミュニケーションの基本的な要素と、この新たに得られた知識が精神医学における新しい治療法の進歩をどのように促進できるかについて議論します。
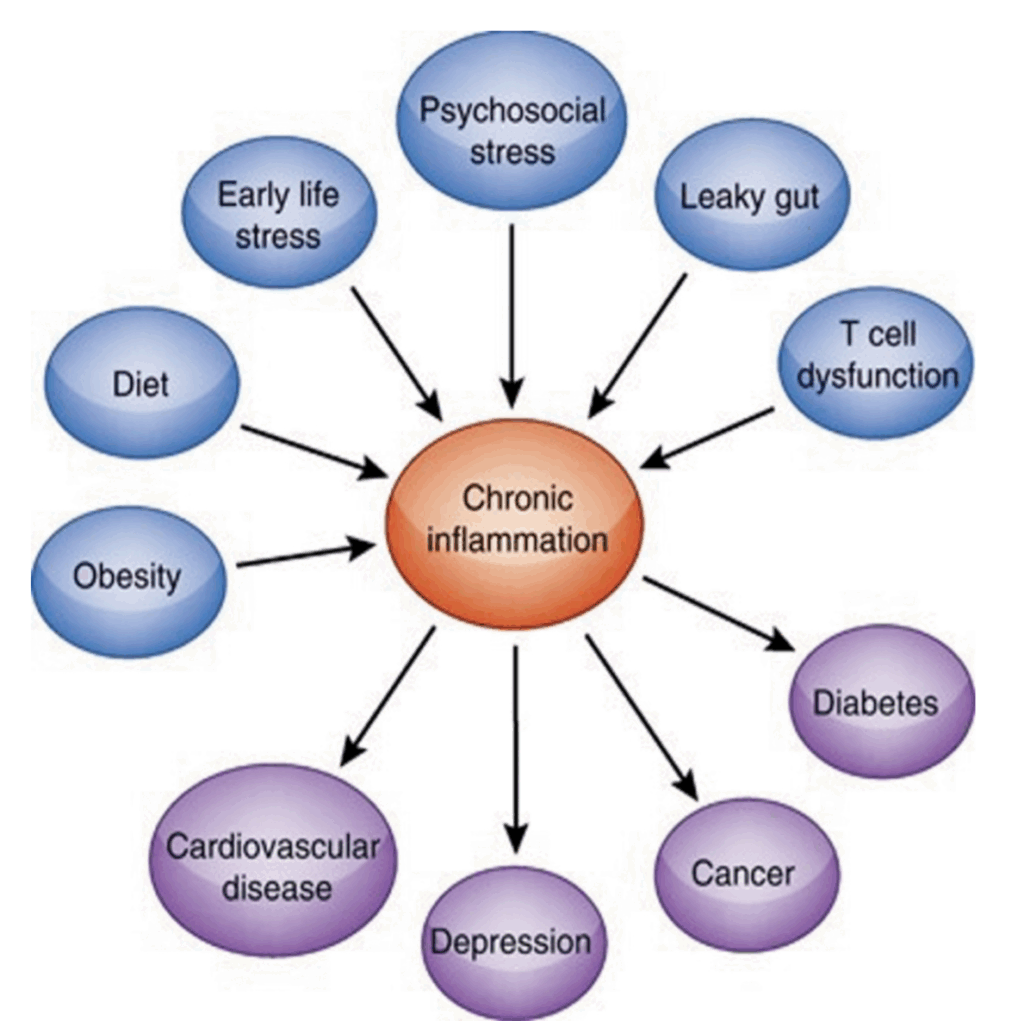
図1.13-1. 慢性的な免疫活性化に起因する健康への無数の影響が紫色で囲まれた楕円で示され、炎症の増加につながる病因因子が青色で囲まれた楕円で示されています。(カラー画像はeBookを参照)。(Haroon E, Raison C, Miller A. Psychoneuroimmunology meets neuropsychopharmacology: Translational implications of the impact of inflammation on behavior. Neuropsychopharmacology 2012;37:137-162. doi:10.1038/npp.2011.205より転載、許可を得て。)
免疫システムの構成要素
免疫システムは、以下の無数の機能を果たします。
- ウイルス、細菌、真菌、寄生虫などの異物病原体の侵入から身体を保護する。
- 新生物的に形質転換した細胞を検出し排除する。
- 特に腸内の様々な共生および常在微生物叢を監視、制御、維持する。
- 脳回路を形成する。
- 神経発達と神経伝達を調節する。
これらの機能は、免疫細胞やグリア細胞の表面にある特殊なセンサー(受容体)を介して達成されます。これらは、侵入する生物や新生物細胞からの病原性分子を認識します。細胞は、直接的な細胞間相互作用と、サイトカインと呼ばれる可溶性因子による免疫系細胞間のシグナル伝達を伴う、豊かな細胞間通信ネットワークを介して通信します。興味深いことに、免疫シグナル分子は、グリア細胞や一部のニューロンのサイトカイン受容体を介して脳細胞とも通信し、あらゆる脅威(身体的または心理社会的)に対する複雑な神経行動応答を組織化します。
表1.13-1.
免疫系の区分:自然免疫と獲得免疫
| 防御線 | 自然免疫 | 獲得免疫 |
| 理化学的バリア | 皮膚、粘膜 | 皮膚および粘膜免疫系 |
| 細胞 | 食細胞(マクロファージ、好中球、ナチュラルキラー細胞) | リンパ球(B細胞、T細胞) |
| 他の細胞に影響を与える可溶性メディエーター | マクロファージ由来サイトカイン、例:IL-1、IL-6、TNF、IFN-α | リンパ球由来サイトカイン、例:IL-2、IL-4、IL-10、IFN-γ |
| 記憶 | トレーニングされた | あり |
| 循環分子 | 補体、急性期反応物質 | 抗体 |
IL, インターロイキン; IFN-α, インターフェロンアルファ; IFN-γ, インターフェロンガンマ; TNF, 腫瘍壊死因子。
Abbas AK, Lichtman AH, Pillai, S. Cellular and Molecular Immunology. Elsevier; 2022, より許可を得て改変。
免疫系の両腕:自然免疫と獲得免疫
表1.13-1は、免疫系の2つの主要な部門、すなわち自然免疫(または生来免疫)と適応免疫(または獲得免疫)の機能的な違いを示しています。免疫細胞は骨髄の幹細胞に由来しますが、骨髄系幹細胞系とリンパ系幹細胞系に分化します。骨髄系細胞は赤血球、血小板、単核食細胞を生み出し、リンパ系細胞はT細胞とB細胞、そしてナチュラルキラー(NK)細胞を生み出します。
自然免疫系と適応免疫系には、重要な違いと重複があります。自然免疫の構成要素は迅速に作用しますが、病原体や感染細胞に対しては比較的非特異的です。一方、適応免疫系は非常に特異的であり、しばしば一生続く「適応免疫記憶」を保持しています。しかし、最近のデータでは、自然免疫系も遭遇した病原体の最近の記憶を保持する可能性があることが示されています。これは「訓練された自然免疫記憶」と呼ばれる現象です。訓練された免疫とは、自然免疫細胞のエピゲノム、ゲノム、そして最終的には機能を改変することによって、病原体の短期的な「記憶」を作り出すことを含みます。
したがって、訓練された免疫は、一生続く可能性がある適応免疫記憶とは対照的に、数ヶ月間持続します。さらに、訓練された免疫は、適応免疫とは異なり、2回目の遭遇に備えて抗体を生成することはありません。言い換えれば、訓練された免疫は最近の記憶に似ており、適応免疫は長期記憶に似ています。そのため、自然免疫(訓練されたものか急性的なものかに関わらず)は、T細胞およびB細胞が媒介する適応/獲得免疫よりも、進化的に原始的で特異性が低いと考えられています。興味深いことに、NK細胞のようなリンパ球系の細胞は、自然免疫細胞として機能します。しかし、実際には、上記のすべての免疫のノードは相互作用し、協力し合っています。最も注目すべきは、免疫系が高いレベルの生物学的冗長性を示し、脅威や危険に直面した場合のフェイルセーフメカニズムとして、いくつかの分子が同じ機能的標的に収束することです。
自然免疫系
自然免疫監視。 自然免疫系は通常、内部または外部の脅威との最初の接触点です。自然免疫を媒介する細胞は、部分的に腸からの微生物成分への曝露によって、常に一定の活性化レベルに維持されています。これらの細胞は事前の活性化を必要とせず、侵入する病原体を遅延なく認識します。したがって、それらは免疫応答の初期段階で感染因子に対する重要な第一防御線を提供します。単核食細胞とNK細胞は、自然免疫細胞の例です。NK細胞は、自然免疫の特殊ではあるが重要な構成要素です。これらの細胞はリンパ芽球系の遺伝的背景から生じますが、適応免疫細胞と同様に自然免疫細胞として機能します。それらは、ウイルスに感染した細胞に結合し、パーフォリンを含む細胞溶解因子を放出することによって、ウイルス感染細胞を破壊することができます。NK細胞は、新生物的に形質転換した宿主細胞、特に造血起源のものを認識し破壊することもでき、したがって一部のがんから保護します。図1.13-2は、異なる免疫細胞の造血起源を示しています。
単核食細胞には以下が含まれます。
- 単球(血中を循環する)
- マクロファージ(循環から組織に入る単球系細胞)
- ミクログリア(脳におけるマクロファージに相当する細胞)
- 樹状細胞(組織に常在する免疫細胞)
- 細網細胞(細網内皮器官[脾臓、リンパ節、リンパ器官]に常在する免疫能力のある線維芽細胞)
注目すべきは、細網内皮器官が、自然免疫系と適応免疫系との間で免疫情報が交換される交差点である可能性があることです。これらの細胞は適応免疫細胞と同様にリンパ芽球系の遺伝的背景から生じますが、自然免疫細胞として機能します。それらは、ウイルスに感染した細胞に結合し、パーフォリンを含む細胞溶解因子を放出することによって、ウイルス感染細胞を破壊することができます。NK細胞は、新生物的に形質転換した宿主細胞、特に造血起源のものを認識し破壊することもでき、したがって一部のがんから保護します。
脅威の検出と免疫シグナル伝達。 図1.13-3は、精神医学に関連する自然免疫活性化中に起こる主要なイベントをまとめたものです。脅威が認識されると、自然免疫系は免疫系および神経系の他のすべての構成要素に警戒信号を送ります。免疫分子とタンパク質は、免疫刺激に対するシステム全体での応答を調整する鍵となります。最も重要な因子は以下で説明します。
表面パターン認識受容体。 自然免疫細胞は、循環抗原を監視し、病原性産物を発見すると迅速な免疫応答を動員する免疫監視システムの一部です。自然免疫細胞は、比較的単純な分子パターン認識受容体(例:トル様受容体、またはTLR)を介して細胞外病原体(例:細菌や寄生虫)を認識します。TLRは2種類の病原性分子パターンを認識します。1つ目は、感染因子に進化的に保存されている分子パターンで、病原体関連分子パターン(PAMPs)と呼ばれます。2つ目は、死滅する細胞や新生物細胞からのデオキシリボ核酸(DNA)やその他の残骸の認識を指し、これらは危険関連分子パターン(DAMPs)として知られています。したがって、DAMPsによって誘発される免疫応答は無菌性炎症と呼ばれ、アデノシン三リン酸(ATP)、DNA、ヒートショックタンパク質(HSPs)、高移動度群ボックス1(HMGB1)など、死滅または損傷した細胞から漏れ出す生化学物質によって誘発される可能性があります。その後、これらの細胞は病原体や死滅した細胞を貪食・分解することによって、脅威を迅速に中和します。
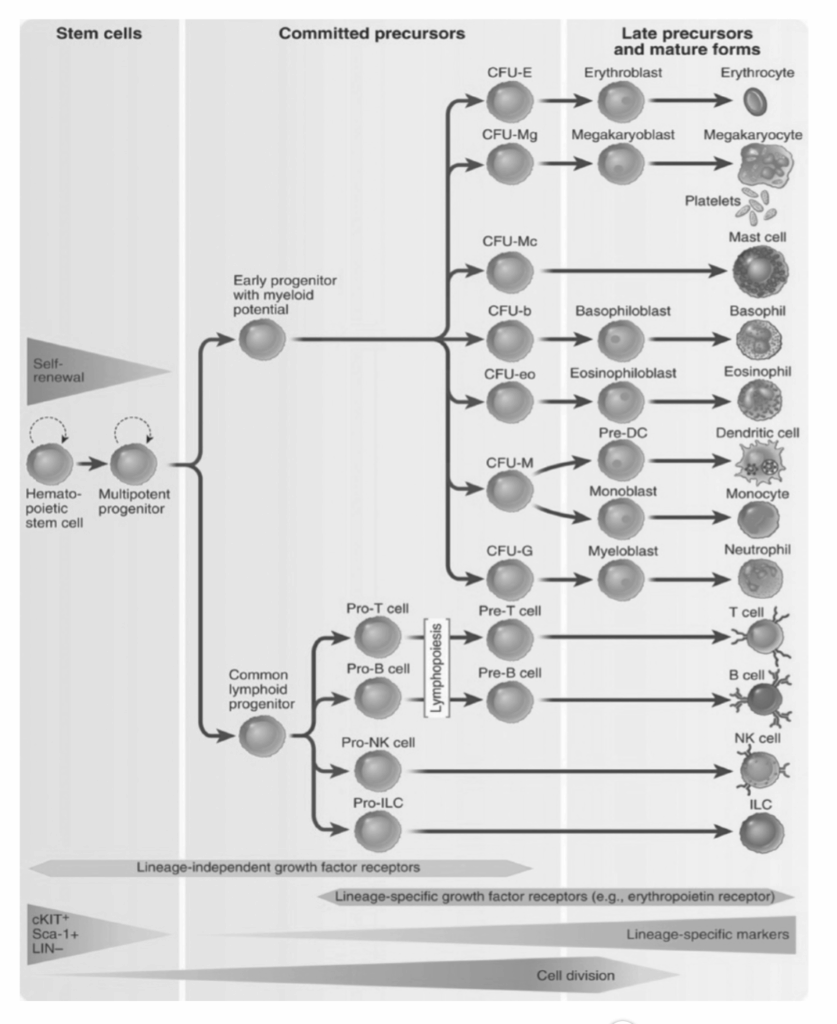
図1.13-2. 血液細胞の主要な系統の発生をこの造血樹に示しています。異なる系統の成熟を促進する主要なサイトカインは、以下の参照文献に記載されています。CFU, コロニー形成単位; CFU-Mc, CFUマスト細胞; CFU-b, CFU B細胞; CFU-eo, CFU好酸球; CFU-G, CFU顆粒球; CFU-M, CFUマクロファージ; DC, 樹状細胞; ILCs, 自然リンパ球; NK, ナチュラルキラー。(Abbas AK, Lichtman AH, Pillai, S. Cellular and Molecular Immunology. Elsevier; 2022, より許可を得て転載。)
細胞内シグナル伝達カスケード。 病原体(および一部のサイトカイン)によるTLRの活性化は、ニューロンにおけるセカンドメッセンジャーシステムに似た細胞内シグナル伝達カスケードを誘発します。これらの細胞内シグナル伝達カスケードには、**核因子κB(NF-κB)と、p38 MAPKとして知られるマイトジェン活性化プロテインキナーゼ(MAPK)**の異なるサブタイプが含まれます。NF-κBとMAPKの両方は、自然免疫応答を開始する上で哨兵的な役割を果たします。
インフラマソーム:非感染性ながらも毒性のある刺激(死んだ細胞、腫瘍細胞、あるいはストレスや苦痛の産物)に対する無菌性炎症反応は、インフラマソームによって媒介されます。インフラマソームは、骨髄系細胞内に存在する細胞質タンパク質複合体で、微生物刺激や無菌性炎症刺激に反応します。インフラマソーム機構が適時に集合すると、カスパーゼ1という酵素が活性化されます。この酵素は、インターロイキン-1(IL-1)ベータやインターロイキン(IL)-18などのサイトカインの不活性な前駆体を活性型に切断し、炎症反応を促進します。
インフラマソームは**TLR(Toll-like receptor)と協調して機能しますが、死んだ細胞や損傷した細胞からの残骸(無菌性炎症)への応答に焦点を当てています。実際、研究によると、ストレスを受けた細胞のATPからアデノシンなどのDAMP(Danger-Associated Molecular Patterns)**が放出されることで、感染性刺激がないにもかかわらず心理的ストレスが炎症を活性化するメカニズムが説明できると示唆されています。これらの研究により、ストレスを受けたニューロンから放出されるアデノシンとグルタミン酸がインフラマソームの活性化を引き起こすことが明らかになりました。
肥満は、慢性炎症のよく知られた原因です。脂肪細胞の形成と死から放出されるDAMPが、肥満における慢性の自然免疫活性化につながります。本章の範囲を超えますが、SARS-CoV-2ウイルス粒子による単球およびマクロファージにおけるインフラマソームの活性化は、COVID-19関連肺機能障害における組織損傷の多くを引き起こします。
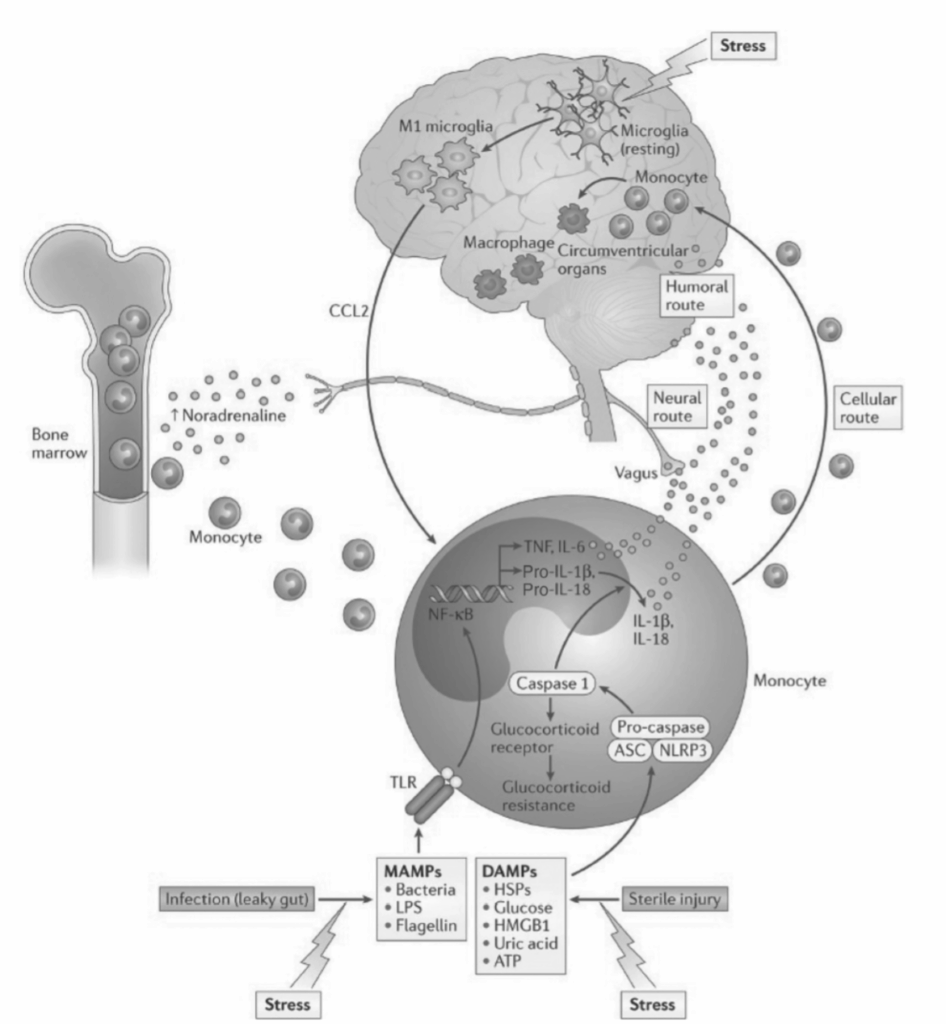
図1.13-3. 局所的に作用するサイトカインと急性期反応タンパク質
図1.13-3. は、局所的に作用するサイトカインが内皮細胞や組織間質細胞に与える影響を示しています。組織間質細胞は走化性因子を産生し、損傷部位に他の免疫細胞を呼び寄せます。内皮細胞は接着分子を産生し、免疫細胞の血管壁への縁辺化と血管外遊出を促進します。脳内では、インターロイキン(IL)-1、IL-6、腫瘍壊死因子(TNF)などの炎症性サイトカインが視床下部-下垂体-副腎(HPA)軸を活性化し、発熱や炎症の代謝要求を補う行動変化を誘発します。また、炎症性サイトカインは肝臓に急性期反応タンパク質を産生させます。
CAM:細胞接着分子、IFN:インターフェロン、NF-κB:核内因子カッパーB、PGE:プロスタグランジンE、TLR:Toll様受容体。(Cowles MK, Miller AH. Stress, cytokines and depressive illness. In: Squire LR, ed. The New Encyclopedia of Neuroscience. Elsevier: Academic Press; 2009, 519-527より許可を得て転載。)
サイトカイン
サイトカインは高分子量のタンパク質で、免疫系の主要な長距離シグナル伝達分子として機能します。その数多くの機能の一部を表1.13-2に示します。活性化された単核細胞は、腫瘍壊死因子(TNF)、IL-1、IL-6などの炎症性サイトカインを放出します。TNFとIL-1はグラム陰性菌に対する応答の主要なメディエーターであり、炎症カスケードで最も早期に放出されるサイトカインの一つであり、その後IL-6が続きます。TNFとIL-1は、サイトカインによって刺激された視床下部細胞でのプロスタグランジンの合成を増加させることで発熱を誘発する内因性発熱物質です。TNFはまた、筋細胞の消耗と食欲抑制を特徴とする悪液質を引き起こします。活性化された単核細胞は、直接的な抗ウイルス特性を持つI型インターフェロン(例:インターフェロン[IFN]-アルファ)も放出します。
急性期反応タンパク質
急性期反応タンパク質(APP)は主に肝臓で合成され、組織損傷を制限し、侵入病原体を隔離・破壊し、修復機能を始動させます。APPは陽性急性期タンパク質と陰性急性期タンパク質に分類されます。陰性急性期タンパク質は、陽性APPの合成のために生産が減少する分子です。陰性急性期タンパク質にはアルブミン、トランスフェリン、レチノール結合タンパク質が含まれ、しばしば慢性炎症の程度を測定するために使用されます(例:悪液質後)。陽性APPは炎症活動を助け、以下を含みます。
表1.13-2. 自然免疫のサイトカインメディエーター
自然免疫におけるサイトカインメディエーター
インターロイキン-6 (IL-6)
- 主要な細胞源: マクロファージ、樹状細胞 (DC)、内皮細胞、T細胞
- 主要な細胞標的と生物学的効果:
- 肝臓:急性期タンパク質の合成
- B細胞:抗体産生細胞の増殖
- T細胞:Th17分化
インターロイキン-2 (IL-2)
- 主要な細胞源: T細胞
- 主要な細胞標的と生物学的効果:
- T細胞:エフェクター細胞およびメモリー細胞への増殖と分化;制御性T細胞の発達、生存、機能の促進
- NK細胞:増殖、活性化
インターロイキン-4 (IL-4)
- 主要な細胞源: CD4+ T細胞 (Th2)、肥満細胞
- 主要な細胞標的と生物学的効果:
- B細胞:IgE、IgG4(ヒトの場合;マウスではIgG1)へのアイソタイプスイッチ
- T細胞:Th2分化、増殖
- マクロファージ:代替活性化およびIFN-γを介した古典的活性化の阻害
インターロイキン-27 (IL-27)
- 主要な細胞源: マクロファージ、樹状細胞 (DC)
- 主要な細胞標的と生物学的効果:
- T細胞:Th1分化の促進;Th17分化の阻害
顆粒球-単球コロニー刺激因子 (GM-CSF)
- 主要な細胞源: T細胞、マクロファージ、内皮細胞、線維芽細胞
- 主要な細胞標的と生物学的効果:
- 未成熟およびコミットされた前駆細胞、成熟マクロファージ:顆粒球と単球の成熟誘導、マクロファージの活性化
顆粒球コロニー刺激因子 (G-CSF, CSF3)
- 主要な細胞源: マクロファージ、線維芽細胞、内皮細胞
- 主要な細胞標的と生物学的効果:
- コミットされた造血前駆細胞:顆粒球の成熟
IFN-α (I型IFN; 複数のタンパク質)
- 主要な細胞源: 形質細胞様樹状細胞 (pDC)、マクロファージ
- 主要な細胞標的と生物学的効果:
- すべての細胞:抗ウイルス状態、クラスI MHC発現の増加
- NK細胞:活性化
IFN-γ (II型IFN)
- 主要な細胞源: T細胞 (Th1, CD8+ T細胞)、NK細胞、ILC1s
- 主要な細胞標的と生物学的効果:
- マクロファージ:古典的活性化(殺微生物機能の増加)
- B細胞:オプソニン化および補体結合性IgGサブクラスへのアイソタイプスイッチ(マウスの場合、ヒトではない)
- T細胞:Th1分化
- 様々な細胞:クラスIおよびクラスII MHC分子の発現増加、抗原処理およびT細胞への提示の増加
インターロイキン-10 (IL-10)
- 主要な細胞源: マクロファージ、T細胞(主に制御性T細胞)
- 主要な細胞標的と生物学的効果:
- マクロファージ、樹状細胞 (DC):IL-12、共刺激分子、クラスII MHCの発現阻害
腫瘍壊死因子 (TNF, TNF-アルファとも呼ばれる; TNFSF2)
- 主要な細胞源: マクロファージ、NK細胞、T細胞
- 主要な細胞標的と生物学的効果:
- 内皮細胞:活性化(炎症、凝固)
- 好中球:活性化
- 視床下部:発熱
- 筋肉、脂肪:異化作用(悪液質)
インターロイキン-1ベータ (IL-1β)
- 主要な細胞源: マクロファージ、樹状細胞 (DC)、線維芽細胞、内皮細胞、ケラチノサイト;生物学的に活性なIL-1の主要なタイプ
- 主要な細胞標的と生物学的効果:
- 内皮細胞:活性化(炎症、凝固)
- 視床下部:発熱
- 肝臓:急性期タンパク質の合成
- T細胞:Th17分化
IL-1受容体拮抗薬 (IL-1RA)
- 主要な細胞源: マクロファージ
- 主要な細胞標的と生物学的効果:
- 様々な細胞:IL-1の競合的拮抗薬
インターロイキン-17A (IL-17A)、インターロイキン-17F (IL-17F)
- 主要な細胞源: CD4+ T細胞 (Th17)、ILC3s
- 主要な細胞標的と生物学的効果:
- 上皮細胞、マクロファージ、その他の細胞種:ケモカインおよびサイトカイン産生の増加;GM-CSFおよびG-CSFの産生
形質転換増殖因子-ベータ (TGF-β)
- 主要な細胞源: T細胞(主にTregs)、マクロファージ、その他の細胞種
- 主要な細胞標的と生物学的効果:
- T細胞:増殖およびエフェクター機能の阻害;Th17およびTregの分化
- B細胞:増殖の阻害;IgAの産生
- マクロファージ:活性化の阻害;血管新生因子の刺激
- 線維芽細胞:コラーゲン合成の増加
MCP (単球走化性タンパク質)-1, (CCL2)
- 主要な細胞源: 多くの細胞
- 主要な細胞標的と生物学的効果:
- 混合白血球:リクルートメント
IL-8, (CXCL8)
- 主要な細胞源: 多くの細胞
- 主要な細胞標的と生物学的効果:
- 好中球:リクルートメント
フラクタルカイン, (CX3CL1)
- 主要な細胞源: 多くの細胞
- 主要な細胞標的と生物学的効果:
- T細胞、NK細胞、単球:リクルートメント
CCL2、CXCL8、CXC3CL1は、それぞれMCP-1、IL-8、およびフラクタルカインに対するケモカインの命名法です。
DC:樹状細胞、IFN:インターフェロン、IFN-α:インターフェロンアルファ、IFN-γ:インターフェロンガンマ、Ig:免疫グロブリン、MHC:主要組織適合性複合体、NK:ナチュラルキラー細胞、Th:ヘルパーT細胞。
Abbas AK, Lichtman AH, Pillai, S. Cellular and Molecular Immunology. 第10版. Elsevier; 2022. より許可を得て改変。
- プロテアーゼ阻害剤:例として、アルファ-1アンチトリプシン、アルファ-1アンチキモトリプシン
- 凝固タンパク質:例として、フィブリノゲン、プロトロンビン
- 補体タンパク質:例として、C2、C3、C4、C5など
- 輸送タンパク質:例として、ハプトグロビン
- 炎症性タンパク質:例として、C反応性タンパク質(CRP)、血清アミロイドA(SAA)
TNFおよびIL-1によって誘導される肝臓由来の血漿タンパク質と、IL-6によって誘導されるタンパク質の組み合わせが、急性期反応を構成します(図1.13-3を参照)。特に、肝臓におけるCRPの合成は、IL-6が肝細胞に作用することで増加します。そのため、血漿CRPは全体的な炎症活性化の優れた指標として認識されています。これは臨床検査室間で標準化されており、急性炎症(肺炎など)や慢性炎症(心血管疾患、自己免疫疾患)のリスクを予測することがよく知られています。
補体因子
肝臓によって産生される補体因子タンパク質もまた、自然免疫応答の重要な構成要素です。これらの機能的に関連するタンパク質は、細胞溶解、オプソニン作用、炎症細胞の誘引(走化性)、免疫細胞による炎症の化学メディエーターの放出刺激、組織損傷を防ぐための抗原-抗体複合体の不活性化など、自然免疫機能を補助するために秩序だった方法で相互作用します。特筆すべきは、補体タンパク質が食細胞による識別と破壊のために細胞にタグを付けることです。例えば、C4A補体などの補体タンパク質は、発達中のミクログリアによる食作用によってシナプスの神経剪定を形成します。過剰な剪定につながる異常な食作用シグナルは、自閉症や統合失調症の一部症例における病因的な役割を果たすと考えられています。新生的な証拠は、母体または胎児の感染を介した免疫学的メカニズムの活性化が、異常な補体活性化に関連する病態を悪化させる可能性があることを示しています。
獲得免疫
Tリンパ球とBリンパ球は、免疫細胞の特殊化の進化における最高の成果であり、免疫システム内の多様性、特異性、および適応機能性を担っています。獲得免疫応答には、認識相、活性化相、および抗原排除の実行相の3つの段階があります。
認識相
認識相では、感染性病原体、抗原、または腫瘍化細胞の存在が、Bリンパ球とTリンパ球の表面にある抗原特異的受容体を用いて感知されます。特定のB細胞またはT細胞上の抗原に対する受容体は、その子孫(クローン)に対して同一かつユニークです。同一の抗原受容体特異性を持つリンパ球のファミリーは、クローン系統と呼ばれます。したがって、抗原認識の多様性は、各個人に存在する膨大な数の異なるB細胞およびT細胞のクローン系統に由来します。抗原が検出されると、対応するクローン系統が選択され、活性化されます。注目すべきは、リンパ球上の表面受容体は、それぞれの病原体や死滅細胞に対して高度に特化され、カスタマイズされており、その表面タンパク質発現(分化クラスター(CD)と呼ばれる)によって分類されることです。興味深いことに、細網内皮器官は、膨大な数のクローン系統を保持する図書館として概念化されており、活性化の準備ができています。いくつかの抗原提示自然免疫細胞は、リンパ管を介してこれらの器官の洞に流入し、リンパ球と近接します。したがって、リンパ球による抗原認識は、脾臓、胸腺、リンパ節などの細網内皮器官でパラクリン的に起こる可能性があります。注目すべきは、この界面でのリンパ球の誤教育が、一部の白血病を引き起こす可能性もあることです。
活性化相
活性化相には、感染性病原体を根絶するために関連する免疫細胞の増殖と動員が含まれます。異物抗原の結合だけでは、B細胞とT細胞の活性化を誘発するには通常不十分であり、補助シグナルが必要です。重要な補助シグナルは、マクロファージや樹状細胞などの抗原提示細胞(APC)と協調して作用するTヘルパー(Th)リンパ球細胞によって分泌されるIL群のサイトカインによって生成されます。Th細胞とAPCは、免疫応答を形成し増幅するために反響的に互いを刺激します。APCはIL-1やその他のサイトカインを分泌し、これらがTh細胞を刺激してIFN-ガンマを含む多数のサイトカインを分泌させます。これによりAPCの活性がさらに増加し、最終的にその抗原提示能力が向上します。したがって、獲得免疫応答に関与するサイトカインは、多くの場合、同時に複数の機能を果たします。刺激性サイトカインの存在下での抗原への結合は、細胞増殖、分裂、Tリンパ球およびBリンパ球の増殖を制御する細胞内システムの活性化をもたらします。活性化はまた、異物抗原に対する同一の高い親和性特異性を持つ免疫細胞のクローン性増殖をもたらします。さらに、一部の子孫は、クローン性増殖中に、抗体分泌形質B細胞や細胞性免疫に関与する細胞傷害性CD8+ Tリンパ球(CTL)などの成熟したエフェクター細胞へとさらに分化します。
対照的に、活性化されたB細胞またはT細胞の一部の子孫は、同じ抗原による将来の刺激に備えて活性化の準備ができたメモリー細胞になります。したがって、同じ抗原への再曝露は、その抗原に対する最初または一次免疫応答よりも、より迅速かつ強力な二次免疫応答(したがって獲得免疫という名前)を引き起こします。注目すべきは、メモリー細胞が長年生存し、ワクチン接種を受けた個人や乳幼児期に特定の感染性病原体と初めて接触した個人が示すように、長期にわたる獲得免疫を提供する可能性があることです。同様に、過剰な免疫記憶が過敏症反応に関与する可能性もあります。
エフェクター相:エフェクター相では、病原体が中和され、排除されます。獲得免疫応答は、脅威を中和するために**液性免疫(抗体媒介性)および細胞性免疫(CTL)**のプロセスを用います。B細胞から分泌される抗体は、液性免疫の主要なエフェクターです。液性免疫は、細菌や寄生虫などの細胞外病原体との闘いに特に効果的です。CTLを用いる細胞性免疫は、標的細胞を直接殺傷するために使用されます。液性免疫とは対照的に、細胞性免疫(CTLに基づく)は、ウイルスおよび細胞内細菌感染から効果的に保護し、NK細胞と同様に、腫瘍細胞に対してある程度の防御を提供する可能性があります。T細胞はまた、ミクログリアを含むマクロファージの抗菌機能を高めるIFN-ガンマなどのマクロファージ活性化サイトカインを放出します。獲得免疫応答の制御は、エフェクター相のもう一つの重要な要素です。これには、免疫抑制性であり自己抗原に対する免疫応答を制限するCD4+CD25+制御性T細胞(CD4細胞とも呼ばれる)が含まれます。さらに、プログラム死-1(PD-1)と呼ばれる抑制性共刺激分子とそのリガンドは、T細胞活性化の制御/抑制に重要な役割を果たすことが示されています。実際、PD-1の欠損は、実験動物における自己免疫体質と関連しています。PD-1の枯渇は、SARS-CoV-2感染などの状況における「サイトカインストーム」につながるメカニズムとしても提唱されています。
自然免疫系との統合
B細胞とT細胞はまた、特定の抗原によってもたらされる脅威を中和するために、自然免疫メカニズムを動員し、強化し、指示します。例えば、循環抗体は、病原体に結合し、それらをコーティングすることによって(オプソニン作用)、病原体を中和することができます。オプソニン化された病原体は、補体因子による溶解や食作用を受けやすくなります。NK細胞や食細胞(好中球やマクロファージなど)は、抗体のFcフラグメント(結晶化可能領域)に対する受容体を持っています。さらに、補体タンパク質は、いくつかの種類の抗体のFcフラグメントに結合し、それらによって活性化されます。したがって、抗体は、自然免疫の細胞性エフェクターや溶細胞性タンパク質を病原体と連結させることができ、自然免疫プロセス自体には本来備わっていないレベルの特異性をもたらします。
サイトカインによる自然免疫と獲得免疫の同期
サイトカインは、免疫システムの異なる構成要素間で免疫情報を伝達する主要なシグナル伝達分子です。サイトカインは、免疫応答の開始、調節、維持に基づいて大まかに分類することができます。したがって、一部のサイトカインは自然免疫と炎症を媒介し、他のサイトカインは獲得免疫を調節し、残りは未成熟な免疫細胞の増殖と分化を制御します(表1.13-2を参照)。構造的には異なりますが、これらのサイトカインは機能が重複しており、協調して免疫の動的な出来事を管理します。
制御性T細胞
獲得免疫応答内では、Th1とTh2として知られる2種類のTh細胞(CD4+ T細胞)に大きな関心が寄せられています。Th1細胞は、T細胞の増殖と活性を促進し、細胞内病原体に対する防御に特に関連する炎症反応を促進するIL-2やIFN-ガンマなどのサイトカインを産生します。Th2細胞は、抗体産生を促進し、寄生虫に対する防御を提供するIL-4などのサイトカインを産生します。さらに、Th2型T制御性細胞は、直接的な細胞間効果と、特にIL-10などの抗炎症性サイトカインの産生の両方を介して、免疫活性化を抑制します。誇張されたTh2応答は、アレルギー反応、過敏症反応、および喘息と密接に関連しています。Th1細胞やTh2細胞とは発生学的に異なるTh17細胞の第3のサブセットも特定されています。これらの細胞はサイトカインIL-17を産生し、最近、かつてはTh1またはTh2の機能不全とより密接に関連していると考えられていた自己免疫疾患(乾癬)やアレルギー性疾患と関連付けられています。最近では、IL-9を分泌し、主に炎症促進性であると思われるTh9細胞と、IL-22を分泌し、炎症促進性または抗炎症性のいずれかの効果を持つ可能性があるTh22細胞を含む、追加の2つのエフェクター細胞サブセットが記述されています。
免疫と脳の相互作用
CNS(中枢神経系)と免疫系は、健康時および疾患時に相互作用し、互いを調節し合います。この複雑な免疫と脳のコミュニケーションは、図1.13-4に示されています。両システムは脅威から身を守りますが、それぞれがユニークな進化的利点を提供します。例えば、免疫系は脅威を中和するための特殊なネットワークを提供しますが、その作用領域は脅威に最も近く、即時性があります。一方、脳は優れた予測マシンとして、将来の脅威を予測し、免疫系を準備することができます。同様に、免疫系は、治癒と回復を促進する方法で行動を制御することができます。CNS内では、常在性の免疫細胞(ミクログリア)は主に実質に存在しますが、移動性の免疫細胞は、髄膜や脈絡叢を含む脳内の特殊なニッチに配置され、必要に応じて活動を開始する準備ができています。アストロサイトは、加齢や脳感染症の際に起こるように、ミクログリアによってプライミングされた後にのみ免疫応答に参加します。BBB(血液脳関門)は脳血管と脳組織を分離し、末梢免疫分子のCNSへの流入を厳密に調節します。通常の状況下では、血液とCNS間の直接的な交換は特定の場所でのみ発生します。血液と脳の交換は通常、脳室周囲器官(CVO)にある有窓毛細血管で発生します。最近発見された脳リンパ系(グリリンパ系)は、CSF(脳脊髄液)と間質液の間の液体と小分子の動きを促進します。グリリンパ系は、髄膜区画から脳へのサイトカインの経路の一つとなる可能性があります。したがって、くも膜下腔内のサイトカイン(末梢循環から)を運ぶCSFは、ビルヒョウ・ロビン腔に入り、血管周囲腔に沿って深く潜り込み、その後脳実質に拡散する可能性があります。その後、サイトカインは脳実質内のサイトカイン受容体を発現するニューロン、グリア、および間質と相互作用する可能性があります。しかし、ホメオスタシス時にサイトカインがくも膜を越えてくも膜下腔に到達するメカニズムは未解明のままです。
神経活動は、脳内の免疫応答を変化させる可能性があります。脳の常在免疫細胞であるミクログリアは、進行中の神経活動からの合図に反応します。神経活動には、シナプスから漏れ出す神経伝達物質のオーバーフローが伴います。ミクログリアは、表面受容体を用いてこれらのオーバーフローした神経伝達物質を感知します。これらの受容体の活性化は、ミクログリア細胞の機能と表現型を変化させます。さらに、脳活動は、走化性分子の放出を増加させ、BBBの透過性を変化させ、末梢循環から脳への免疫細胞の動員を促進することによって、CVOの隣接する上皮細胞を刺激することもできます。
うつ病や心的外傷後ストレス障害(PTSD)などのストレス誘発性精神疾患における免疫と脳のコミュニケーション病理の比較は、単純な説明では不十分な、より複雑な全体像を明らかにしています。単極性および双極性気分障害における末梢および中枢免疫の変化を結びつける十分な証拠があります(本章の後のセクションを参照)。新たなデータは、このプロセスがPTSDではあまり関連していない可能性を示しています。これについては、PTSDに関するセクションで議論します。
日本語に翻訳します。
ニューロン上の免疫受容体
免疫系と神経系はともに脅威を検出し、中和し、恒常性を維持します。両システムは内部および外部環境を認識し、特定の受容体を用いて周囲の刺激を監視します。例えば、自律神経系の求心性線維(迷走神経求心性神経)と脊髄後根神経節の末梢感覚ニューロンはともに**免疫受容体(TLRやサイトカイン受容体)**を示し、免疫学的シグナルを脳に伝達します。これらの免疫受容体が刺激されると、末梢ニューロンは痛みやかゆみ刺激に対してより敏感になり、行動の変化を引き起こします。例えば、免疫関連の感覚的合図によって有害とみなされた物質や状況を避けることで、生物が損傷、危険、または感染から身を守る際に、回避行動が起こります。
免疫細胞上の神経化学受容体
神経経路のシグナル伝達に加えて、末梢免疫細胞は、表1.13-3に示すように、様々な体組織によって調節される、またはそこから産生される広範囲の化学物質に対する受容体を発現します。T細胞やB細胞に発現するアドレナリン受容体は、リンパ球で最初に同定された受容体の一つでした。その後、ノルアドレナリン(NE)、ドーパミン(DA)、アセチルコリン(ACh)、セロトニン(5HT)、グルタミン酸などの様々な小分子神経伝達物質に対する受容体が、ミクログリアを含む免疫細胞上で認識されました。免疫細胞上の神経伝達物質受容体は、CNS(中枢神経系)のそれらと同様に、細胞表面に存在し、ほとんどの場合、Gタンパク質やセカンドメッセンジャー経路と結合しています。CNSの化学物質やホルモンが免疫系に与える影響は、免疫応答のどの段階(認識、活性化、またはエフェクター)が関与しているかによって異なります。例えば、ノルアドレナリンは、認識中は免疫活動を促進するが、エフェクター相では応答を抑制することが示されています。その結果、持続的なストレスはベースラインの炎症増加と関連していますが、感染症と戦う身体の能力や予防接種に対する強力な免疫応答を発達させる能力も低下させます。これらのデータは、曝露のタイミングと用量が重要であることを示唆しています。
リンパ組織の神経支配
免疫と脳のコミュニケーションの最初の兆候の一つは、免疫組織への交感神経系(SNS)の神経支配の発見でした。SNS線維は、リンパ球の成長、教育(例:自己と非自己の識別)、および機能に関与する細網内皮器官(骨髄、胸腺、脾臓、リンパ節)で発見されています。例えば、上頸神経節と星状神経節は胸腺に交感神経線維を供給し、腸間膜神経叢と腹腔神経叢は脾臓に交感神経入力を供給します。さらに、骨髄への交感神経供給は、骨の位置に対応する脊髄レベルに由来します。迷走神経または反回神経からの遠心性入力は、通常、副交感神経による拮抗調節を提供します。交感神経線維は通常、血管供給と並行してリンパ組織に入ります。これらの線維は、動脈に沿って並ぶ平滑筋の収縮性を制御することにより、血管緊張を調節することで広く知られています。それにもかかわらず、浸潤する神経線維は、発達中のリンパ球やマクロファージの近くにも終末を形成します。血流をリンパ器官に変化させたり、NEや神経ペプチドなどの神経伝達物質を局所的に放出したりすることで、SNS線維は免疫細胞の機能を調節することができます。
