
- 1.28 機能的脳結合性と精神病理
- 図 1.28-1 機能的結合性ネットワークと精神病理
- 安静時機能的結合性を評価するためのモダリティ
- 図 1.28-2. 機能的結合性解析を特徴付ける次元
- 安静時機能的結合性の解析方法
- ROI-ROIまたはネットワークベースの結合性
- ROI/シードベースの結合性
- 独立成分分析 (Independent Components Analysis)
- 関連性 vs. 方向性 (Association versus Directionality)
- 静的評価 vs. 動的評価 (Static versus Dynamic Assessments)
- 機能的結合性に対するネットワークアプローチ (Network Approaches to Functional Connectivity)
- 安静時機能的結合性分析を精神病理の研究に適用する上での利点と課題 (Benefits and Challenges in Applying Resting-State Functional Connectivity Analyses to the Study of Psychopathology)
- 精神病理における安静時機能的結合性研究からの洞察
- 図1.28-3. 統合失調症における視床結合性の変化パターン
- 図1.28-4. 扁桃体結合性のパターン
- 図1.28-5. 扁桃体から内側前頭前野への結合性と子どもの感情調節の関係
- 結論
- 参考文献
1.28 機能的脳結合性と精神病理
ディーナ・M・バーチ 博士
1950年代に現代の「生物学的精神医学」が出現して以来、精神疾患の病因を理解するための多くの研究は、さまざまな精神病理の発症と維持に寄与する神経メカニズムの特定に焦点を当ててきました。この研究は、ヒトの脳の構造的および機能的特性を評価するためのSPECT、PET、MRIなどの新しい神経画像技術の進歩によって支援されてきました。本章では、ヒトの脳の機能的結合性の測定の利用と、脳結合性の理解がさまざまな精神病理の発症と維持につながるメカニズムの一般的な理解にどのように役立つかに焦点を当てます。本章は、ヒトの機能的脳結合性を測定し理解するための方法の開発の短い歴史から始まります。次に、本章では、現在利用可能な機能的脳結合性を研究するための方法の簡単な概要と、それらの相対的な長所と短所の議論を提供します。ヒトの「内因性」機能的脳結合性ネットワークに関する現在の知識の概要も提供されます。最後に、本章では、精神疾患の神経生物学を理解するために機能的脳結合性を用いた研究の一部を選択的に記述します。特に、精神病と抑うつに焦点を当てます。これらは、機能的結合性障害に関する既存の文献が最も多い精神病理の2つの側面であるためです。
脳機能の観点から見ると、歴史的に、精神病理における機能的神経画像研究の多くは、SPECT/PETまたはfMRIのいずれかの使用に焦点を当ててきました。これらのアプローチは、グルコースまたは酸素代謝(PET)、血流(SPECT)、または血液酸素レベル依存性(BOLD)活動の測定を通じて、神経活動の間接的な推定値を提供します。精神病理の神経メカニズムに関する疑問に対処する最初の研究では、参加者がさまざまな認知、感情、または社会的なタスクを実行している間の脳領域の差動活性を評価するために、主にタスクベースのPETまたはfMRIが使用されました。これらの研究は、認知または感情タスクの実行中の異常なBOLD活動が異常な基礎となる神経プロセスを示すという推論の下で、精神疾患のさまざまな症状でどの脳領域が影響を受ける可能性があるかについての仮説の発展と検証に不可欠でした。いくつかの例として、この研究は、精神病における認知機能障害における背外側前頭前野(DLPFC)の役割、抑うつにおける扁桃体の役割、少なくとも一部の不安障害における扁桃体と島皮質の役割、および依存症における線条体メカニズムの役割に関する理論の発展につながりました。
タスク関連の脳活動に関する基礎的および翻訳研究は、特定の認知および効果的プロセスの神経基盤に関する現在の理解と、これらが精神病理においてどのように異常になるかという点で、非常に有益でした。しかし、時間が経つにつれて、特定のタスクが単一の脳領域のみを活性化することはまれであり、特定の精神病理のドメインが単一の脳領域のみにおける異常な活性化と関連することはまれであることが明らかになりました。さらに、基礎神経科学研究は、個々の脳領域(または個々のニューロン!)における活動は、脳の異なる領域からの入力と出力の結果であることを以前から明らかにしています。このような認識は、特定の脳領域のみに焦点を当てるのではなく、神経回路に関する疑問に焦点を移すことにつながりました。より具体的には、このシフトは、健康における成功した認知および感情機能を生み出す上での異なる脳領域間の関係、および回路レベルでの異常が精神病理の特定の側面の発症と維持にどのように寄与するかという疑問へと移行しました。
統合失調症における認知機能障害におけるDLPFCの役割に関する疑問は、この点で優れたケーススタディを提供します。精神病における認知障害に関連するDLPFC機能障害に関する初期の研究の多くは、ほとんどDLPFC自体にのみ焦点を当てていました。しかし、例えば、認知制御タスク中にDLPFCで観察された異常な活動が、実際には他の脳領域からの誤った入力の結果であった場合はどうでしょうか?言い換えれば、統合失調症においてDLPFC自体は正常に機能していたが、それが受け取った情報が観察された異常につながっていた場合はどうでしょうか?あるいは、問題が特定の脳領域内の活動ではなく、脳領域間の接続または活動を調整する能力にあった場合はどうでしょうか?神経ネットワーク間および内部の活動を調整する上での脳振動の役割に関する研究の増加は、精神病理における障害を特定の個々の脳領域内ではなく、機能の回路レベルに局在させるという仮説と一致しています。さらに、多くの点で、回路レベルの仮説を、精神病理における他の分析レベルでの障害に関するより大規模な文献(例:神経伝達物質の利用可能性、受容体、E-Iバランス)と結びつけることはより容易です。このような障害は、しばしば領域特異的ではなく、皮質領域内または皮質領域間、あるいは皮質と皮質下領域間の領域間コミュニケーションを合理的に変化させると予想されるプロセスとメカニズムを伴います。
機能的結合性の研究の出現
これらの種類の疑問を検証する能力は、ヒトにおける機能的結合性手法の開発によって推進されてきました。元来、機能的結合性の概念は、非侵襲的神経画像法を用いてヒトで観察される機能的結合性に寄与すると考えられる、ニューロンのスパイク列の同時記録に適用されていました。機能的結合性の主な推論は、2つの領域が高度に相関するニューロン活動(すなわち、高い機能的結合性を持つ)を持つ場合、それらが共通の処理メカニズムに関与する可能性が高いということです。もしそうであれば、機能的結合性は、認知または感情的な要求の完了中にどの脳領域が通信しているかを理解するためのツールを提供し、したがって、どのような脳回路が認知、感情、および/または社会的処理の異なる領域におけるパフォーマンスと能力をサポートしているかを理解するためのツールとなります。
安静時機能的結合性
ヒトにおける機能的結合性手法の開発と応用における大きなブレークスルーは、1995年にビスワルらが、右および左の運動皮質の領域からの自発的活動が、個人が安静にしていても高く相関していることを報告し、個人がタスクを実行していなくてもこれらの脳領域間に機能的結合性を示唆したときに起こりました。興味深いことに、この相関活動は、BOLD信号の自発的な低周波変動(0.01〜0.10 Hz)で観察されました。この周波数帯域は、タスクベースの研究ではノイズとして捨てられることが多かったものの、このような相関は他の周波数でも見られます。この研究は、現在安静時機能的結合性と呼ばれる主要な探求分野を刺激しました。重要なことに、この安静時の脳の状態の活動は、脳が身体の総質量のわずか2%であるにもかかわらず、身体のエネルギーの主要な部分(約20%)を消費すると考えられています。さらに、特定のタスクへの関与による代謝の変化は通常5%未満であり、これは、進行中の安静時活動が、タスクパフォーマンスによって観察される変化よりも、病気関連の変動性の豊かさにおいて、より豊かな、あるいはそれ以上の情報源を提供する可能性があることを示唆しています。さらに、タスク関連活動の試行ごとの変動性の大部分が、脳活動における一貫性のある組織化された自発的変動を反映していることを示す確固たる研究もあり、これがヒトの脳機能における意味のある変動源であるという別の証拠を提供しています。
安静時機能的結合性ネットワーク
この脳機能の側面への関心を促した安静時機能的結合性の主要な側面の1つは、安静時機能的結合性が、進行中の脳活動におけるタスク誘発性の摂動がない場合でも、一貫して機能的に結合している脳領域の本質的に組織化されたネットワークを明らかにしているという認識です。これは、安静時に協調的な活動を示す脳領域の一貫した、堅牢で、再現性のあるネットワークを特定した数多くの「ネットワーク」マッピング研究によって裏付けられています。これらには、デフォルトモードネットワーク(DMN)、前頭頂ネットワーク、帯状蓋ネットワーク、背側注意ネットワークなどが含まれます。これの最近の例の1つは図1.28-1に示されており、安静時機能的結合性の非常に均一なパターンを持つと特定された356の領域(「区画」)が、以前の安静時研究で特定されたネットワークにうまくマッピングされる12の安静時ネットワークにグループ化されました。このようなネットワークは、機能的結合性を評価するためのさまざまな方法間で頑健に特定され、個人間で頑健に特定されますが、特定の領域のサイズと位置には重要な個人差を示し、少なくとも部分的に出生後の早い時期に発生します。重要なことに、これらのネットワークの多くは、タスクベースの研究で特定されたネットワークと密接に、または少なくとも部分的にマッピングされており、この協調活動の機能的意味合いを検証しています。
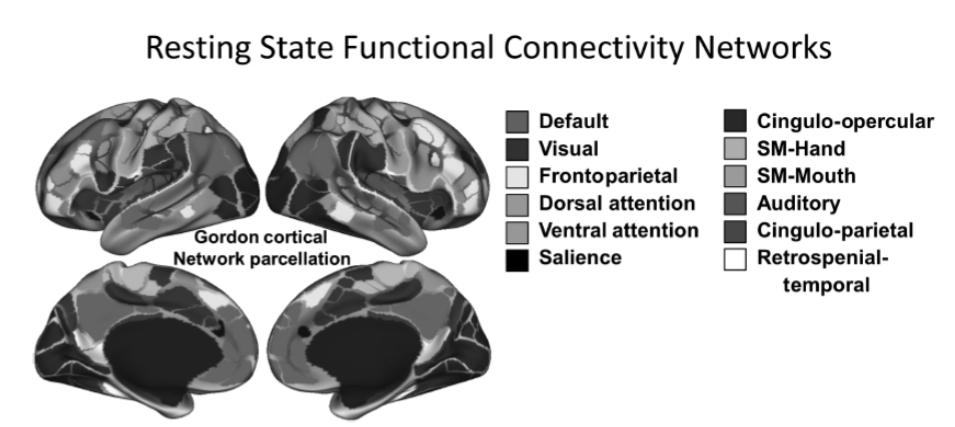
図 1.28-1 機能的結合性ネットワークと精神病理
図 1.28-1は、安静時データから導き出された機能的結合性ネットワークを示しています。上段は脳の側面図、下段は脳の内側面図です。色の付いた「パーセル(小区分)」は、全脳の機能的結合性マップにおける勾配の変化の境界を特定するために、「洪水法による分水嶺」アルゴリズムを用いた境界特定アプローチから導き出されました。この境界マッピングから導き出されたパーセルのコミュニティ構造は、InfoMapアルゴリズムを用いて生成されました。(カラー画像については電子書籍を参照)。(Gordon EM, Laumann TO, Adeyemo B, Huckins JF, et al. Generation and evaluation of a cortical area parcellation from resting-state correlations. Cereb Cortex. 2016;26(1):288-303, Oxford Universityの許可を得て転載。doi: 0.1093/cercor/bhu239.)
このような機能的結合性ネットワークは、部分的に構造的結合性に基づいていますが、直接的な構造的結合性を示す領域とは同形ではありません。そのため、安静時ネットワークと既知のタスクネットワークのマッピングは、安静時ネットワークが、部分的に、発生時の共活性化の履歴から生じる組織化された相互作用を反映しているという仮説につながっています。ただし、前述のように、この組織の主要な要素は出生時にも存在します。このようなネットワークは、特定のタスクの実行に依存せず、安静時にも存在し、タスクや環境の状態に比較的安定していることから、現在では「内在性」結合性ネットワークと頻繁に呼ばれています。
複数のスケール、モダリティ、年齢にわたるヒトの機能的および構造的コネクトームを研究する最近の大規模プロジェクトである「ライフスパン全体にわたるヒトコネクトームプロジェクト」は、ヒトの脳におけるこれらのコアネットワークの存在を裏付けています。さらに、ヒトコネクトームプロジェクトの結果は、機能的脳結合性の個人差と、精神的健康、およびライフスパン全体にわたる負および正の生活結果に関連する幅広い行動の個人差との間の重要な関係に関する現在の知識を確認し、拡張しました。
原則として、あらゆるヒトの脳ネットワークは精神病理の理解に関連しますが、最も一般的に特定されるいくつかのネットワークは、精神病理でしばしば障害が見られる機能やプロセスに特に関連しています。例えば、前頭頭頂ネットワークと帯状蓋ネットワークは、様々な認知制御機能と一貫して関連付けられています。前頭頭頂ネットワークには、外側前頭前野と頭頂皮質の両方の背側領域が含まれ、帯状蓋ネットワークには、背側前帯状回(dACC)、両側の背側前島皮質、そして時には視床領域と前前頭前野領域の両方が含まれます。後述するように、これらのネットワークは、統合失調症などの認知制御に既知の障害がある集団や、大うつ病などの感情調節に障害がある集団で異常であることが判明しています。
背側および腹側注意ネットワークも認知機能と関連付けられていますが、トップダウンおよび/または刺激駆動型注意機能とより密接に関連しています。背側注意ネットワークは、背側、前頭、頭頂領域だけでなく、より背側の補足運動野および眼野領域も含む点で、前頭頭頂ネットワークと密接に連携しています。腹側注意システムには、側頭頭頂接合部と腹外側前頭前野(VLPFC)が含まれ、環境中の顕著な出来事への注意と関連付けられており、進行中の処理を妨げる出来事が世界で発生したときにしばしば活性化されます。不安障害における脅威関連刺激への注意の変容に寄与する可能性があるという仮説の下で、結合性の変化が不安障害と関連付けられています。
DMNは様々な機能と関連付けられていますが、特に、内部の感情状態への注意と、内部と外部の注意モードを区別または切り替える能力と関連している可能性があります。さらに、DMNがタスク実行中に活動を減少させることを示す多くの文献があり、DMNの「シャットダウン」が失敗すると認知機能が損なわれる可能性があるという示唆もあります。DMN(図1.28-1を参照)には幅広い領域が含まれますが、最も顕著なのは内側前頭前野、内側後帯状回、および楔前部領域です。さらに、多くの研究は、複数のDMNサブネットワークが存在し、それぞれがやや異なる機能を果たす可能性があることを示唆しています。DMN領域間の結合性の増加は、大うつ病における反芻と関連付けられています。
サリエンスネットワークは、他のいくつかのネットワークよりもやや最近特定されたもので、帯状蓋ネットワークと通常関連付けられるよりも吻側の前帯状回および島皮質領域が含まれます(図1.28-1を参照)。ただし、両者とも辺縁系および皮質下領域との強い結合性を持っています。サリエンスネットワークの結合性は、不安と覚醒と関連付けられており、環境中の顕著な出来事に対する反応を処理および調整する役割を果たす領域のネットワークであると仮説が立てられています。
安静時機能的結合性を評価するためのモダリティ
ヒトの安静時機能的結合性は、様々な方法を用いて評価することができます。圧倒的に最も一般的なのは、BOLD関連活動の自発的変動に焦点を当てたfMRIの使用です。このアプローチは非侵襲的であり、使用するパルスシーケンスによって良好から優れた空間解像度を提供しますが、BOLD関連信号が神経活動の間接的な尺度である(すなわち、局所的な磁気均一性を変化させる神経活動に対する代償的な血流反応を反映している)ことと、BOLD信号が遅く、時間的に伸びているという事実に本質的に限定されます。
安静時機能的結合性は、EEGまたは脳磁図(MEG)を用いて測定することもできます。これらは、はるかにきめ細かい時間解像度と異なる周波数で結合性を解像する能力を提供しますが、これらの方法は信号を局所化する能力(すなわちEEG)や、線条体や視床などの領域の活動の深いソースに対処する能力(すなわちMEG)に限界がある場合があります。機能的結合性は、硬膜下に電極グリッドを配置するなどの、より侵襲的な技術である電気皮質記録(ECoG)を用いて評価することもできます。このようなアプローチは、神経活動によって生成される電気活動に直接マッピングできるという明白な利点と、良好な空間局所化を有しますが、医学的理由で頭蓋内記録を必要とする集団での使用に限定されます。
さらに、機能的結合性に関する研究の多くは、参加者が目を閉じるか開けるかして静かに休息している間に取得されたデータに焦点を当てていますが、現在では、特定のタスクの実行中や映画の視聴中に取得されたデータからも、いくつかの注意点はあるものの、非常に類似した結果が得られることが多数の研究で示されています。そのようなアプローチの1つは、タスクの実行中に取得されたデータを使用し、回帰分析によってタスク構造のBOLD活動への決定論的な影響を取り除き、その後、回帰の残差に基づいて機能的結合性分析を行うことです。このような研究の結果は、従来の安静時研究の結果と非常に類似している可能性があります。ただし、これがどの程度当てはまるかは、データが統合される異なるタスク状態の数に依存する可能性があります。単一のタスク状態でのみこれを行うことは、本質的に、あるタスク状態と別のタスク状態における結合性の違いを比較することに焦点を当てた精神生理学的相互作用分析アプローチ(PPI)を用いて生成されたデータと機能的に同等です。さらに、先行する「状態」や特定の種類のタスクの実行が、その後の安静時結合性の評価に影響を与える可能性があるという文献上のいくつかの証拠があります。そのため、いくつかのタスク状態を統合することは、特定のタスク状態の決定論的または残差効果を排除し、したがって一般化可能性を高め、安静時結合性と類似性を高めるのに効果的である可能性があります。人気が高まっているもう1つのアプローチは、人々が映画を見ている間の機能的結合性を調べることです。これにより、行動予測において安静時データと同じくらい良好なパフォーマンスを示すという証拠があります。当然のことながら、このようなアプローチは、子供のように静かに休息することが困難な特別な集団にとって特に魅力的です。

図 1.28-2. 機能的結合性解析を特徴付ける次元
下段は、機能的結合性データの解析に対する「仮説駆動型」から「データ駆動型」への次元を示しています。中段は、結合性の関連性を特徴付けることに焦点を当てた解析アプローチと、因果関係または方向性関係を特定しようとするアプローチの次元を示しています。上段は、単一セッションにおける結合性の静的推定に焦点を当てた解析アプローチと、セッション内またはセッション間の変化のダイナミクスに焦点を当てたアプローチの次元を示しています。
安静時機能的結合性の解析方法
ほとんどの神経画像法と同様に、安静時機能的結合性データの使用は、研究者にとって多くの選択肢をもたらします。主に、機能的結合性をどのように計算するか、異なる脳領域をどのように識別するか、ノイズをどのように処理するかという点でです。これらの選択肢はそれぞれデータの構造と品質に影響を与えますが、すべての安静時研究に最適な「最良の」方法についての明確なコンセンサスはなく、文献には一般的に使用されるアプローチの傾向があるだけです。これらの問題に関連するすべての可能なバリエーションや選択肢のレビューは本章の範囲を超えており、他でレビューされています。本章では、精神病理に関する現在の文献を理解する上で最も関連性の高い選択肢に焦点を当てます。
精神病理に関するほとんどの研究では、安静時機能的結合性解析の主な目標は、特定の、ア・プリオリな領域とその結合性への関心から、または機能的ネットワークの障害をよりよく理解するために、異なる脳領域間の関係の変動を評価することです。そのため、特性分析アプローチの1つの次元(図1.28-2を参照)は、比較的ア・プリオリで仮説駆動型から、比較的データ駆動型へと移行します。この次元に沿って、最も一般的な3つの分析手法は、関心領域(ROI)またはア・プリオリなネットワークベースの分析、シード/ROIから全脳結合性分析、および独立成分分析(ICA)またはその他のクラスタリング方法です。これら3つの方法にはそれぞれ長所と短所があり、手元の研究課題を最も正確にテストできる分析を決定するのは研究者次第です。
ROI-ROIまたはネットワークベースの結合性
ROIからROI、またはネットワークベースの分析は、通常、文献で既に特定され特徴付けられている特定の領域のセットまたは特定の領域のネットワークを出発点とし、これらの領域から時系列を抽出し、それらの特定のROI間またはその特定のネットワーク内の領域間の接続を分析することに焦点を当てます。例えば、特定の精神病理の次元が、2つの特定の領域(例:DMNの内側前頭前野と楔前部領域)間の異常な結合性、またはDMNに関連するすべての領域または一部の領域間の異常な結合性と関連しているという仮説をテストしたいと考えるかもしれません。例えば、先行研究では、前頭頭頂ネットワーク、帯状蓋ネットワーク、またはデフォルトモードネットワーク内の領域間または領域間の結合性が統合失調症で変化するかどうか、そしてそれが認知機能の変動と関連するかどうかが検討されています。別の例として、Perinoらは、背側および腹側注意システム間の結合性が若年者の不安を予測するかどうかを検討しました。注目すべきは、最近の研究が、これらのネットワーク内の領域のサイズと位置に重要な個人差がある可能性があり、十分なデータがあれば、個人特有の領域とネットワークを確実に評価できることを示唆していることです。このアプローチは、行動と精神病理への関連性を高める可能性があります。
ROI/シードベースの結合性
もう1つの一般的なアプローチは、特定のROIまたは「シード」から開始し、シードと脳の残りの部分、または少なくとも脳の一部との間の結合性を調べます。シードベースの結合性は柔軟な方法であり、望むならば実行できますが、特定の関心領域の機能的結合性マップを理解するためにより一般的に使用されます。文献にはこれの多くの例があり、例えば、うつ病に関連する扁桃体、または扁桃体のサブ領域の結合性を調べた研究などです。シード領域は、シード領域が特定のネットワークの堅牢なメンバーであると仮定して、機能的ネットワークを識別するためにも使用できます。しかし、多くの領域が例えばDLPFCと強く結合しているからといって、それらの領域が単一のネットワークを形成していると necessarily 示すわけではありません。シード由来の機能的ネットワークの場合、ほとんどの他のノードが互いに、そしてシードとも機能的に結合していることを確認することが重要です。
まとめると、ROI-ROI、ネットワーク、またはシードベースの結合性方法の最大の利点の1つは、特定の研究課題を直接テストできることであり、そのような研究における発見の解釈を比較的簡単にする点です。ただし、あらゆる機能的結合性研究で生じる潜在的な解釈上の注意点は除きます。
しかし、重要なことに、ROIまたはシードに焦点を当てた研究のこの利点は、そのような研究のいくつかの欠点にも寄与する可能性があります。ROI/シードベースの研究は、研究者が脳の特定の領域またはネットワーク間の関係について具体的な予測を立て、それらの疑問を直接テストすることを可能にします。しかし、すべての他の相関のシードとして解剖学的領域を事前選択することで、研究者はデータにバイアスを導入する可能性があります。すべての被験者において、関心のある機能領域に最も正確で信頼性の高い時系列を提供するための完璧な方法はありません。2つの異なるネットワークの機能領域が脳内で隣接している可能性を考えると、誤った領域からのいくつかのボクセルの包含が、2つの異なる機能ネットワークを反映する結合性マップにつながる可能性があります。それに加えて、少なくとも一部の臨床集団は解剖学的異常を持っている可能性があり、領域の識別を困難にする可能性があるという考慮事項もあります。しかし、前述のように、機能的結合性のパターンに基づいて個人特有の脳領域を確実に識別できるという証拠が増えており、これによりこれらの問題の一部に対処できます。さらに、シードまたはROIに焦点を当てた研究は、「街灯の下」問題に悩まされる可能性もあります。これは、研究者がア・プリオリな領域で有意な結果を見つけることに偏っており、単にそれらの領域が調査されたためであり、研究者は異なる一連の領域を使用すれば、より強力またはより一貫した結果が得られたかどうかを知らないということです。
シードベースの結合性アプローチのもう1つの限界は、それらが実際には多変数システムであるものに対して、単変数分析を提供するということです。このようなROIまたはシードベースの結合性分析では、1つ以上の領域の時系列が、1つ以上の他の領域またはボクセルの時系列と相関付けられ、ペアワイズ関係を理解しようとします。このプロセスで無視されているのは、観察されたペアワイズ関係につながる複雑な相互作用であり、例えば、他の接続によるそのペアの領域またはボクセルの結合性の変調などです。したがって、シードベースの結合性分析は、前述のように機能的ネットワークの解剖学的定義を制限するだけでなく、これらの機能的ネットワークの解釈は、ネットワークが埋め込まれているシステムの完全な複雑さを考慮に入れていません。
独立成分分析 (Independent Components Analysis)
ROIまたはシードベースの結合性解析の一般的な代替手段として、独立成分分析(ICA)があります。これは、脳の機能ネットワークを特定するための多変量データ駆動型アプローチです。ICAは、BOLDデータの全体を空間的に分離された、最大限に統計的に独立した成分に分解するアルゴリズムです。これにより、ICAは、ROI/シードベースの結合性に関連する上記の制限のいくつかに対応します。具体的には、ICAはア・プリオリな領域の定義を必要とせず、最適ではないボクセルクラスターを選択したり、特定の接続のみを調べたりする潜在的なバイアスを解決します。また、ICAによって特定された成分は、すべてのボクセルからの時系列に関する情報を含む完全なBOLDデータセットを使用して決定されるため、多変量データの複雑さも考慮に入れます。
当然のことながら、ICAもシードベースの結合性と同様に、データを偏らせる可能性のあるいくつかの制限を伴います。最も顕著なのは、ICAはデータの成分を識別しますが、各成分が何を表しているかを決定するのは研究者次第であるということです。例えば、研究者は、どの成分がノイズであり、どの成分が機能ネットワークを表すかを決定する必要があります。ただし、以前に検証されたノイズ成分の自動識別など、研究者がこれを達成するのに役立つ戦略は存在します。最後に、ICAデータの解釈は、シードベースの結果の解釈ほど常に単純ではありません。具体的には、ICAの尺度は成分の大きさであり、特定のペアワイズ関係の大きさではありません。成分の大きさは、空間分布の変動、信号の変動の減少、または領域間の時系列の相関の減少など、さまざまな理由で減少する可能性があり、これらのうちどれが結果を左右しているのかが常に明確であるとは限りません。
関連性 vs. 方向性 (Association versus Directionality)
安静時データの分析が変化するもう1つの次元(図1.28-2を参照)は、単に関連性を特徴付けることに焦点を当てるか、または因果的または方向性のある関連性を特定しようとするかという程度です。安静時データの分析において最も一般的なアプローチは、相関またはICAを単純に使用することです。これらは関連性に方向性を前提とせず、領域間の直接的な接続と他の領域との共有分散の両方による共分散を捉えます。スペクトルの反対の端には、動的因果モデリング、グレンジャー因果性、およびラグ付き時系列分析などのアプローチがあり、これらは共分散のパターンから時間的または因果的な方向性を確立しようとしますが、安静時fMRIデータでこの方法を使用することには課題があります。これら2つの極端の中間にあるのは、部分相関の使用であり、これは、領域またはボクセル間の結合性の直接的な関連性を分離し、他の領域との共有接続による共分散を部分的に排除しようとします。
静的評価 vs. 動的評価 (Static versus Dynamic Assessments)
安静時データの分析が変化するもう1つの次元は、ある時点での個人の機能的結合性パターンの静的な推定に焦点を当てるか、または時間経過に伴う結合性「状態」の動的な変動に焦点を当てるかです。現在、多くの研究が、脳領域間の結合性パターンが時間とともにどのように変化し、限られた結合性「状態」または結合性パターンに分類されるかを検討し始めています。これらの状態パターンは、臨床集団によって、達成される状態の数、異なる状態で費やされる時間、または状態間の移行の速度の点で異なる場合があります。
機能的結合性に対するネットワークアプローチ (Network Approaches to Functional Connectivity)
シード/ROIベースおよびデータ駆動型アプローチを補完するものとして、研究者はネットワークサイエンスのツールを、主にグラフ理論を用いて脳の組織を理解するためにますます適用しています。グラフ理論アプローチを採用する場合、研究者は、グローバル効率、ローカル効率、スモールワールドネス、参加係数など、いくつかのグラフ特性の変動を調べています。グローバル効率は、ネットワーク内の脳領域間(例:DMN内の領域間)で情報がどれだけ容易に伝達されるかを指標化することにより、脳ネットワークの機能統合を測定します。ローカル効率は、脳領域の隣接領域がその領域(グラフ理論用語で「ノード」)が除去された後もどの程度互いに通信し続けるかを測定し、したがって、ネットワークのフォールトトレランスのようなものを表します。参加係数は、特定の脳領域が通信する異なるネットワークの数を示し、全体的な脳の通信においてその領域がどれほど重要であるかについての情報を提供します。参加係数の高い領域はしばしば「ハブ」と呼ばれ、情報伝達に特に重要であると考えられています。ヒトの脳で特定されているハブには、DLPFC、楔前部、島皮質などがあり、特に互いに強く結合しているものは「リッチクラブ」と呼ばれます。グラフ理論アプローチは、多くの種類の精神病理における脳組織を調べるために使用されてきましたが、おそらく最も頻繁に統合失調症において使用されており、これについては以下で詳しく説明します。
安静時機能的結合性分析を精神病理の研究に適用する上での利点と課題 (Benefits and Challenges in Applying Resting-State Functional Connectivity Analyses to the Study of Psychopathology)
安静時機能的結合性に関する研究は、ヒトの脳活動と組織が理解される方法を形作っただけでなく、臨床集団の研究にも重要な影響を与えてきました。精神疾患を持つ個人においてタスクベースのデータよりも安静時データを検討することの潜在的な利点の1つは、タスクベースの画像データほどタスクパフォーマンスによって複雑にならない可能性があることです。タスクベースの画像データは、健康な個人と様々な患者集団を比較する研究において困難な交絡因子であることが証明されています。
主に、タスクベースの研究の懸念は、コントロールと患者の間で観察されるBOLD活動の違いが、タスクの完了能力の違いによるものなのか、それとも脳領域を使用する固有の問題によるものなのかということです。なぜなら、タスクの難易度の調整は、被験者内のBOLD活動に影響を与えることが多いからです。このため、多くの研究者は、健康なコントロールではBOLD信号とパフォーマンスに変動を生み出すのに十分な難しさでありながら、患者が確実に学習し実行できる十分な容易さのタスクを設計するという困難な状況に直面してきました。さらに、このタスクパフォーマンスの交絡因子は、スキャナー内で認知または効果的なタスクを許容できる高機能患者に焦点を当てる傾向のある研究者の募集戦略を形作ってきました。このため、統合失調症におけるタスクベースのfMRI研究の大部分は、高機能患者の集団の使用によって偏っている可能性があり、したがって、より症状のある個人には常に一般化できるとは限りません。したがって、安静時機能的結合性は、統合失調症を持つより広範な個人の神経生物学的異常を評価する機会を提供する可能性があります。
しかし、安静時研究には、精神病理の神経メカニズムを理解するためにそのようなデータを使用する際に、独自の特定の課題が伴うことも心に留めておくことが重要です。最初にして最も重要な課題は、参加者の動きによって導入される有害な交絡因子であり、これは臨床集団でしばしば大きくなります。動きは、あらゆる種類のアーティファクトと交絡因子を安静時データに導入する可能性があり、処理中に適切に対処されない場合、臨床集団における結合性変化の性質と原因について誤った結論につながる可能性があります。例えば、動きの効果の1つは、近くにある領域間の接続の大きさを増加させる一方で、より離れた領域間の接続の強さを減少させることです。これは距離依存性運動アーティファクトと呼ばれる交絡因子です。このようなアーティファクトの存在は、データで動きの交絡因子が十分に考慮されていない場合に、臨床集団における長距離接続の全体的な減少と局所接続の増加を発見したと主張する研究の解釈を困難にします。
もう1つの潜在的な課題は、覚醒度/眠気、呼吸数、開眼/閉眼状態などの要因の変動であり、これらもすべて、安静時機能的結合性パターンをある程度変調させる可能性があります。さらに、長期研究でそのような尺度を使用したい場合、例えば、病気への移行、治療反応、または時間の経過に伴う長期的な経過や変化を予測しようとする研究では、追加の課題があります。測定の信頼性の低さから実際の変化を区別できる必要があります。この後者の懸念は、機能的結合性だけでなく、あらゆる神経画像測定に関連するものです。
精神病理における安静時機能的結合性研究からの洞察
現在までに、精神病理の様々な側面を持つ様々な集団で、数千もの安静時機能的結合性研究が実施されています。そのため、これらの研究すべてをレビューすることは本章の範囲をはるかに超えています。しかし、精神病理を研究するために安静時機能的結合性を使用することで得られた洞察の一部を示すために、本章では、安静時研究の文献が最も多いと議論されている2つの領域、すなわち統合失調症と大うつ病に焦点を当てた研究を選択的にレビューします。具体的な焦点は、研究間で再現され、将来の研究の明確な経路または方向性を示す結果の収束が始まりつつある領域を特定することです。
統合失調症スペクトラム障害における機能的結合性の異常
現在までに、統合失調症スペクトラム障害における機能的結合性の様々な側面を検討した500以上の実証研究があり、その大部分は、精神疾患の診断・統計マニュアル第4版(DSM-IV)または第5版(DSM-5)によって統合失調症と診断された個人を対象に行われています。これらの研究は、様々な方法とアプローチを用いており、そのため、非常に多様な知見を生み出しています。しかし、統合失調症患者における機能的結合性の変化の明確で説得力のある側面を示す、ますます一貫性のある結果が得られている領域があります。
視床結合性の変化。
統合失調症における機能的結合性への強い焦点は、統合失調症が「切断症候群」であるという長年の仮説と一致しています。これは、複数の脳領域にわたる協調行動の失敗によって認知機能と行動に障害が生じるというものです。多くの研究者が指摘しているように、この仮説のいくつかのバージョンは、ヴェルニッケやブロイラーの研究の初期にすでに提唱されていました。さらに、脳領域間の接続の異常に関する理論も、統合失調症の病態生理に関するより現代的な理論において中心的な役割を果たしてきました。アンドレアセンらが提唱したそのような著名な理論の1つは、統合失調症が皮質-皮質下-小脳回路の統合の障害を伴うと示唆しており、この仮説は「認知失調」と名付けられました。視床の構造と機能の変化は、統合失調症の文献で顕著です。
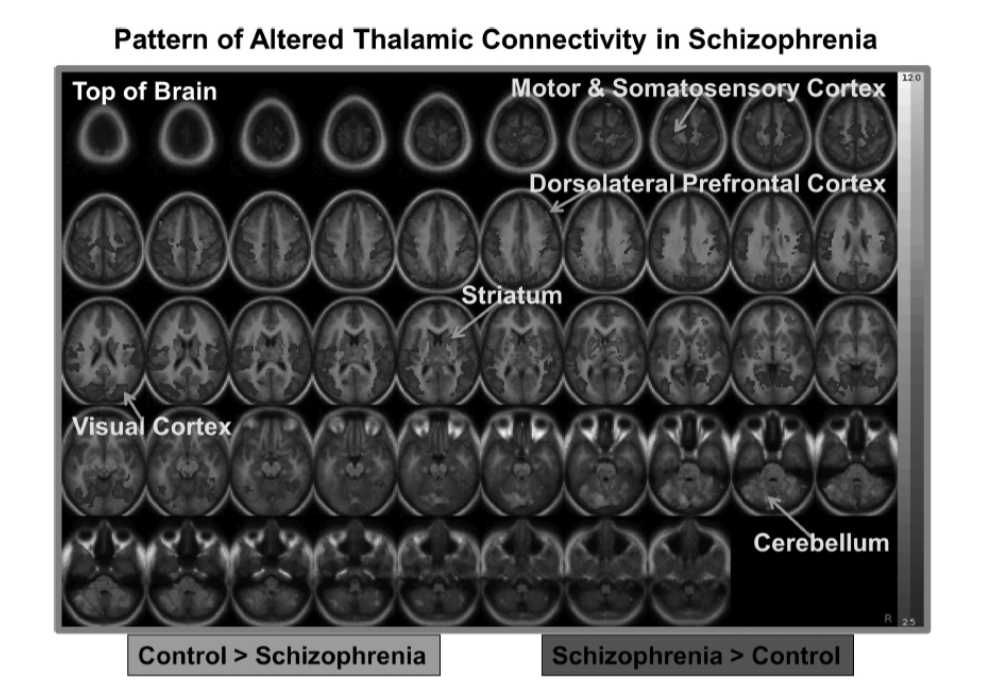
図1.28-3. 統合失調症における視床結合性の変化パターン
未発表データ。健常者(赤)よりも統合失調症患者で視床結合性が低い領域、および統合失調症患者(青)で健常者よりも視床結合性が高い領域を示す。統合失調症で視床結合性の低下を示す領域には、小脳、線条体、背外側前頭前野、背側前帯状回が含まれる。統合失調症患者で視床結合性の増加を示す領域には、運動皮質と体性感覚皮質、視覚皮質、および前側頭極が含まれる。(カラー画像については電子書籍を参照。)
霊長類の解剖学的研究で示されているように、視床は特定の視床核と皮質脳領域の異なる領域を結ぶ並列経路に地形学的に組織化されており、皮質下領域と皮質領域間の様々な種類の情報処理のための並列ループの形成を助けています。特に、視床の内側背側核および前核はDLPFCに投射しています。現在、多くの確固たる証拠が、両側の視床領域、特に内側背側核および前核から、両側のDLPFC、dACC、線条体の一部、および両側の小脳への結合性を示しています。多くの研究では、これらの結合性の低下パターンは、視床間の結合性の増加と関連付けられています。これらの結合性の変化パターンは図1.28-3に示されており、統合失調症で視床結合性の低下を示す領域が青色で、結合性の増加を示す領域が赤色で示されています。公開されている文献と同様に、このサンプルにおける統合失調症患者は、両側の前頭前野および頭頂皮質、ACC、ならびに両側の線条体および小脳との視床結合性の低下を示しています。対照的に、彼らは運動皮質、体性感覚皮質、および視覚皮質との両側性結合性の増加を示しています。
同様の結合性の変化パターンは、統合失調症の臨床的および遺伝的ハイリスクの個人、さらには双極性障害など、精神病を特徴とする他の疾患でもある程度見られます。さらに、高臨床リスクの個人では、視床結合性の変化パターンは、その後本格的な精神病診断に移行した人々の間で最も顕著でした。加えて、構造的結合性の研究も、視床(特に内側背側)から前頭前野への結合性の変化、内包前肢の異方性度(fractional anisotropy)の低下を含めて支持し始めています。重要なことに、視床とDLPFCの間の結合性の低下は、認知機能障害を予測します。視床結合性の変化に関する研究全体にわたるこれらの堅牢で一貫した発見は、統合失調症スペクトラム障害の個人ではこれらの脳領域間の協調が障害されているという考えと完全に一致しています。さらに、この障害された視床結合性と認知機能との関係は、視床皮質ネットワークが効果的な認知機能に重要な脳振動を組織化する上で不可欠であり、分析の他のレベル(例:E-Iバランス)での障害によるそのような回路の混乱が大規模な脳機能を著しく損なう可能性があるという考えと一致しています。
視床結合性に関する結果の様々な側面は、それが精神病リスクの潜在的なバイオマーカーとして主要な重みを与えられる必要があることを示唆しています。第一に、そのような結果が、投薬を受けたことのない個人でも発見されているという事実です。第二に、それが疾患の経過全体にわたって存在し、臨床症状の発現または遺伝的リスクのいずれかによって精神病のリスクを共有する人々にも存在するという事実です。したがって、次の重要なステップは、視床結合性を、精神病の発症の他の早期指標またはリスクマーカーと関連付けることです。そのようなマーカーには、線条体におけるシナプス前ドパミン利用可能性の増加の証拠、および視床におけるグルタミン酸レベルの変化が含まれます。
デフォルトモード結合性の変化 (Altered Default Mode Connectivity)
多くの研究者が、統合失調症患者がDMNの結合性に変化を持っている可能性があるという考えに興味を持っています。上記で述べたように、DMNに帰属される主要な機能の1つは、内部感情状態への注意です。多くの研究者が、個人が特定のタスクに従事したり外部世界に注意を向けたりする必要がある場合、内部状態へのこの注意が抑制されなければならず、これが、個人が集中した認知タスクに従事するときに通常見られるDMNの活動の低下につながると仮説を立てています。したがって、統合失調症の観点からのDMNへの関心は、統合失調症患者が持つ可能性のある知覚体験の変化、あるいは妄想的な内容の一部が、DMNの活動だけでなく結合性の変化によって、内部の思考、感情、および経験への注意の増強を反映している可能性があるという考えです。さらに、認知タスクの実行中にDMN活動を抑制できないことが、統合失調症における認知機能障害につながるメカニズムの1つであると仮説が立てられています。
このような仮説は興味深いものですが、広範な障害としての統合失調症におけるデフォルトモード結合性の異常に関する実証文献は、実際には非常に混在しており、明確なデフォルトモード結合性の障害に関する一貫した証拠は比較的少ないです。しかし、この文献における課題の1つは、多くの研究が、症状プロファイルに関係なく、統合失調症患者と健常対照者を比較することに焦点を当ててきただけであることかもしれません。同じ診断を持つ個人間、および同じ個人内で時間とともに症状プロファイルと重症度が大きく異なる可能性があるという事実を考えると、特定の症状の重症度の変動に関連してデフォルトモード結合性を調べることも重要です。例えば、聴覚的幻覚と、DMNの一部である側頭頭頂接合部および皮質正中線構造(前部および後部帯状回、楔前部)の結合性の変化、ならびにデフォルトモードと言語ネットワーク領域間の結合性の障害を結びつける研究が増えています。関係の方向性は、聴覚性言語幻覚の存在が、これらのデフォルトモード構造間の結合性の低下と関連する傾向があるということです。これは、聴覚幻覚がデフォルトモード領域の過結合性または過活動性による内部状態への異常な注意を反映しているという仮説の下で予測されていた方向ではないかもしれませんが、内部状態の監視に重要であると考えられているネットワークの障害が幻覚を持つ個人に存在するという考えと一致しています。
統合失調症における機能的脳結合性のグラフ理論分析
本章の序盤で、脳組織の主要な特性を要約する手段として、機能的脳結合性を理解するためのグラフ理論アプローチの応用について説明しました。このようなアプローチは、統合失調症における機能的(および構造的)脳結合性の研究にますます適用されています。知見はやや混在し複雑ですが、特に特定のネットワークを調査する際に、統合失調症患者がグローバル効率とローカル効率の両方で低下を示すという傾向が明らかになっています。グローバル効率および/またはローカル効率の低下と最も頻繁に関連するネットワークは、前帯状回および/または島(すなわち、帯状蓋ネットワーク)または前頭領域(すなわち、前頭頭頂ネットワーク)を含む傾向があります。このような知見は、ROIベースまたはICAベースの方法を用いた他の多くの研究と一致しており、統合失調症における前頭頭頂ネットワークおよび帯状蓋ネットワーク内またはネットワーク間の機能的結合性の低下も特定されています。興味深いことに、前頭頭頂ネットワークおよび帯状蓋ネットワークのローカル結合性とグローバル結合性の両方の低下は、統合失調症における認知機能障害とも関連しており、これらのネットワークの変化の機能的関連性を示唆しています。
また、統合失調症患者が脳の「ハブ」領域、特に高度に相互接続されたハブの「リッチクラブ」の一部であると考えられている領域の機能と結合性が損なわれているという興味深い証拠もあります。より具体的には、統合失調症患者は、島、上側頭前頭皮質、楔前部、上頭頂小葉を含む、ハブとして特徴付けられる領域間の接続において有意な減少を示します。しかし、これらのハブ脳領域と非ハブ領域間の接続(「フィーダー」接続と呼ばれる)や、非ハブ領域間の接続には減少を示しません。重要なことに、前部島などの一部の領域では、参加係数によって指標化されるハブ性の程度の個人差も、統合失調症における認知機能障害のレベルを予測します。収束する証拠の一部として、多くの研究は、前部島を含むこれらのハブ領域の少なくともいくつかにおいて、灰白質容量の減少も示しています。
統合失調症の機能的結合性文献のまとめ
前述のように、統合失調症における機能的結合性に関する非常に大規模な文献があり、デフォルトモード結合性に関する結果を含め、一部の知見は混在しています。しかし、いくつかの知見は一貫して再現可能であることが際立っています。圧倒的に最も堅牢な一連の知見は、前頭、帯状、線条体、および小脳領域との視床結合性の低下であり、これは統合失調症が「切断症候群」であるという長年の理論と強く一致する結果です。さらに、統合失調症においてグローバル効率とローカル効率の低下を示唆する文献が増えており、特に前頭頭頂ネットワークと帯状蓋ネットワークにおいて顕著であり、この疾患におけるハブ領域の結合性の低下の証拠もあります。これら両方の知見は認知機能を予測します。これらの2つの知見のセットは関連している可能性が非常に高く、統合失調症で見られる大規模な視床の切断は、ネットワークが情報を処理および伝達する効率の低下に寄与し、またハブ領域がネットワーク間で情報を伝達する能力を妨げる可能性があります。多くの場合、これらの障害は疾患の発症時または発症前にも存在し、より顕著です。
統合失調症における機能的結合性に関する文献の要約
前述のとおり、統合失調症における機能的結合性に関する研究は非常に多岐にわたり、デフォルトモード結合性に関する結果を含め、一部の知見は混在しています。しかし、一貫して再現性のあるいくつかの知見が際立っています。
最も堅牢な一連の知見は、視床と前頭葉、帯状回、線条体、小脳領域との結合性低下であり、これは統合失調症を「切断症候群」とみなす長年の理論と強く一致しています。さらに、統合失調症におけるグローバル効率とローカル効率の低下を示唆する文献が増加しており、特に前頭頭頂ネットワークと帯状蓋ネットワークにおいて顕著です。また、この疾患におけるハブ領域の結合性低下の証拠もあり、これらはいずれも認知機能を予測します。
統合失調症で見られる大規模な視床の切断が、ネットワークが情報を処理および転送する効率の低下に寄与し、またハブ領域がネットワーク間で情報を通信する能力を妨げる可能性があるため、これら2つの知見のセットは関連している可能性が非常に高いです。多くの場合、これらの機能障害は疾患の発症時、あるいはそれ以前から存在し、本格的な精神病を発症する個人でより顕著です。したがって、これらの知見は、機能的結合性の異常が精神病の病態生理の重要な構成要素であり、将来のバイオマーカーおよび介入研究にとって有望な道筋であることを示唆しています。
大うつ病における機能的結合性の異常
統合失調症に関する文献と同様に、大うつ病における機能的結合性の様々な側面を検討する実証研究が急速に増加しており、多様な分析アプローチが用いられています。統合失調症の文献と同様に、知見には多様性が見られますが、これは方法論、疾患の病期、投薬因子に関連している可能性があります。しかし、機能的脳結合性の変化が気分障害の発症および/または維持にどのように寄与しているかについて重要な手がかりを提供する、いくつかの新たな傾向が見られます。
扁桃体結合性と感情調節
扁桃体は、恐怖条件付け、脅威感受性、および一般的に環境中の顕著な出来事への応答における役割のために、多大な研究の注目を集めてきた皮質下構造です。さらに、多くの研究が、貧困、ストレス、トラウマが扁桃体の発達、機能、構造に影響を与えることを示しています。さらに、多くの研究者が、扁桃体構造/機能におけるそのような変化が、初期の逆境を後の精神的健康の負の結果と結びつける媒介メカニズムであると示唆しています。重要なことに、ヒトにおけるこれらの知見は、ストレスや母性ケアが扁桃体の機能と構造、および行動に有害な影響を与えることを示す多数の動物研究とも一致しています。
扁桃体は、脳の他の領域と正と負の両方の結合性を示す標準的なパターンを示します。図1.28-4は、学齢期の児童と青年を対象としたサンプルを用いて、子供と成人の両方で見られる典型的な扁桃体結合性パターンを示しています。ご覧のとおり、扁桃体は、海馬を含む内側および前側頭葉の他の領域、ならびに腹内側前頭前野(この領域では10歳以上で)と正の安静時機能的結合性を示します。扁桃体と正の結合性を示すこれらの領域は、感情およびストレス応答性にとっても重要です。
いくつかの研究では、扁桃体は、前帯状回(図1.28-4のように腹内側前頭前野)の腹側領域、島皮質の腹側領域、および前側頭極を含む、サリエンスネットワークの他の領域と結合することがわかっています。対照的に、図1.28-4に示すように、扁桃体は、両側の背側前頭葉および頭頂葉領域、ならびに背側前帯状回と負の機能的結合性(逆相関)も示します。海馬および扁桃体と負の相関を示すこれらの領域は、感情調節の研究で活性化されるのと同じ領域であり、前頭頭頂ネットワークおよび帯状蓋ネットワークの一部である領域と大部分が重複しています。重要なことに、これらの「逆相関」は、扁桃体領域で起こる感情およびストレス応答のトップダウン調節を示すと考えられており、気分および効果的な機能における扁桃体と前頭頭頂ネットワークおよび帯状蓋ネットワーク間のこれらの接続の完全性の重要な役割を示唆しています。

図1.28-4. 扁桃体結合性のパターン
未発表データ。扁桃体と正の機能的結合性を示す領域(明るい灰色)と、扁桃体と負の機能的結合性または「逆相関」を示す領域(濃い灰色)が示されています。扁桃体と正の機能的結合性を示す領域には、海馬、腹内側前頭前野、前側頭極が含まれます。扁桃体と負の機能的結合性を示す領域には、背外側前頭前野が含まれます。
重要なことに、いくつかの研究は、幼少期のストレスや母性剥奪が、扁桃体と腹内側前頭前野、前帯状回前部、およびその他の領域との間の結合性の変化と関連していることを示しています。これらの結合性は、効果的な感情調節に不可欠であると考えられています。さらに、大うつ病患者では、扁桃体とサリエンスネットワーク、前頭頭頂ネットワーク、および帯状蓋ネットワークの両方の構成要素との間の結合性の変化パターンも発見されています。
研究によって結果には多少のばらつきがありますが、より一貫した知見の1つは、扁桃体と前頭前野の様々な領域(背外側前頭前野、吻側前頭前野、腹側前頭前野など)、および前帯状回の一部との結合性の低下です。前述のとおり、一つの仮説は、このような扁桃体から前頭前野/帯状回への結合性低下が、感情処理のトップダウン調節の障害を反映しており、このような結合性低下がこの精神病理における感情調節の困難に寄与しているというものです。興味深いことに、これらの結合性の変化パターンは、子供、青年、若年・中年成人、高齢者を含むうつ病患者の間で、生涯を通じて見られることが判明しており、扁桃体から前頭前野/帯状回への結合性変化がうつ病性障害の生涯にわたる特徴である可能性を示唆しています。
さらに、このような結合性の変化は、うつ病の母親を持つことでうつ病発症のリスクがある人々にさえ存在しており、このような結合性障害が明らかな疾患の発症に先行し、病態生理学的経路の一部である可能性さえあることを示唆しています。例えば、うつ病の初期兆候や症状を持つ未就学児では、扁桃体と内側前頭前野の結合性低下が、感情調節の障害と負の感情の増加と関連しています(図1.28-5参照)。
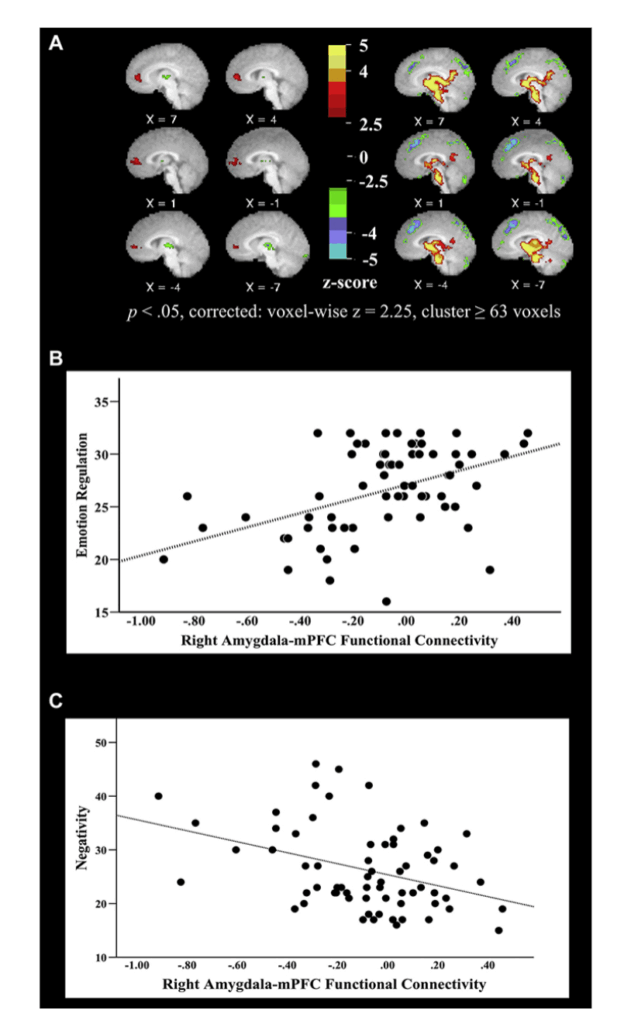
図1.28-5. 扁桃体から内側前頭前野への結合性と子どもの感情調節の関係
図のパネルAは、うつ病の初期兆候を示す未就学児の全脳解析から得られた画像で、扁桃体との結合性の変化が感情調節の障害と関連している領域を示しています。パネルBとCは、扁桃体-内側前頭前野結合性の低下と、親が報告した感情調節の障害および負の感情の増加との関係を示しています。(カラー画像については電子書籍を参照。)(Gaffrey MS, Barch DM, Luby JL, Petersen SE. Amygdala-functional connectivity is associated with emotion regulation and amygdala reactivity in 4-6-year-olds. JAACAP. 2021;60(1):176-185. ©2021 American Academy of Child and Adolescent Psychiatry. doi: 10.1016/j.jaac.2020.01.024より転載。)
デフォルトモードネットワーク結合性と反芻
大うつ病に関連する結合性変化の一貫した知見の2番目の領域は、DMN(デフォルトモードネットワーク)結合性の分野です。前述のとおり、DMNに帰属される主要な機能の1つは、内部感情状態への注意を調節し、自己言及的処理を司ることです。現象学的に、うつ病の主要な特徴の1つは、負の気分、負の自己評価、そして頻繁に負の出来事や思考に対する反芻的な対処スタイルへの没頭です。
したがって、研究が大うつ病におけるデフォルトモード領域間の結合性の増加、特に背内側前頭前野を含むDMNのより前部を形成する領域における増加を示唆しているのは驚くべきことではありません。興味深いことに、うつ病患者では、背内側前頭前野と、サリエンスネットワークの一部である吻側前島皮質との間の結合性の増加も示されています。DMNの一部とサリエンスネットワーク間のこの結合性の増加は、顕著な出来事への注意に関連するDMN活動の過剰な統合または優位性を潜在的に反映していると解釈されており、これは周囲の環境における顕著な情報に対する負の評価バイアスに寄与している可能性があります。
多くの研究はまた、膝下前頭前野とDMNの結合性増加の証拠を発見しています。膝下前頭前野は、気分障害に関する文献で一貫して注目されてきた脳領域であり、MDD患者では、生涯にわたってグリア細胞の減少、代謝の変化、容積の減少および/または薄化を示すことが示されています。膝下前頭前野は、行動的引きこもりと効果的な負の評価のメカニズムにとって重要であると仮説が立てられており、この領域の活性化は、資源の節約(すなわち、引きこもり)、安全志向の行動の促進、および罪悪感の経験と関連しています。
重要なことに、研究は、膝下前頭前野とDMN間の過結合の程度が、うつ病患者の反芻レベルを予測することを示しています。この研究は、この過剰に強力な統合が、DMNによってサポートされる自己言及的処理と、膝下前頭前野によってサポートされる負の情動と引きこもりを結びつけ、MDD患者における反芻と感情調節の障害のサイクルにつながるという仮説をもたらしました。扁桃体結合性の障害と同様に、このようなデフォルトモードの変化は、うつ病の家族歴を持つことによってうつ病発症のリスクがある人々にさえ存在しており、このような結合性障害が明らかな疾患の発症に先行し、病態生理学的経路の一部である可能性さえあることを再び示唆しています。
線条体結合性と快感消失
近年、うつ病内および診断横断的に快感消失の機能的結合性相関関係を検討する研究が増加しています。快感消失は、快感を経験する能力の低下、または快感を予測する能力の低下として定義されます。この研究の多くは、側坐核や尾状核などの線条体領域から、全ネットワーク(例:デフォルトモード、帯状蓋、サリエンスなど)またはこれらのネットワーク内の特定の領域(例:DMNの膝下帯状回、前頭頭頂ネットワークのDLPFC)への結合性に焦点を当てています。この線条体結合性への焦点は、線条体の機能的活動と結合性が報酬応答性における個人差を理解するために重要であるという証拠に後押しされています。報酬応答性は快感消失を考える上で中心的な概念です。大うつ病患者における知見はやや混在していますが、いくつかの研究では、線条体と他のネットワーク(デフォルトモード、前頭頭頂、帯状蓋ネットワークなど)との結合性低下が、より大きな快感消失と関連していることが発見されています。
大うつ病における機能的結合性文献の要約
統合失調症における機能的結合性に関する文献と同様に、うつ病における結合性に関する文献も多岐にわたり、やや混在しています。しかし、ここでもいくつかの知見が、ある程度再現可能であると際立っています。
- 第一に、扁桃体と、前頭頭頂ネットワークおよび帯状蓋ネットワークの一部である前頭前野/帯状回領域との間の結合性の変化の証拠です。これらの結合は、感情のトップダウン調節と、ストレスや顕著な環境イベントへの応答に重要な扁桃体などの領域に対する制御にとって極めて重要であると考えられています。これらの結合性の変化は、様々な形態の早期ストレスやトラウマを経験している個人に存在し、大うつ病の個人では生涯にわたって存在し、うつ病のリスクがある人々にさえ存在します。したがって、これは、ストレスとトラウマがその後の感情調節不全と気分病理の発症に結びつく病因経路に関する重要な手がかりとなる可能性があります。
- 第二に、比較的矛盾のない結果の領域は、膝下前頭前野とこのネットワークの他の領域間の関連を含む、DMNの結合性増加の証拠です。この結合性増加は、大うつ病患者における反芻と特異的に関連しており、自己言及的処理をサポートする脳領域と、負の情動処理と引きこもりの影響を受ける脳領域との間の過剰に強固な統合を反映している可能性があります。
- 第三に、線条体とデフォルトモード、前頭頭頂、帯状蓋ネットワークとの間の結合性障害とうつ病および診断横断的な快感消失との関連の証拠ですが、そのような知見の堅牢性と再現性を確立するためには、さらなる研究が必要です。
結論
本章の目的は、ヒトの機能的コネクトームを評価する方法、ヒトの脳に存在する主要な内在性機能的結合性ネットワークに関する既存の知識、およびこれらのネットワークにおける結合性異常が主要な精神疾患の病態生理を理解する上でどのように関連しているかについて、現在の分野の簡単な概要を提供することでした。多くの点でこの分野はまだ初期段階にありますが、画像モダリティ、機能的結合性方法、および集団を越えて特定された機能的脳ネットワークの注目すべき一貫性と堅牢性は、ヒトの行動への影響を理解することの重要性と関連性を証明しています。
ライフスパン全体にわたるヒトコネクトームプロジェクトからの研究は、内在性ネットワーク結合性の個人差が、ヒトにおける正負両方の幅広い特性および特徴と関連していることを確認しました。さらに、この研究は、脳の機能的組織の核心的な側面が、生涯にわたる発達的進化を伴うものの、人生の早期に生じることを示しています。さらに、本章で選択的にレビューされた研究は、異なる精神病理の形態と関連する堅牢で再現可能かつ区別可能な機能的結合性の変化が存在することを示しています。これには、精神病と関連する視床結合性の変化、精神病における認知機能に関連する前頭頭頂ネットワークおよび帯状蓋ネットワークのグローバル効率とローカル効率の障害、統合失調症における前部島などの領域のハブ構造の変化に関する確固たる証拠が含まれます。大うつ病においては、感情調節の理解に関連する扁桃体から前頭前野-帯状回への結合性の変化、および反芻に関連するデフォルトモード結合性の増加に関する一貫した証拠があります。
次の重要なステップ(いくつか進行中)は、これらの結合性変化が精神病とうつ病、または特定の症状次元の発症において果たす役割、ならびに結合性が治療によってどの程度修正され得るか、そしてそれが症状改善の媒介として機能するかどうかを検討することです。さらに、この神経回路分析レベルの変化を、遺伝子、遺伝子発現、神経伝達物質シグナル伝達、および/または細胞構造などの他の分析レベルの変化と結びつけることが不可欠となるでしょう。
参考文献
- Andreasen NC, Paradiso S, O’Leary DS. “Cognitive dysmetria” as an integrative theory of schizophrenia: a dysfunction in cortical-subcortical-cerebellar circuitry? Schizophr Bull. 1998;24(2):203-218.
- Anticevic A, Haut K, Murray JD, et al. Association of thalamic dysconnectivity and conversion to psychosis in youth and young adults at elevated clinical risk. JAMA Psychiatry. 2015;72(9):882-891.
- Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med. 1995;34(4):537-541.
- Chen JE, Glover GH, Greicius MD, Chang C. Dissociated patterns of anticorrelations with dorsal and ventral default-mode networks at rest. Hum Brain Mapp. 2017;38(5):2454-2465.
- Cheng W, Rolls ET, Qiu J, et al. Functional connectivity of the human amygdala in health and in depression. Soc Cogn Affect Neurosci. 2018;13(6):557-568.
- Cole MW, Bassett DS, Power JD, Braver TS, Petersen SE. Intrinsic and task-evoked network architectures of the human brain. Neuron. 2014;83(1):238-251.
- Dauvermann MR, Whalley HC, Romaniuk L, et al. The application of nonlinear dynamic causal modelling for fMRI in subjects at high genetic risk of schizophrenia. Neuroimage. 2013;73:16-29.
- Finn ES, Bandettini PA. Movie-watching outperforms rest for functional connectivity-based prediction of behavior. Neuroimage. 2021;235:117963.
- Fox MD, Raichle ME. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci. 2007;8(9):700-711.
- Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ. Psychophysiological and modulatory interactions in neuroimaging. Neuroimage. 1997;6(3):218-229.
- Gabbay V, Ely BA, Li Q, et al. Striatum-based circuitry of adolescent depression and anhedonia. J Am Acad Child Adolesc Psychiatry. 2013;52(6):628-641.e13.
- Geller WN, Liu K, Warren SL, et al. Specificity of anhedonic alterations in resting-state network connectivity and structure: a transdiagnostic approach. Psychiatry Res Neuroimaging. 2021;317:111349.
- Giraldo-Chica M, Woodward ND. Review of thalamocortical resting-state fMRI studies in schizophrenia. Schizophr Res. 2017;180:58-63.
- Gong L, He C, Zhang H, Zhang H, Zhang Z, Xie C. Disrupted reward and cognitive control networks contribute to anhedonia in depression. J Psychiatr Res. 2018;103:61-68.
- Gordon EM, Laumann TO, Adeyemo B, Huckins JF, Kelley WM, Petersen SE. Generation and evaluation of a cortical area parcellation from resting-state correlations. Cereb Cortex. 2016;26(1):288-303.
- Gordon EM, Laumann TO, Gilmore AW, et al. Precision functional mapping of individual human brains. Neuron. 2017;95(4):791-807. e7.
- Hamilton JP, Farmer M, Fogelman P, Gotlib IH. Depressive rumination, the default-mode network, and the dark matter of clinical neuroscience. Biol Psychiatry. 2015;78(4):224-230.
- He Z, Lu F, Sheng W, et al. Abnormal functional connectivity as neural biological substrate of trait and state characteristics in major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2020;102:109949.
- Hyatt CJ, Calhoun VD, Pearlson GD, Assaf M. Specific default mode subnetworks support mentalizing as revealed through opposing network recruitment by social and semantic FMRI tasks. Hum Brain Mapp. 2015;36(8):3047-3063.
- Li BJ, Friston K, Mody M, Wang HN, Lu HB, Hu DW. A brain network model for depression: from symptom understanding to disease intervention. CNS Neurosci Ther. 2018;24(11):1004-1019.
- Liu R, Wang Y, Chen X, Zhang Z, Xiao L, Zhou Y. Anhedonia correlates with functional connectivity of the nucleus accumbens subregions in patients with major depressive disorder. Neuroimage Clin. 2021;30:102599.
- Ma S, Calhoun VD, Phlypo R, Adali T. Dynamic changes of spatial functional network connectivity in healthy individuals and schizophrenia patients using independent vector analysis. Neuroimage. 2014;90:196-206.
- Marek S, Tervo-Clemmens B, Nielsen AN, et al. “Identifying reproducible individual differences in childhood functional brain networks: an ABCD study.” Dev Cogn Neurosci. 2019;40:100706.
- Mulders PC, van Eijndhoven PF, Schene AH, Beckmann CF, Tendolkar I. Resting-state functional connectivity in major depressive disorder: a review. Neurosci Biobehav Rev. 2015;56:330-344.
- Noble S, Scheinost D, Constable RT. A decade of test-retest reliability of functional connectivity: a systematic review and meta-analysis. Neuroimage. 2019;203:116157.
- Noble S, Scheinost D, Constable RT. A guide to the measurement and interpretation of fMRI test-retest reliability. Curr Opin Behav Sci. 2021;40:27-32.
- Palaniyappan L, Deshpande G, Lanka P, et al. Effective connectivity within a triple network brain system discriminates schizophrenia spectrum disorders from psychotic bipolar disorder at the single-subject level. Schizophr Res. 2019;214:24-33.
- Perino MT, Myers MJ, Wheelock MD, et al. Altered whole-brain resting-state functional connectivity patterns associated with pediatric anxiety and involuntary attention capture. Biol Psychiatry Glob Open Sci. 2021;1(3):229-238.
- Power JD, Mitra A, Laumann TO, Snyder AZ, Schlaggar BL, Petersen SE. Methods to detect, characterize, and remove motion artifact in resting state fMRI. Neuroimage. 2014;84:320-341.
- Rzepa E, McCabe C. Decreased anticipated pleasure correlates with increased salience network resting state functional connectivity in adolescents with depressive symptomatology. J Psychiatr Res. 2016;82:40-47.
- Rzepa E, McCabe C. Anhedonia and depression severity dissociated by dmPFC resting-state functional connectivity in adolescents. J Psychopharmacol. 2018;32(10):1067-1074.
- Scheinost D, Tokoglu F, Hampson M, Hoffman R, Constable RT. Data-driven analysis of functional connectivity reveals a potential auditory verbal hallucination network. Schizophr Bull. 2019;45(2):415-424.
- Sendi MSE, Zendehrouh E, Ellis CA, et al. Aberrant dynamic functional connectivity of default mode network in schizophrenia and links to symptom severity. Front Neural Circuits, 2021;15:649417.
- Sharma A, Wolf DH, Ciric R, et al. Common dimensional reward deficits across mood and psychotic disorders: a connectome-wide association study. Am J Psychiatry. 2017;174(7):657-666.
- Sheffield JM, Kandala S, Tamminga CA, et al. Transdiagnostic associations between functional brain network integrity and cognition. JAMA Psychiatry. 2017;74(6):605-613.
- Smith SM, Miller KL, Salimi-Khorshidi G, et al. Network modelling methods for fMRI. Neuroimage. 2011;54(2):875-891.
- Smith SM, Nichols TE, Vidaurre D, et al. A positive-negative mode of population covariation links brain connectivity, demographics and behavior. Nat Neurosci. 2015;18(11):1565-1567.
- Sylvester CM, Yu Q, Srivastava AB, et al. Individual-specific functional connectivity of the amygdala: a substrate for precision psychiatry. Proc Natl Acad Sci U S A. 2020;117(7):3808-3818.
- Tang Y, Kong L, Wu F, et al. Decreased functional connectivity between the amygdala and the left ventral prefrontal cortex in treatment-naive patients with major depressive disorder: a resting-state functional magnetic resonance imaging study. Psychol Med. 2013;43(9):1921-1927.
- van den Heuvel MP, Sporns P, Collin G, et al. Abnormal rich club organization and functional brain dynamics in schizophrenia. JAMA Psychiatry. 2013;70(8):783-792.
- VanTieghem MR, Tottenham N. Neurobiological programming of early life stress: functional development of amygdala-prefrontal circuitry and vulnerability for stress-related psychopathology. Curr Top Behav Neurosci. 2018;38:117-136.
- Yang XH, Tian K, Wang DF, et al. “Anhedonia correlates with abnormal functional connectivity of the superior temporal gyrus and the caudate nucleus in patients with first-episode drug-naive major depressive disorder.” J Affect Disord. 2017;218:284-290.
- Yeo BTT, Krienen FM, Sepulcre J, et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106(3):1125-1165.
