
アディーナ・ロスキーズによる論文「How Does Neuroscience Affect Our Conception of Volition?」の全文を日本語に翻訳。学術的な内容を正確に伝えるため、専門用語は一般的な訳語を用い、必要に応じて補足。
神経科学は私たちの意志の概念にどう影響を与えるか?
アディーナ・L・ロスキーズ
ダートマス大学哲学部、ハノーバー、ニューハンプシャー州 03755;
email: adina.roskies@dartmouth.edu
Annu. Rev. Neurosci. 2010. 33:109–30
キーワード
自由意志、決定、意図、制御、主体性、決定論
要旨
意志(volition)や意図(the will)に関する明確な概念は存在しないものの、私たちは意志、主体性(エージェンシー)、そして自発的行動を特徴づける直感的な考えを持っている。本稿では、私たちの意志に関する直感的な観念に関わる、数々の神経科学的研究から得られた結果を概観する。これらの神経科学的な結果は、私たちが意志や意図に関連すると見なす行動を媒介する神経回路について、いくつかの洞察を提供する。一部の研究者は、神経科学が私たちの自由意志に関する見解を覆すだろうと主張しているが、今日に至るまで、私たちの常識的な信念を根本的に破壊することに成功した結果は一つもない。それでもなお、神経科学から浮かび上がる全体像は新たな問いを提起しており、最終的には、自由意志のために何が必要かといういくつかの直感的な観念に対して、プレッシャーをかけることになるかもしれない。
目次
- はじめに … 110
- 意志への哲学的・実験的アプローチ … 110
- 意志と自由 … 112
- 意志への神経科学的アプローチ … 113
- 開始としての意志 … 113
- 意図としての意志 … 115
- 意思決定としての意志 … 117
- 実行制御としての意志 … 121
- 感覚としての意志 … 122
- 結論 … 123
はじめに
神学者や哲学者の間で長く議論の的であった「意志(the will)」、すなわち意志の能力(faculty of volition)は、19世紀の心理学者にとって科学的研究の対象であった。しかし、哲学と心理学における行動主義革命とともに、意志は科学のレーダーから消え去った。意志は、経験的アプローチにはなじまない、あまりにも徹底した心的概念であると見なされたからである。しかし、より最近の二元論的視点の放棄と、脳を研究するための新しい技術の発展に伴い、意志というテーマは一種のルネサンスを迎えている。哲学における自由意志の議論もまた花開いており、意志の復活の推進力の多くは、認知科学および神経科学によって提起された新たな挑戦と機会にある(サイドバー「自由意志の哲学」参照)。
意志への哲学的・実験的アプローチ
哲学においても科学においても、論争の余地のない、単一の意志の概念というものは見当たらない(Audi 1993; Brass & Haggard 2008; Zhu 2004a,b)。一般的に言えば、意志とは、内生的な行動、自律性、または選択の根拠を指すために用いられる構成概念である。その詳細については直感は様々である。ある者は、自発的行動を反射的または環境によって特定される行動と対比させる。またある者は、意志は主に行動を決定する決定を下すことに関与すると主張する。さらに別の者は、これらの見解を組み合わせて、選択は心的行為に他ならないと主張する。意志は、決定するという内生的な心的行為や意図の形成を指すために使われることがある(Searle 1983, Zhu 2004a)。また、決定や意図そのものを指して使われることもある(Adams & Mele 1992)。意志された行為そのものの成功裏の実行とは独立しており、意志の中心的な側面は試みることにあると示唆する者もいる(Adams & Mele 1992)。
上記リストの異質性は、意志の概念に対する神経科学の影響をレビューする上での困難さの一つの手がかりとなる。というのも、意志の明確な概念がないことは、それを実験的に研究する課題を複雑にするからである。加えて、神経科学的研究が意志の概念に関わるためには、意志は何らかの方法で操作可能にされなければならない。これらの考察を踏まえ、私は議論を5つの異なる実験的テーマに沿って構成する。これらは、個別にあるいは組み合わさって、意志という直感的ではあるが明確ではない概念によって示されるものの多くを捉えているように思われる。これらのテーマとは、(a) 行動の開始、(b) 意図、(c) 決定、(d) 抑制と制御、そして (e) 主体性の現象学である。これら5つのテーマは、Haggard & Brass(2008)による意図的行動の「何を、いつ、すべきかどうか(What, when, whether)」モデルに緩やかに対応している。
上記の各テーマに関連する、比較的特定可能な研究群が存在するが、それらの多くは互いに混じり合っている。さらに、これらの各々は、意志に関する常識的な概念のいくつかの要素に対応している。例えば、もし自発的行動を刺激によって生成された行動と対比させるならば、自己開始された運動と、外部刺激への応答である類似の運動とを区別する神経イベントを調べることは、内生的に生成された行動の根底にある近位のメカニズムへの洞察を提供するはずである。
しかし、もし意志が本質的に運動と結びついているのではなく、むしろ将来の行動のための抽象的な計画と結びついていると考えるならば、単純な運動につながる近位のメカニズムは、人が持つ、あるいは形成する長期的な計画や意図よりも関心が低いかもしれない。意図に関するいくつかの研究は、運動計画のこれらの高レベルの側面に取り組もうとしている。意志に関する哲学的な議論は、しばしば、ある方法または別の方法で行動することを選択する能力に焦点を合わせる。歴史的にこの選択への重点は、選択の心性論と行動の物理主義との間の暗黙の二元論の名残かもしれないが、決定を形成する手段としての決定の根底にあるプロセスは、たとえ二元論を否定したとしても、意志の中心的な側面であるように思われる(現代のほとんどの哲学者や科学者がそうであるように)。
意志への異なるアプローチは、意志が将来の行動に与える予期的影響よりも、被験者が行動を抑制または制御する現行の能力に焦点を合わせる。制御が意志の中心であると直感的には考えないかもしれないが、道徳や法律における責任の帰属にとっての制御の重要性を認識することは、意志に対する制御の関連性を明確にするのに役立つかもしれない。さらに、心理学と神経科学からのいくつかの証拠は、行動はしばしば無意識的に開始されることを示唆しており(例:Libet 1985)、したがって、もし自由意志が存在するとすれば、それは無意識的に開始された行動に対する制御または拒否権の形をとることになるだろう。最後に、私たちが自由に行動したり選択したりできると信じているかどうかにかかわらず、私たちは通常、特定の行動を自己に起因するものとして、他の行動をそうでないものとして知覚する。ほとんどの人は、自発的行動には現象学(phenomenology)が伴うことに同意しており、神経科学は主体性の感覚の生理学的基盤を解明し始めている。
したがって、これら5つのテーマに関する研究は、私たちの意志の概念に影響を与えたり、解明したりするかもしれない。始める前にいくつかの注意点がある。第一に、意志の概念自体に関する合意の欠如と、意志および主体性の議論との密接な関係に鑑み、私は、願わくはさらに話をややこしくすることなく、意志(volition)、意図(will)、主体性(agency)という用語を互換的に使用することにしたい。これは、それらの間に実質的な区別がなされないということではない。第二に、議論される各分野の広範な文献のため、私はこれらの各研究分野の網羅的なレビューではなく、神経科学がどのように意志の概念に影響を与えたか、与えることができるか、あるいは与えるかもしれないかについての議論に焦点を当てる。最後に、他者の主体性を知覚し、帰属させることの神経基盤に関するかなりの文献があることを認識している(Ciaramidaro et al. 2007, Cunnington et al. 2006, de Lange et al. 2008, Fogassi et al. 2005, Hamilton & Grafton 2006, Ramnani & Miall 2004, Rizzolatti & Sinigaglia 2007)。いくつかの仮説では、他者の主体性の知覚を支える神経システムは、自己の主体性の知覚において作動するものと同じである(Ciaramidaro et al. 2007, Cunnington et al. 2006, Fogassi et al. 2005, Iacoboni et al. 2005, Lamm et al. 2007, Rizzolatti & Sinigaglia 2007)。この文献は間接的に私たちの意志の理解に関わるかもしれないが、ここでは議論しない。最後に、この論文は意志に関する哲学的研究をレビューすることを試みるものではない。しかし、私は意志と自由の関係についての短い余談から始める。
用語解説
- 意志(Volition): 自発的な行動や選択を可能にする能力;意図。
- 意図(Intention): 将来の行動のための計画を表す心的状態。多くの種類の意図が存在しうる。
- 現象学(Phenomenology): ある経験が、その主体の観点からどのようなものであるかということ。
- 二元論(Dualism): 心的(mental)なものと物理的(physical)なものは、根本的に異なる2つの存在論的カテゴリーに属し、互いに還元することはできない。
サイドバー:自由意志の哲学
哲学者は伝統的に、自由意志の問題を決定論と非決定論の観点から枠組み作ってきた。両立論者(Compatibilists)は決定論と両立する自由の説明を提供しようと試みる。非両立論者(incompatibilists)は、もし決定論が真実なら私たちは自由ではありえないと否定する。非両立論者には2つの種類がある:固い決定論者(Hard determinists)は決定論の真実の側に賭け、私たちが自由であることを否定する。リバタリアン(Libertarians)は、私たちは非決定論的な出来事のおかげで自由であると主張する。両立論者の課題は、もし私たちがそうする以外に行動できなかったとしても、いかにして私たちが自由であり、道徳的に責任を負うことができるかを示すことである。リバタリアンの課題は、自らの描像を科学的に妥当なものにし、同時に、非決定論的な出来事が、主体性と責任を与えるために、いかにして選択や行動と適切なつながりを持つことができるかを示すことである。
伝統的に、自由は道徳的責任と密接に結びついていると考えられてきた。しかし現在では、一部の哲学者は道徳的責任を自由から切り離そうと試みている。また他の者は、決定論ではなく、行動の原因のメカニズム的説明こそが、自由と責任への最大の脅威となると示唆している。古い哲学的問いに対するこれらの新しい視点、そして行動の神経基盤を理解することへの科学的進出は、自由意志の哲学を熟考する上でエキサイティングな時代をもたらしている。
意志と自由
私たちの意志の概念を、人間の自由に関する問いから切り離すことは、不可能ではないにしても困難である。意志という構成概念は、主として自律的な主体性の可能性、性質、または感覚を説明するために存在する。意志に関連する神経科学の文献を議論する前に、自由意志の問題を裁定するにはあまり見込みのない神経科学的アプローチについていくつか述べたい。
哲学において、自由に関する議論は伝統的に、自由が決定論とどう関係するか、そして決定論が真実であるか否かという問いに部分的にかかってきた。非両立論者は決定論が自由を排除すると信じているのに対し、両立論者は決定論が自由と両立するか、あるいは自由にとって必要でさえあると信じている。宇宙(あるいは脳)が決定論的であるか否かという問いは経験的な事実の問題であるが、それは神経科学によって確立できる事実ではない。一部の神経科学者は、神経科学的研究が、神経系のランダム性、ノイズ、または確率的行動における非決定論の神経的現れを特定することによって、決定論の真偽を解明できると考えているようである。これは、少なくとも2つの理由から、誤りであると私は信じる。神経科学は神経系におけるランダム性や確率的行動を反映しているように見えるデータを提供するかもしれないが、神経科学的研究の認識論的限界は、私たちが神経科学的技術から集める証拠が、その決定を下すには不十分な基盤であるというものである(Roskies 2006)。さらに、単に神経系におけるランダム性を確立するだけでは、人間の自由を説明するには不十分であろう。ランダム性は、それが自由の根拠となるためには、意志のプロセスにおいて適切な役割を果たしていることが示されなければならない。
第一の点に関して、神経科学が決定論の真偽に関わるためには、何か測定するものが必要であり、それは決定論を予測可能性として操作可能にすることによって行われる。しかし、予測可能性はせいぜい決定論の貧弱ないとこであり、その家系のルーツを裏切ることもある。決定論的なシステムは原理的には予測可能であるが、実際には予測可能性は決定論への導きにはならない。あるレベルで確率的に見える振る舞いは、より低いレベルでの決定論的プロセスの結果である可能性がある。例えば、Mainen & Sejnowski (1995)は、直流電流の注入では確率的に見えたスパイクのタイミングが、実際には、ニューロンがその通常の入力を特徴づける同じ可変電圧パターンで注入された場合には非常に信頼性が高いことを見出した。この発見は、ニューロンの発火が決定論的に作動することを示唆するかもしれない(ただしDorval 2006参照)。しかし、Mainen & Sejnowskiのデータは、スパイクのタイミングが著しく信頼性が高いものの、完全ではなかったことを示していた。これらの不一致は、確率的行動に起因する可能性もあれば、監視されていない入力や、試行間で変化したニューロンの他の特徴といった、完全に決定論的な要因に起因する可能性もある。同様の曖昧さは、より細かい粒度で見ても生じる。例えば、神経伝達物質放出の確率的特性は、根本的に確率的なプロセスによるものかもしれないし、あるいは、シナプス前終末の小胞の空間分布のような、細胞以下のレベルで時間とともに構造的に変化するシステムの純粋に決定論的な操作によるものかもしれない(Franks et al. 2003)。したがって、神経科学的データから決定論について判断するためには、私たちの神経生理学的技術が教えてくれるよりもはるかに多くのニューロンの微物理的な構造について知る必要があり、また、私たちが記録しているニューロンに影響を与えるシステムの全体的な状態に関する完全な情報を持つ必要があるだろう。
神経イベントの特定の性質がランダムな行動の兆候(例:ポアソン分布)であるかもしれないが、そのような性質は決定論的なメカニズムによっても生成されうる(Glimcher 2005)。例えば、コンピュータの「乱数」生成器は単なる決定論的なアルゴリズムであり、多くのカオス過程も決定論的である。一部の神経科学者はニューロンの行動が非決定論的であると確信しているようだが、結論はまだ出ていない。神経系におけるノイズへの言及は非決定論の精神を呼び起こすが、ある人にとってのノイズは別の人にとってのシグナルである。脳がどのように情報をコード化し、読み出すかについての明確で完全な理解がなければ、私たちは単に説明のつかない活動をノイズとレッテル貼りし、そこから脳のプロセスが非両立論者が必要とする意味で非決定論的であると推測することはできない。そうは言っても、神経活動における変動性の役割を、決定論の問題によらずに、決定や行動に関連するものとして見る方法はあるかもしれない。
神経科学は、行動の根底にあるメカニズムを解明することによって、自由意志に関する見解に影響を与えることができる(ここでメカニズムは決定論の問題については沈黙している)。単にメカニズムを解明するだけで、一般人の自由に関する見解に影響を与えるかもしれないが(Monterosso et al. 2005, Nahmias et al. 2007)、二元論が偽であるという仮定の上では、メカニズム的または因果的な説明だけでは、自由の問題に答えるには不十分である。真の関心は、神経科学的説明が、意志が自由意志の要件に関する私たちの直感的な観念と一致する特徴を持つか、あるいは欠いているかを示すかどうかにある。神経科学的データが私たちの自由意志への信念に持つ関連性について議論する際、私は、意志の特定の経験的特徴が自由に関する通常の観念に圧力をかけるかどうかに焦点を当てる。
用語解説
- 決定論(Determinism): すべての出来事が宇宙の状態と自然法則によって完全に特定されるという主張;真にランダムまたは確率的なプロセスの否定。
- 非両立論者(Incompatibilists): 自由は決定論と両立しないと信じる人々。
- 両立論者(Compatibilists): 自由は決定論と両立すると信じる人々。
意志への神経科学的アプローチ
開始としての意志(Volition as Initiation)
意志は、反射や単純な刺激-応答の連合のような、外的に誘発される行動とは対照的に、内生的に生成される、あるいは自己開始される行動において重要であると考えられている。この見解は、行動タイプのそのような二分法は意味をなさないことなど、多くの点で批判されるかもしれない。にもかかわらず、多くの神経科学的研究は、自己開始された行動と外的に手がかりを与えられた行動中の脳活動を比較し、外的に手がかりを与えられた行動とそうでない行動を支える機能的構造の違いを発見してきた(Haggard 2008)。
単純な運動行為の内生的生成と安静時とを比較したイメージング研究は、一貫して一次運動野、SMA(補足運動野)およびpreSMA、前部帯状回の領域、大脳基底核、およびDLPFC(背外側前頭前野)の活動を示している。手がかりを与えられた応答は、行動の感覚的誘導を媒介する頭頂および外側運動前野を含むネットワークを伴うようである。
イメージング技術が利用可能になる以前、EEG記録は頭頂部で、運動活動に1〜2秒先行するゆっくりとした負の電位を明らかにした(図1)。この「準備電位」(RP)は、当初はSMAで生じると仮定されていた(Deecke & Kornhuber 1978, Jahanshahi et al. 1995)。さらなる研究は、RPが複数の構成要素プロセスを反映していることを示唆しており(Haggard & Eimer 1999, Libet et al. 1982, Shibasaki & Hallett 2006)、RPの初期構成要素の源はpreSMAに局在化されている(Shibasaki & Hallett 2006)。RPの大きさは、手がかりを与えられた運動よりも自己ペースの運動で大きく、研究はこの電気的信号の後期およびピーク位相が自発的または自己開始された運動行為に関連していることを示している。一方、最も初期の構成要素は、準備や動機付けに関連する認知プロセスにより関与しているかもしれない(Jahanshahi et al. 1995, Libet et al. 1982, Trevena & Miller 2002)。
運動の自己開始に内側前頭前野が関与することを示す豊富な証拠にもかかわらず、自己開始された行動と手がかりを与えられた行動中の脳活動の違いの源と機能の決定は、コンセンサスが少ない問題である。
用語解説
- SMA: 補足運動野(supplementary motor area)
- DLPFC: 背外側前頭前野(dorsolateral prefrontal cortex)
- 準備電位(Readiness potential, RP)またはBereitschaftspotential: EEGを用いて正中線で記録される電気的陰性電位で、自発的運動に先行する。
図1
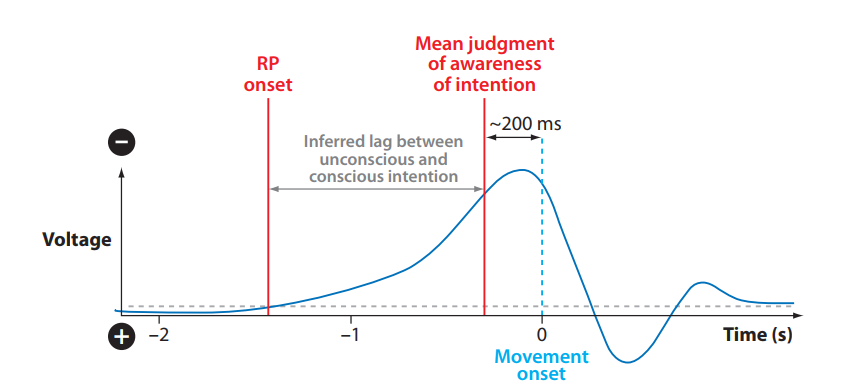
自己ペースの行動は、吻側SMA、前部帯状回、およびDLPFCで生じる(Jenkins et al. 2000)。fMRIを用いて、Deiberら(1999)は、Picard & Strick(1996)によって定義された内側前頭部の解剖学的領域に結果をマッピングし、preSMAおよびrCZ(吻側帯状回ゾーン)での活動が、外的に手がかりを与えられた運動よりも自己開始された運動で大きいことを見出した。Cunningtonら(2002)は、自己ペースの運動ではpreSMAの活動のタイミングに違いを見出したが、活動レベルには差がなく、rCZでの活動も報告している。Lauら(2004b)は、行動の選択への注意と開始とを切り離そうとし、DLPFCではなくpreSMAが開始中に優先的に活動することを見出した。しかし、この研究では、この領域でのより大きな活動は課題遂行時間と相関しており、したがって開始に対する特異性を反映していないかもしれない。Muellerら(2007)は、他の変数が制御されれば、preSMAは自己開始された運動課題と手がかりを与えられた運動課題の間で差を示さなくなり、自己開始された運動をrCZの活動と関連付け、preSMAとは関連付けない、と主張している。
サルのpreSMAの損傷は、自己開始行動を阻害するが、手がかりを与えられた行動は阻害しない(Thaler et al. 1995)。ヒトでは、SMA(preSMAを含む)領域への直接的な電気刺激は、動きたいという衝動を生じさせ、より強い刺激は行動を引き起こす(Fried et al. 1991)。さらに、preSMAへのrTMS(反復経頭蓋磁気刺激)は、課題切り替え中の手がかりのない運動系列の開始を妨害する(Kennerley et al. 2004)。これらの研究は、行動開始におけるこれらの領域の関与についてさらなる証拠を提供する。これらの領域はまた、競合する応答の自動的な抑制にも関与しているかもしれない。損傷患者を用いた無意識的プライミング研究に基づき、Sumnerら(2007)は、SMA(しかしpreSMAではない)が、代替的な自発的行動の遂行において運動計画の自動的な抑制を媒介すると報告している。これらの領域の損傷はその抑制を妨げ(Sumner et al. 2007)、不随意的な運動行動を特徴とするいくつかの症候群に現れる(Haggard 2008)。
要約すると、行動開始と最も一貫して関連付けられる領域はrCZとpreSMAであるが、それらの機能の解釈は依然として議論の余地がある。様々な要因が、多くの研究の結果を一致させることを困難にしている。いくつかのパラダイムは、被験者に指示された行動のタイミングを決定することを要求するが、他のものは行動の代替案間の選択を要求する。報告された結果は、刺激-行動連合の複雑さや葛藤によって誘発される活動といった、課題の交絡因子を反映している可能性があり、意志に特に関連する何かではないかもしれない(Nachev et al. 2008)。実際、preSMAとrCZは、応答葛藤と開始に優先的に関与する下位領域に細分化できるという証拠がいくつかある(Nachev et al. 2005, Picard & Strick 2001)。将来の研究は、自己開始された活動と外的に手がかりを与えられた活動に関与する領域、およびそのような処理を媒介する回路をよりよく解決するだろう。これらの結果は、将来の実験のためのより明確な標的を提供するかもしれない。しかし、関与する計算についてより多くが知られるまで、自己開始に関与する領域の正確な同定は、私たちの意志の概念にほとんど影響を与えない。
用語解説
- rCZ: 吻側帯状回ゾーン(rostral cingulate zone)
- rTMS: 反復経頭蓋磁気刺激(repetitive transcranial magnetic stimulation)
意図としての意志(Volition as Intention)
意図は、熟慮と行動の間のギャップを埋める表象的な状態である。おそらく、意図は意識的であることも無意識的であることもある。さらに、行動計画の異なるレベルに関与する、異なる種類の意図が存在するかもしれない(Pacherie 2006)。もし私たちが、意図がすべての自発的運動の近位の原因であると仮定するならば、行動の開始と意図の研究は同じ現象に関係しているかもしれない(私たちはこれらを近位の意図または運動意図、あるいは一部の人が呼ぶように意志(volitions)と呼ぶかもしれない)。しかし、私たちはまた、より広く、より抽象的な意味で、将来の行動のための意識的または目的を持った計画を構成する持続的な状態として、行動の実行に先立って、そして独立して存在する意図を指すことが一般的である。道徳的および法的な文脈において、私たちが人が意図的に行動したかどうかを尋ねるとき、私たちはしばしばこのより一般的な意図の概念を用いる。
一般に、意図された行動は、行動する意図を伴い、多くの人々は、自由になされた行動は私たちの意識的な意図によって引き起こされなければならないと想定している。私たちの意識的な意図の有効性は、ベンジャミン・リベットの研究によって挑戦された。彼は、動く意図の気づきの相対的なタイミングと、行動の開始を反映する神経信号とを調べた。リベットは、準備電位(RP)の開始時間は、動くという衝動や意図の気づきよりも約350ミリ秒以上前に起こると報告した(Libet 1985; Libet et al. 1982, 1983b,c)。リベットらは、この食い違いを、行動は意識的に開始されないという証拠として見なした(Banks 2002, Libet 1985, Libet et al. 1983a参照)。多くの人々は、これらの結果を、自由意志への挑戦と捉えている。その行動が自由になされるためには、意識的な意図が行動の開始を駆動し、したがって先行しなければならないという想定に基づいているからである。リベットのRPのタイミングに関する基本的な神経生理学的発見は精査に耐えてきたが(Haggard & Eimer 1999, Matsuhashi & Hallett 2008, Trevena & Miller 2002)、彼の解釈は広く批判されてきた。例えば、リベットのデータは、RPが常に運動に続くかどうか、したがって、それが一般的な準備信号や意図に関連する信号とは対照的に、本当に運動開始を反映しているかどうかを決定することを可能にしない(Mele 2006, Roskies 2010)。Haggard & Eimerは、時間的相関を用いて、リベットらが同定した予期的脳プロセスが意図の気づきの根底にある可能性を探っている。彼らの結果は、異なる信号であるLRP(外側化準備電位)が、RPよりも運動意図に関連する脳プロセスのより良い候補であることを示唆している(Haggard & Eimer 1999)。Trevena & Millerは、LRPがRPよりも運動開始とより密接に関連していることに同意し、彼らのデータは、意図の気づきがLRPに先行する可能性を示唆している。他の人々は、リベットの実験デザインが動くという意識的な意図の開始を正確に測定することに失敗しており(Bittner 1996, Lau et al. 2006, Roskies 2010, Young 2006)、代わりに異なる状態を測定しているかもしれないと主張している(Banks 2002)。他の研究は、リベットのパラダイムが意識的な気づきの時間の判断を偏らせる可能性を示唆しており(Lau et al. 2006, 2007)、したがって相対的なタイミングに関する推論は信頼できないかもしれない。(リベットに関するさらなるコメントについては、Banks 2002, Banks & Pockett 2007, Mele 2009, Pacherie 2006, Sinnott-Armstrong & Nadel 2010を参照。)要するに、リベットの研究は、たとえそれがある種の反復的な運動課題において、個々の運動が意識的に開始されないかもしれないことを示唆するとしても、人間の自由という一般的な概念を覆すにはほとんど役立たない。
用語解説
- LRP: 外側化準備電位(lateralized readiness potential)
最近のあるイベント関連研究は、運動意図のタイミングを調べるもので、Haynesらは、前頭極および頭頂皮質の領域からのfMRIデータに対してパターン分類技術を用い、運動決定を予測した。驚くべきことに、予測を助ける情報は、決定が意識的になされる7〜10秒前に利用可能であったが、気づきの前の予測成功率は偶然(約60%)よりわずかに高かっただけである(Soon et al. 2008)。この研究は、先行する脳の状態、おそらくは無意識的なものが、意思決定に影響を与えたり、偏らせたりすることができることを示している。決定や行動への神経的な先行要因や行動への物理的な影響は、物理的に具現化された認知システムからは期待されることであるが、任意の脳情報が、それらがなされるずっと前に将来の任意の決定に対して多くのガイダンスを提供できることは驚くべきことである。この研究の弱い予測的成功は、私たちの意志や自由の概念を覆すものではないが、それにもかかわらず、選択に関する通常の考え方に対して重要な課題を提起する。
ほとんどの神経科学的研究は、抽象的な人間の意図に明確に焦点を当てていない。その一部は、それらを客観的に測定する方法を見つけることが非常に難しいからである。ある研究で、Lauら(2004a)は、被験者に、自分の意図のタイミングまたは運動そのものに注意を向けながら、意のままにボタンを押すように指示した。意図への注意は、運動への注意と比較して、preSMA、DLPFC、およびIPS(頭頂間溝)におけるBOLD(血液酸素化レベル依存)信号の増加につながった。特定の認知課題の側面への注意が、それらの側面を処理することに関与する領域への血流を増加させることを示す多くのイメージング結果に依拠して(Corbetta et al. 1990, O’Craven et al. 1997)、彼らは、運動意図がpreSMAで表象されることを示すものとして、自分たちの結果を解釈した。これらの結果は、自己開始された運動活動につながる近位の意図がpreSMAで表象されるという見方と一致するだけでなく、意識的な意図もまたそこで表象されるという見方とも一致する。
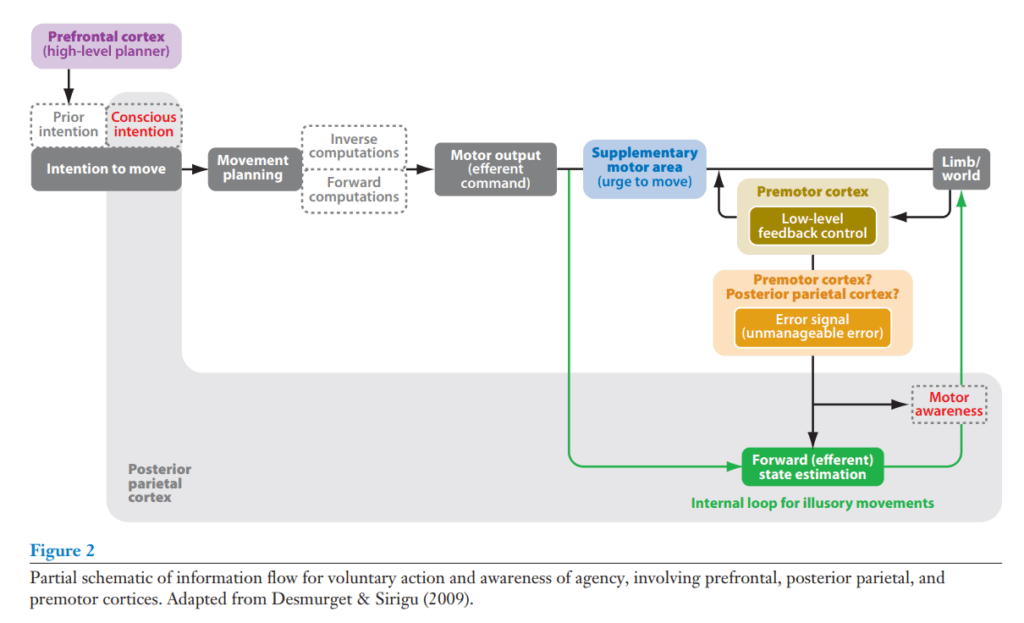
preSMAに加えて、Lauの研究は、しばしば意図的行動に関与する前頭および頭頂領域を強調した(図2)。Hesseら(2006)は、運動計画における前頭頭頂ネットワークを同定しており、これには左縁上回、IPS、および前頭領域が含まれる。左前部IPSもまた、運動計画に重要な目標表象と関連付けられている(Hamilton & Grafton 2006)。これらの結果は、後部頭頂領域が運動意図を表象するという見方と一致する(Andersen & Buneo 2003, Cui & Andersen 2007, Quian Quiroga et al. 2006, Thoenissen et al. 2002)。Siriguら(2004)は、頭頂皮質への損傷が、自発的行動は妨げられないものの、行動する意図の気づきを妨害すると報告している。意図の経験におけるPPC(後部頭頂皮質)の役割は、後のセクションでさらに議論される。
用語解説
- BOLD: 血液酸素化レベル依存(blood oxygenation level dependent)
- IPS: 頭頂間溝(interparietal sulcus)
- PPC: 後部頭頂皮質(posterior parietal cortex)
しばしば、私たちは意図を、運動活動とより密接に関連しない、より抽象的な計画として考える。多くの研究は、背側前頭前野(DPFC)が意図された行動を伴う課題で活動的であることを示している。DPFCの内側部分は、自分自身の意図について考えることに関与しているかもしれないが(den Ouden et al. 2005)、DLPFCは認知応答および運動応答の生成に関与しているかもしれない(Frith et al. 1991, Hyder et al. 1997, Jenkins et al. 2000, Lau et al. 2004a)。しかし、観察された活動が、選択、制御、または行動への注意に対応するかどうかを決定することは困難である。Lauら(2004b)は、自由応答課題において、作業記憶と注意を制御しようとし、どの領域が行動の選択に関与しているかを決定しようとした。DLPFCは、自由選択条件では外的に指定された選択条件よりも活動的ではなかった。これは、それが選択よりも選択への注意とより関係があることを示唆している。対照的に、preSMAは他の条件よりも自由選択でより活動的であった。これは、preSMAが行動の自由選択に関与しているというさらなる証拠を提供する。さらに、選択への注意はDLPFCを伴う。意図の気づきには注意が必要かもしれないので、DLPFCの活動は意識的な意図を反映しているかもしれない。
これまでのところ、議論された領域は、意図の内容についてはほとんど明らかにしていない。前頭前野および頭頂皮質の領域からのfMRIデータに対するパターン分析を用いて、Haynesらは、被験者が数字を足すか引くかという意識的だが内密の意図を、最大70%の精度で予測することができた(Haynes et al. 2007)。特定の意図に関連する情報は、したがって、被験者が意図した行動を心に留めている間、これらの領域(内側、外側、および前頭極前頭領域を含む)に存在する。予測的な領域は、一般的に意図の表象や内生的行動に関与するものとは異なっているように見え、意図に関連する情報が課題に応じて異なって表象される可能性を提起する。
今日まで、神経科学は、内生的な開始と行動の選択の根底にあるメカニズムが、主に神経イベントと気づきの相対的なタイミングに関して、意志の常識的な概念から逸脱するいくつかの特徴を持つことを示してきた。特定の文脈では、選択と運動意図の神経メカニズムが無意識的に活性化されるかもしれないが、意図が作動する多様なレベルを考慮に入れると(Mele 2009, Pacherie 2006, Roskies 2010)、現在のデータはいずれも意志や自由意志の基本的な概念を覆すものではない。人間の自由の死に関する報告は、大いに誇張されている。
意志としての意思決定(Volition as Decision-Making)
ある有力な見解では、意志の典型的な行使は、将来の行動を開始したり表象したりするのではなく、どのような行動方針をとるかを選択する私たちの能力にある。多くの哲学者は、自由意志の自由を、どの意図を形成するかを自由に選択する能力に位置づけてきた。決定は、しばしば意図と開始に先行する。
霊長類の神経生理学の研究者たちは、単一細胞記録と集団モデリングを用いて、知覚的意思決定のダイナミクスの豊かな全体像を構築している。その結束性と幅広さのために、私はウィリアム・ニューサム、マイケル・シャドレン、および同僚たちの研究室からの一連の研究に集中する。彼らは、視覚運動パラダイムを用いて、不確実性下での意思決定の神経基盤を詳細に解明してきた。この研究は他で広範にレビューされている(Glimcher 2001, 2003; Gold & Shadlen 2007)。ここでは主要な発見を簡潔に要約する。
用語解説
- サッケード(Saccade): 視野のある一点から別の点へ目がジャンプする眼球運動。
- LIP: 外側頭頂間野(lateral interparietal area)
これらの研究は共通のパラダイムを共有している:アカゲザルはランダムドット運動ディスプレイを見る。サルの課題は、刺激を固視し、正味の運動方向を決定し、固視点の右と左に置かれた2つのターゲットの一方に正味の運動方向にサッケードを行うことによって、コヒーレントな運動の方向を示すことである。特定の方向への成分運動ベクトルを持つドットの割合を変更することで、課題は多かれ少なかれ難しくなり、それによって正味の運動のコヒーレンス(または強度)が変化する。課題遂行中に異なる脳領域の細胞から記録することにより、決定へのニューロンの貢献が解明されうる。
視覚野MT(中側頭野)およびMST(内側上側頭野)の細胞は、特定の方向の運動に同調している。受容野が視覚刺激の位置と一致するこれらの領域の細胞から記録すると、それらの神経活動が、細胞の好む運動方向への瞬間的な運動信号の強度を反映していることが示される(Britten et al. 1992, Celebrini & Newsome 1994, Newsome et al. 1989)。領域LIP(外側頭頂間野)の神経活動は、異なるプロファイルを示す。LIPのニューロンは、視覚情報と運動情報の両方を表象する(Shadlen & Newsome 1996)。LIP細胞は、線条体外運動野からの信号を時間的に統合するように見える(Huk & Shadlen 2005)。それらはまた、眼球運動の計画と実行においても活動的である(Andersen & Buneo 2002)。
ランダムドット運動課題では、例えば、右への強いコヒーレントな運動信号を持つ刺激は、対応するサッケードターゲットを包含する応答野を持つLIPニューロンの活動の急上昇につながる(Shadlen & Newsome 2001)。発火の増加率は運動強度に比例し、LIPニューロンの活動が特定の絶対レベルに達すると、サルはターゲットにサッケードを行い、発火は停止する(Roitman & Shadlen 2002)。したがって、LIPニューロンは、特定の閾値に達するまで、感覚野からの運動強度の証拠を蓄積するように見える。そして決定がなされる(Huk & Shadlen 2005)。この解釈は、もしサルが手がかりを与えられるまで応答を保留するように訓練されると、計画された応答方向の応答野を持つLIPニューロンは遅延期間中に高い発火率を維持し、サッケード後にのみその活動を停止するという発見によって強化される。したがって、MTやMSTのニューロンとは異なり、これらのニューロンは純粋に刺激駆動型ではなく、刺激がない状態での継続的な発火は、課題の完了まで、刺激の運動方向に関するサルの「決定」の維持を反映していると解釈される(Shadlen & Newsome 2001)。これらのニューロンの活動は、正しい試行と誤った試行の両方で、サルの応答を予測する(Shadlen & Newsome 2001)。他の実験は、意思決定プロセスにおけるLIPニューロンの因果的関与の証拠を提供する。例えば、サッケードターゲットに対応する応答野を持つLIPニューロンの微小刺激は、サルの選択を偏らせ、その応答のタイミングに影響を与える(Hanks et al. 2006)。
これらの神経プロセスは、数学的にモデル化され、サルが示す行動パターンをよく捉える、競争-閾値モデルまたは蓄積-境界モデルに組み込まれている(Gold & Shadlen 2007, Mazurek et al. 2003)。人間における心理物理学的研究は、サルと人間がこの課題を同様に遂行することを示しており、私たちのこれらの運動刺激に関する決定には類似の神経プロセスが関与していることを示唆している(Palmer et al. 2005)。
しかし、運動刺激に基づく決定を下す課題が、私たちが通常関心を持つ種類の決定、特に意思決定を意志の現れとして考えるときに、多くを語るものかどうかを問うかもしれない。結局のところ、(a) 道徳的責任が適用され、自発性の概念が重要である種類の決定は、価値、結果、理由、感情など、この単純な知覚システムが行うよりもはるかに複雑な考慮を伴う、と主張するかもしれないし、(b) この課題の刺激駆動的な性質こそが、私たちが意志によって意味するものではない、と主張するかもしれない。
これらの考慮に照らして、このパラダイムが自発的選択のモデルとして機能するにはあまりにも貧弱であるように思われる。
これらの研究をより一般的な方法で捉え、このモデルがどのようにして人間の意思決定のモデルの核として機能しうるかを想像しやすくする方法がある。これらのサル研究では、特定の応答特性を持つニューロンの集団が、サルが課題で下すことができる選択を表象し、それらの相対的な発火率は、決定に至るプロセス中にそれらに与えられる重みを表象するように見える。私たちは、これらのニューロン集団を、「右/左への運動」あるいは「右/左のターゲットへの目の移動」といった、異なる仮説や代替案を表象するものとして考えるかもしれない。もしこれが正確であれば、これらのニューロンは決定の代替案に対応する命題的内容を持つ表象の一部である(Gold & Shadlen 2007)。異なる神経集団が他の命題を表象できると想像することは難しくなく、私たちは現在、命題的内容が神経系でどのように表象されるかを構想するための一般的な枠組みを欠いているが、私たちはそれができることを知っている、なぜなら私たちはそれを表象するからである。一度、抽象的な命題の神経表象を構想できれば、それらを、行動のための理由や考慮事項を表象するものとして、そしてそれらの相対的な発火率を、決定や行動の理由に与えられる重みを反映するものとして考えるのは、ほんの小さな一歩である。私たちが自由な行動、あるいは道徳的に責任を負う行動について考えるとき、それらの行動は通常、理由への応答として私たちが下す決定であるか、あるいはそれに基づいている(Fischer & Ravizza 1998)。
さらに、多くの研究がこのパラダイムを新しい方法で拡張し、一般的なパラダイムが、人間の意思決定に関わる、より豊かで、よりニュアンスに富んだ、抽象的な考慮事項をどのように組み込むことができるかを示唆している。例えば、視覚運動課題での決定に関連するLIPのニューロンの発火率は、結果の期待値とその確率にも影響され、これらは決定計算において役割を果たす(Platt & Glimcher 1999, Yang & Shadlen 2007)。したがって、より高い報酬に関連する結果(決定)はより重く重み付けされ、閾値への上昇の時間経過は、より高い見返りのある結果や、動物がより起こりやすいと期待するようになった結果に対してより速く起こる。これらのニューロンの発火は、古典的な決定理論によって認識され、主観的価値の概念に具現化された意思決定の多くの側面をコード化するように見える(Dorris & Glimcher 2004, Glimcher 2001, Platt & Glimcher 1999)。選択肢の数が増えても同様の計算が行われ、この種のモデルが複数の結果を持つ決定に一般化できることを示唆している(Churchland et al. 2008)。このシステムは、したがって、最も微妙でニュアンスに富んだ種類の人間の意思決定の中心となる主要な要素を概念化するための基本的な枠組みを提供する。
ほとんどの試行では、ランダムドット運動課題は知覚的に駆動される:刺激自体の性質が正しい選択を特定する。しかし、決定は通常、不確実性の条件下でなされ、いくつかの試行では、 stimulusは決定的な証拠を提供しない。サルは時折、0%のコヒーレントな運動を持つランダムドット運動ディスプレイを提示される。視覚刺激は存在するが、 stimulusの情報は曖昧であり、「正しい」または報酬のある選択とは無関係である。それでも、これらの試行では、サルは右向きと左向きの方向を、0%の運動の同一の動画に応答してさえも、一見ランダムに選択する。したがって、サルの選択は完全に外部刺激によって駆動されるのではなく、むしろサル自身の内部の要因によって駆動される。そして、代替的な選択を表象する集団の活動レベルはほぼ均等に一致しているが、LIPの活動の小さな変動と、一方または他方の方向へのサルの最終的な応答との間に、わずかな相関が見出される(Britten et al. 1996, Shadlen et al. 1996)。これは、応答が実際にこれらの神経集団間の競争によって駆動されることを示唆している。
一部の者は、これらの0%運動の場合でさえ、神経発火レベルと選択の間の相関の存在を決定論の証拠と見なすかもしれないが、他の者は、刺激に依存しない変動を意思決定におけるランダムなノイズの存在と有効性の証拠と見なすかもしれない。先に述べた理由から、どちらの立場も正当化されないと私は思う。ある人のノイズは別の人の信号であり、システムのすべての神経入力から記録することができなければ、そのような活動が本当に神経発火の確率的変動性によるものなのか、あるいは動的に進化するシステムの他の部分からの入力、局所的な進行中の活動、あるいは刺激に関連しない環境入力による活動なのかを判断することはできない。これらの代替案を排除できなければ、これらの変動が非決定論的なプロセスによるものかどうか、そして入力がノイズとして見なされるべきか、単に未同定の信号として見なされるべきかを確かめることはできない。これらの理由から、私たちの現在の知識を考えると、それらをリバタリアン的な自由の根拠として、あるいはその欠如の根拠として指摘することは不十分であるように思われる。
用語解説
- リバタリアン的な自由(Libertarian freedom): 私たちは決定論の偽りのおかげで自由意志を持つという見解。
意思決定の神経基盤に関する研究は、伝統的な自由の問いを裁定する助けにはならないが、もし意思決定の神経基盤の一般的なモデルとして受け取られるならば、それは啓発的である。この研究は、私たちが直感的に期待するすべての基本要素―選択肢の表象、価値、証拠、変化する入力に伴うシステムの経時的な進化の動的な特徴付け、そしてさらには確信度(Kiani & Shadlen 2009)―を組み込んでいる、決定プロセスの比較的に包括的なモデルを提供する。これは特徴付けの第一歩に過ぎず、人間の意思決定との間には関連する違いがある。例えば、このシステムは動物が訓練された課題によって厳密に限定されており、決定と運動準備の神経基盤は密接に関連している(Gold & Shadlen 2000)。もし同じ刺激が使用されるが、決定を示す応答が眼球運動でない場合、証拠は、LIPではなく他の神経集団が運動の方向の決定を表象することを示唆している(Cui & Andersen 2007, Gold & Shadlen 2007)。対照的に、一部の人間の意思決定は、より抽象的なレベルで作動するかもしれない―確かに人間は、具体的な運動表象を必要としない応答がない状況で決定を下すことができる。サルもまた抽象的な決定を下すことができるかどうかは、未解決の問題のままである。さらに、私たちが現在持っている全体像は、いかなる決定プロセス中にも脳が行っていることの、まだ部分的で断片的な見方に過ぎない。他の多くの脳領域も意思決定に貢献している。例えば、DLPFCの神経活動もまた、ランダムドット運動課題におけるサルの決定を予測し(Kim & Shadlen 1999)、応答は期待報酬価値に敏感であった(Leon & Shadlen 1999)。このサルの皮質領域は、上で議論した頭頂領域と相互に接続されており、これらの領域の時間的協調が意思決定において重要である可能性がある(Pesaran et al. 2008)。報酬処理に関与する他の領域も、間違いなく関与している(O’Doherty 2001, Schultz et al. 2000参照)。
決定に関する研究は、意図に関する研究とどのように関連するのか?上で議論したランダムドット運動パラダイムでは、LIPの神経活動を意図と同一視することは魅力的である。その活動は応答の生成と因果的に結びついているように見え、サルが応答を遅延させる必要がある場合、LIPの活動は刺激がない状態で持続する。これは、熟慮と行動の間の時間的ギャップを埋める意図に期待することと全く同じである。しかし、述べたように、LIPの活動はモダリティに特異的であり、特定の運動意図、つまり眼球運動を伴うものを反映しており、非様相的な応答ではない。ほとんどの意図、たとえ人間の動物の多くの意図でさえも、モダリティ特異的な運動プログラムで実現される可能性はある。しかし、明確な運動応答がない将来の行動のための意図を非様相的に表象する手段が存在する可能性もある。例えば、大学を卒業する意図、仕事を探す意図などである。いくつかの証拠は、人間においてDLPFCを応答モダリティから独立した決定と結びつけている(Heekeren et al. 2008)。言語は、人間においてそのような表象を可能にするかもしれない。言語能力が神経計算からどのように生じるかに応じて、サルの神経生理学は、言語的にコード化された意図の性質について洞察を提供する場合もあれば、提供しない場合もあるかもしれない。
人間の意思決定のモデルとしてのいくつかの欠点にもかかわらず、決定に関するサル研究は、私たちに意志をメカニズム的に考えることを促す。一部の哲学者は、決定論ではなく、メカニズムが私たちの決定の根底にあるという認識こそが、自由への最も強力な挑戦であると主張する(Nahmias et al. 2007)。この考えを支持するいくつかの証拠はあるが、メカニズムの脅威と、メカニズムと還元主義の関係については、私たちが理解していないことが多くある。もしメカニズムが自由と相容れないならば、意思決定の根底にあるメカニズムに関する私たちの増大する理解が、自由としての意志の概念を覆すことになるかもしれない。しかし、正しい選択が外部から指示されない場合に選択肢の中から選択する私たちの能力の根底にあるプロセスへの、この研究が提供する洞察を受け入れるように、私たちの自由に関する見解が適応することも、等しく考えられる。
意志としての実行制御(Volition as Executive Control)
意志の制御の側面は、高次の皮質領域が下位の領域による行動の実行に影響を与えることができるという概念である。これはいくつかの形をとるかもしれない。例えば、一つの概念は、意志が行動の意識的な選択を伴うというものである(Bunge 2004, Donohue et al. 2008, Fleming et al. 2009, Hyder et al. 1997, Lau et al. 2004b, Matsumoto et al. 2003, Rowe et al. 2008, Rushworth 2008)。もう一つは、監視が、実行されるにつれて行動がとる形に影響を与えることができるというものである(Barch et al. 2000, Kerns et al. 2004, Ridderinkhof et al. 2004, Schall & Boucher 2007, Schall et al. 2002)。意図された、あるいは計画された行動を抑制する能力を含むものとして制御を考えるのは、ほんの一歩先である(Aron et al. 2007, Brass & Haggard 2007, Brown et al. 2008, Kühn et al. 2009b)。不適切な行動を抑制することによって自分の行動を制御する能力は、法の一部が法的責任にとって重要であると認識しているものである。
前頭前野は一般的に実行制御に関与しているが、前頭前野は大きく不均一な領域であり、前頭下位領域の機能的役割については、まだ多くが決定されていない。前頭前野の一部の領域は、実行制御にとって特に重要であるように見える。多くの研究は、注意制御と課題切り替えにおいて、PFCと頭頂皮質の領域との相互作用を関与させている(Badre 2008; Bode & Haynes 2009; Chiu & Yantis 2009; Dosenbach et al. 2007, 2008; Praamstra et al. 2005; Rossi et al. 2009; Serences & Yantis 2007)。頭頂領域の一部のような、皮質の他の領域は、進行中の行動を導く役割を果たしているように見える(Dosenbach et al. 2007, 2008)。
前頭前野のいくつかの領域は、意志に関する研究で何度も現れる。DLPFCは、選択や意思決定を伴う多くの課題で活性化される(Cunnington et al. 2006, Heekeren et al. 2006, Jahanshahi et al. 1995, Kim & Shadlen 1999, Lau et al. 2004a)。DLPFCは、行動間の選択やルール選択において活性化されるため、抽象的および具体的な決定に関与している(Assad et al. 1998; Bunge 2004; Bunge et al. 2003, 2005; Donohue et al. 2008; Rowe et al. 2008)。先に述べたように、応答選択、意識的熟慮、葛藤解決を含む、選択と行動選択を伴う課題におけるDLPFCの役割について、競合する仮説が存在する。いくつかの研究は、DLPFCの活動が行動選択への注意を反映している(そして、おそらくは意識的制御)ことを示唆しているが(Lau et al. 2004b)、他の研究は、DLPFCの活性化が常に意識的経路と関連付けられるわけではないことを示している(Lau & Passingham 2007)。DLPFCはまた、人間におけるより抽象的な形の制御にも関与している。例えば、Knoch & Fehr(2007)のrTMS研究は、誘惑に抵抗する能力が右DLPFCに依存することを示している。
自発的な抑制的行動制御を支えるネットワークを識別することは、より直接的であるように見える。リベットは、彼の実験的証拠に基づいて、意識的意図は行動を生み出す上で因果的に有効ではないと主張し、RPと行動の間の遅れが、無意識的に生成された行動の抑制を可能にし、それによって自由意志の精神を「自由な不作為(free won’t)」で保存するかもしれないという見解で自らを慰めた(Libet et al. 1983b)(図1)。しかし、彼はこれを純粋な推測として残した。より最近の研究は、意図された行動の抑制の神経メカニズムに光を当て始めている(ただし、それらはリベットが「自由な不作為」が本当に自由であるために期待したかもしれない二元論的な風味を欠いている)。例えば、Brass & Haggard(2007)は最近、被験者が応答を意図し、その後ランダムにその応答を抑制することを選択する必要があるリベット様の課題でfMRI実験を行い、前頭内側皮質領域での活動の増加を報告した。彼らは、これらの前頭内側領域が自己生成された行動を自発的に抑制することに関与していると推測している。同様の領域が、優勢な応答を抑制する決定にも関与している(Kühn et al. 2009b)。接続性分析は、内側前頭部の抑制がトップダウン形式でpreSMAに影響を与えることを示唆している(Kühn et al. 2009b)。他の証拠は、抑制がより低いレベルの運動階層、例えば一次運動野の局所的な皮質ネットワークでも起こることを示唆している(Coxon et al. 2006)。
背側内側前頭領域は抑制プロセスに直接関与しているように見えるが、自発的な行動決定を媒介する同じ領域が、行動を控えるという自発的な決定にも関与しているように見える。ERPとfMRIの両方の研究からの証拠は、意図的に行動しない、あるいは行動する意図を形成した後に行動しないと決定することの神経的兆候が、行動する決定のそれと非常によく似ていることを示している(Kühn & Brass 2009b, Kühn et al. 2009a)。例えば、前部帯状皮質と背側preSMAの領域は、自由に選択されたボタン押しと、ボタンを押さないという自由な決定の両方で活動的である。行動する決定と行動を控える決定との間の類似した神経基盤は、行動と不作為の両方が私たちが責任を負うことができる意志の行為であるという常識的な概念に信憑性を与える。
感覚としての意志(Volition as a Feeling)
意欲の経験は、多面的な意志能力の一側面である。一部の者は、経験が説明すべき全てであると考えている。なぜなら、それは他の点では幻想的な意志の経験だからである(Wegner 2002)。主体性には少なくとも2つの現象学的側面がある:私たちが行動に先立つものとして同定する、行動する意図や衝動の気づきと、とられた行動が自分のものであったという事後的な感覚である。
第一の点に関して、最近の結果は、自発的意図の経験が頭頂皮質に依存することを明らかにしている(図2)。この領域への電気刺激は運動意図を誘発し、より強い刺激は時として運動が起こったという誤った信念につながった(Desmurget et al. 1999)。対照的に、運動前野皮質の刺激は、運動の気づきなしに運動につながった(Desmurget et al. 2009)。これは、主体性の気づきが運動前野領域よりも主に頭頂部に依存することを示唆しているが、Friedらは、SMAへの刺激もまた動きたいという欲求を誘発したと報告している。SMAへの刺激によって誘発された意図は、頭頂部刺激によって誘発されたものとは対照的に、自発的意図よりも強迫観念の現象学を持っていた(Fried et al. 1991)。Desmurgetらが、前頭前野の刺激が感じられた意図を誘発しなかったことを見出さなかったとしても、彼らが刺激したBA6の部位は、Friedらが刺激した領域よりも外側にある傾向があった。加えて、下頭頂葉の損傷は、運動意図のタイミングの気づきを変化させる。運動前に意図に気づくようになる代わりに、これらの損傷患者は、運動の直前にのみ気づきを報告した(Sirigu et al. 2004)。対照的に、運動タイミングを正確に報告する彼らの能力は損なわれなかった。
主体性の感覚または行動の所有権の生成に関与する神経信号を同定することにおいても、かなりの進歩が見られている(図2)。主体性の感覚は、行動の効果からの自己受容感覚的および知覚的フィードバックの両方に依存するように見える(Kühn & Brass 2009a, Moore & Haggard 2008, Moore et al. 2009, Pacherie 2008, Tsakiris et al. 2005)。多くの研究は、行動計画にはしばしば、システムがさらなる感覚フィードバックの期待を形成することを可能にする遠心性信号が伴うことを示している。この期待が裏切られなければ、主体性の感覚に貢献する(Linser & Goschke 2007, Sirigu et al. 2004)。Graftonらは、予測された運動結果と実際の運動結果との間に不一致がある場合に、右角回(下頭頂皮質)での活動、および所有権の気づき(Farrer et al. 2008)を見出した。順行モデルの予測が感覚情報または自己受容情報と一致するときの頭頂皮質からの信号は、主体性の感覚を生み出す上で重要かもしれない。さらに、主体性の気づきの一部の側面は、遡及的に構築されるように見える。最近の研究は、人々の意図形成の時期に関する判断が、時間シフトした感覚フィードバックによって変更されうることを示している。これは、意図の気づきが、直接知覚されるのではなく、少なくとも部分的に応答から推測されることを示唆している(Banks & Isham 2009)。期待もまた役割を果たすことができる(Voss et al. 2008)。これらの研究は、リベットの測定パラダイムが報告された意図の気づきの時間に影響を与えるかもしれないという批判に信憑性を与える(Lau et al. 2006, 2007)。加えて、効果に対する行動の知覚された開始は、行為者がその行動を意欲的であると知覚するかどうかによって調節される(Engbert et al. 2008, Haggard 2008)。
述べたように、前頭領域もまた意図の気づきに貢献するかもしれない。Friedの刺激研究は、SMA(おそらくpre-SMA)の領域の刺激が意図の気づきにつながることを示した(Fried et al. 1991)。行動実行後のSMA上のTMSもまた、報告された意図の気づきの時間に影響を与える(Lau et al. 2007)。これは、意図の気づきが部分的に再構築であるというさらなる証拠である。
これらの結果は、頭頂皮質が運動意図と、自発的行動中の行動のための予測信号または順行モデルを生成するモデルと一致する(図2)。運動計画は実行のために前頭領域に中継され、これらの領域の活性化は意図とタイミングの気づきの側面にとって重要であるかもしれない。同時に、頭頂領域は内部予測を感覚フィードバックと比較する[ただし、最近の有望なモデルは、比較器が運動前野皮質にあることを示唆している(Desmurget & Sirigu 2009)]。フィードバック信号だけでは、所有権の感覚には不十分である(Tsakiris et al. 2005)。信号が一致する場合、私たちは運動意図に気づかないままであるかもしれないが(Sirigu et al. 2004)、行動を自分自身のものとして知覚する。私たちは、順行モデルと知覚からの情報との間に不一致が検出されたときにのみ、運動意図に気づかされるのかもしれない。したがって、遠心性の内部モデルと環境からのフィードバックの両方が、主体性の知覚と自己所有権において重要である(Moore et al. 2009)。
通常の状況下では、私たちは自発的行動を自発的なものとして経験する。異常な状況下では、人々は誤って主体性を自分自身に帰属させたり、帰属させなかったりすることがある(Wegner 2002, Wegner & Wheatley 1999)。主体性の感覚が、意志が因果的に有効であるというのは単なる幻想であると示唆する者もいる(Hallett 2007, Wegner 2002)。しかし、主体性の経験が常に真実を反映しているわけではないとしても、一般に、主体性の感覚が実際の主体性を反映していない、意志が因果的に有効ではない、あるいは自由意志が感覚以上のものではないと結論づけるべきではない。意志の経験が神経的な基盤を持つという単なる事実もまた、意志の自由を否定する根拠にはならない。主体性の経験を媒介する回路と、運動の開始、意図の形成などに関与する回路との相互作用をよりよく理解することは、意志のこれらの様々な側面がどのように関連し、特定の形態の脳損傷や、特定の外部イベントの配置によって、どのように解離しうるかを説明するかもしれない。
結論
全体として、神経科学は私たちの意志の概念にあまり影響を与えていない。それは、意図、選択、そして主体性の経験という概念を大部分維持してきた。神経科学が私たちの概念に影響を与えたところでは、それは典型的には、意識と行動の関係に関する伝統的な見解に挑戦してきた。例えば、以前考えられていたよりも多くの行動の側面が無意識的なプロセスによって支配されている。しかし、私たちは意識の神経基盤についてほとんど牽引力を持っていないので、それらの挑戦のいずれも、私の考えでは、伝統的な見解を覆すことに成功していない。しかし、神経科学は、意志が単一の能力ではなく、むしろ、柔軟で知的な行動を可能にする、大部分分離可能なプロセスの集合であることを示すことを約束している。それは、将来、これらの異なる意志の側面を支える神経システムと計算を解明することによって、私たちの意志の概念に影響を与えるかもしれない。脳ネットワークのさらなる解明は、意志の要素を分類するためのより良い方法を提供するかもしれない(Brass & Haggard 2008; Pacherie 2006, 2008)。神経科学が決定論の問題への正面攻撃によって自由の問題に関わることはないと私は信じているが、これらのプロセスの神経基盤に関する私たちの理解を深めることは、私たちが現在よりも意志をよりメカニズム的に考えるようにさせるかもしれず、それが最終的には、自由のために何が必要かという私たちの通常の概念に圧力をかけることになるかもしれない。しかし、今のところ、神経科学がもたらした最も重要な貢献は、自発的行動の性質に関する新しい問いを定式化することを可能にし、それらに取り組むための新しい方法を提供することであった。
要約ポイント
- 私たちが意志として考えるものは、単一の能力ではないかもしれない。
- 神経科学は、決定論の問題を解決しないだろう。
- 前頭および頭頂領域のネットワークが、自発的行動の開始、選択、および制御に関与している。
- PreSMAとrCZは、内生的に生成される運動に関与している。
- 意思決定の多くの側面の神経基盤は、よく理解されており、数学的にモデル化できる。
- 行動する選択と行動を控える選択は、類似の脳回路を伴うように見える。
- 主体性の感覚は、部分的に頭頂皮質によって媒介される。それは、予測的信号と事後的フィードバックの両方に依存する。
- これまでのところ、意識的意図のタイミング、メカニズム、あるいは主体性の幻想のいずれも、意志の存在や有効性を覆すものではない。
今後の課題
- メカニズムが意志と自由の説明にもたらす挑戦とは何か?
- 選択、計画、行動開始、制御、および主体性の感覚を媒介する異なる回路は、互いにどのように相互作用するのか?
- それらの回路は、自己の表象をどのように関与させ、根底に持ち、相互作用するのか?
- preSMAとrCZにおける自発的行動で同定された信号の根底にある神経計算は何か、そしてそれらは行動開始においてどのような役割を果たすのか?
- どのようなプロセスが意思決定における閾値および/またはベースライン活動を設定するのか?
- 内部ループは、刺激に束縛されない行動をどのように可能にするのか?
- 前頭領域は、他の脳領域の神経活動をどのように制御、調節、および修正するのか?
- 神経系における変動性の源は何か?ノイズは選択と行動においてどのような役割を果たすのか?
開示声明
著者は、このレビューの客観性に影響を与えると認識されうる、いかなる所属、メンバーシップ、資金提供、または金銭的利害関係も認識していない。
謝辞
この研究は、ジョンズ・ホプキンス大学バーマン生命倫理研究所へのNEH共同研究助成金、および法と神経科学におけるマッカーサープロジェクトによって部分的に支援された。初期の草稿に対するコメントについて、エラン・クライン、ナンシー・マコーネル、アル・メレ、ショーン・ニコルズ、マイケル・シャドレン、およびウォルター・シノット=アームストロングに感謝したい。
アディーナ・ロスキーズの論文「How Does Neuroscience Affect Our Conception of Volition?」の最後に記載されている参考文献(LITERATURE CITED)のリストを、見やすく整理して提示します。
参考文献リスト
A
- Adams F, Mele A. 1992. The intention/volition debate. Can. J. Philos. 22:323–38
- Andersen RA, Buneo CA. 2002. Intentional maps in posterior parietal cortex. Annu. Rev. Neurosci. 25:189–220
- Andersen RA, Buneo CA. 2003. Sensorimotor integration in posterior parietal cortex. Adv. Neurol. 93:159–77
- Aron AR, Behrens TE, Smith S, Frank MJ, Poldrack RA. 2007. Triangulating a cognitive control network using diffusion-weighted magnetic resonance imaging (MRI) and functional MRI. J. Neurosci. 27:3743–52
- Assad WF, Rainer G, Miller EK. 1998. Neural activity in the primate prefrontal cortex during associative learning. Neuron 21:1399–407
- Audi R. 1993. Volition and agency. In Action, Intention, and Reason, ed. AR Mele, pp. 74–108. Ithaca, NY: Cornell Univ. Press
B
- Badre D. 2008. Cognitive control, hierarchy, and the rostro-caudal organization of the frontal lobes. Trends Cogn. Sci. 12:193–200
- Banks WP, ed. 2002. Consciousness and Cognition, Vol. 11. New York: Academic Press
- Banks WP, Isham EA. 2009. We infer rather than perceive the moment we decided to act. Psychol. Sci. 20:17–21
- Banks WP, Pockett S. 2007. Benjamin Libet’s work on the neuroscience of free will. In Blackwell Companion to Consciousness, ed. M Velmans, S Schneider, pp. 657–70. Malden, MA: Blackwell
- Barch DM, Braver TS, Sabb FW, Noll DC. 2000. Anterior cingulate and the monitoring of response conflict: evidence from an fMRI study of overt verb generation. J. Cogn. Neurosci. 12:298–309
- Bittner T. 1996. Consciousness and the act of will. Philos. Stud. 81:331–41
- Bode S, Haynes JD. 2009. Decoding sequential stages of task preparation in the human brain. NeuroImage 45:606–13
- Brass M, Haggard P. 2007. To do or not to do: the neural signature of self-control. J. Neurosci. 27:9141–45
- Brass M, Haggard P. 2008. The what, when, whether model of intentional action. Neuroscientist 14:319–25
- Britten KH, Newsome WT, Shadlen MN, Celebrini S, Movshon JA. 1996. A relationship between behavioral choice and the visual responses of neurons in macaque MT. Vis. Neurosci. 13:87–100
- Britten KH, Shadlen MN, Newsome WT, Movshon JA. 1992. The analysis of visual motion: a comparison of neuronal and psychophysical performance. J. Neurosci. 12:4745–65
- Brown JW, Hanes DP, Schall JD, Stuphorn V. 2008. Relation of frontal eye field activity to saccade initiation during a countermanding task. Exp. Brain Res. 190:135–51
- Bunge SA. 2004. How we use rules to select actions: a review of evidence from cognitive neuroscience. Cogn. Affect. Behav. Neurosci. 4:564–79
- Bunge SA, Kahn I, Wallis JD, Miller EK, Wagner AD. 2003. Neural circuits subserving the retrieval and maintenance of abstract rules. J. Neurophysiol. 90:3419–28
- Bunge SA, Wallis JD, Parker A, Brass M, Crone EA, et al. 2005. Neural circuitry underlying rule use in humans and nonhuman primates. J. Neurosci. 25:10347–50
C
- Celebrini S, Newsome WT. 1994. Neuronal and psychophysical sensitivity to motion signals in extrastriate area MST of the macaque monkey. J. Neurosci. 14:4109–24
- Chiu Y-C, Yantis S. 2009. A domain-independent source of cognitive control for task sets: shifting spatial attention and switching categorization rules. J. Neurosci. 29:3930–38
- Churchland AK, Kiani R, Shadlen MN. 2008. Decision-making with multiple alternatives. Nat. Neurosci. 11:693–702
- Ciaramidaro A, Adenzato M, Enrici I, Erk S, Pia L, et al. 2007. The intentional network: how the brain reads varieties of intentions. Neuropsychologia 45:3105–13
- Corbetta M, Miezin FM, Dobmeyer S, Shulman GL, Petersen SE. 1990. Attentional modulation of neural processing of shape, color, and velocity in humans. Science 248:1556–59
- Coxon JP, Stinear CM, Byblow WD. 2006. Intracortical inhibition during volitional inhibition of prepared action. J. Neurophysiol. 95:3371–83
- Cui H, Andersen RA. 2007. Posterior parietal cortex encodes autonomously selected motor plans. Neuron 56:552–59
- Cunnington R, Windischberger C, Deecke L, Moser E. 2002. The preparation and execution of self-initiated and externally-triggered movement: a study of event-related fMRI. NeuroImage 15:373–85
- Cunnington R, Windischberger C, Robinson S, Moser E. 2006. The selection of intended actions and the observation of others’ actions: a time-resolved fMRI study. NeuroImage 29:1294–302
D
- Deecke L, Kornhuber HH. 1978. An electrical sign of participation of the mesial ‘supplementary’ motor cortex in human voluntary finger movement. Brain Res. 159:473–76
- Deiber M-P, Honda M, Ibanez V, Sadato N, Hallett M. 1999. Mesial motor areas in self-initiated versus externally triggered movements examined with fMRI: effect of movement type and rate. J. Neurophysiol. 81:3065–77
- de Lange FP, Spronk M, Willems RM, Toni I, Bekkering H. 2008. Complementary systems for understanding action intentions. Curr. Biol. 18:454–57
- den Ouden HE, Frith U, Frith C, Blakemore SJ. 2005. Thinking about intentions. NeuroImage 28:787–96
- Desmurget M, Epstein CM, Turner RS, Prablanc C, Alexander GE, Grafton ST. 1999. Role of the posterior parietal cortex in updating movements to a visual target. Nat. Neurosci. 2:563–67
- Desmurget M, Reilly KT, Richard N, Szathmari A, Mottolese C, Sirigu A. 2009. Movement intention after parietal cortex stimulation in humans. Science 324:811–13
- Desmurget M, Sirigu A. 2009. A parietal-premotor network for movement intention and motor awareness. Trends Cogn. Sci. 13:411–19
- Donohue SE, Wendelken C, Bunge SA. 2008. Neural correlates of preparation for action selection as a function of specific task demands. J. Cogn. Neurosci. 20:694–706
- Dorris MC, Glimcher PW. 2004. Activity in posterior parietal cortex is correlated with the relative subjective desirability of action. Neuron 44:365–78
- Dorval AD. 2006. The rhythmic consequences of ion channel stochasticity. Neuroscientist 12:442–48
- Dosenbach NUF, Fair DA, Cohen AL, Schlaggar BL, Petersen SE. 2008. A dual-networks architecture of top-down control. Trends Cogn. Sci. 12:99–105
- Dosenbach NUF, Fair DA, Miezin FM, Cohen AL, Wenger KK, et al. 2007. Distinct brain networks for adaptive and stable task control in humans. Proc. Natl. Acad. Sci. USA 104:11073–78
E
- Engbert K, Wohlschlager A, Haggard P. 2008. Who is causing what? The sense of agency is relational and efferent-triggered. Cognition 107:693–704
F
- Farrer C, Frey SH, Van Horn JD, Tunik E, Turk D, et al. 2008. The angular gyrus computes action awareness representations. Cereb. Cortex 18:254–61
- Fischer J, Ravizza M. 1998. Responsibility and Control: A Theory of Moral Responsibility. New York: Cambridge Univ. Press
- Fleming SM, Mars RB, Gladwin TE, Haggard P. 2009. When the brain changes its mind: flexibility of action selection in instructed and free choices. Cereb. Cortex 19:2352–50
- Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, Rizzolatti G. 2005. Parietal lobe: from action organization to intention understanding. Science 308:662–67
- Franks KM, Stevens CF, Sejnowski TJ. 2003. Independent sources of quantal variability at single glutamatergic synapses. J. Neurosci. 23:3186–95
- Fried I, Katz A, McCarthy G, Sass K, Williamson P, et al. 1991. Functional organization of human supplementary motor cortex studied by electrical stimulation. J. Neurosci. 11:3656–66
- Frith CD, Friston K, Liddle PF, Frackowiak RSJ. 1991. Willed action and the prefrontal cortex in man: a study with PET. Proc. R. Soc. Lond. Ser. B 244:241–46
G
- Glimcher PW. 2001. Making choices: the neurophysiology of visual-saccadic decision making. Trends Neurosci. 24:654–59
- Glimcher PW. 2003. The neurobiology of visual-saccadic decision making. Annu. Rev. Neurosci. 26:133–79
- Glimcher PW. 2005. Indeterminacy in brain and behavior. Annu. Rev. Psychol. 56:25–56
- Gold JI, Shadlen MN. 2000. Representation of a perceptual decision in developing oculomotor commands. Nature 404:390–94
- Gold JI, Shadlen MN. 2007. The neural basis of decision making. Annu. Rev. Neurosci. 30:535–74
H
- Haggard P. 2005. Conscious intention and motor cognition. Trends Cogn. Sci. 9:290–95
- Haggard P. 2008. Human volition: towards a neuroscience of will. Nat. Rev. Neurosci. 9:934–46
- Haggard P, Eimer M. 1999. On the relation between brain potentials and the awareness of voluntary movements. Exp. Brain Res. 126:128–33
- Hallett M. 2007. Volitional control of movement: the physiology of free will. Clin. Neurophysiol. 118:1179–92
- Hamilton AF, Grafton ST. 2006. Goal representation in human anterior intraparietal sulcus. J. Neurosci. 26:1133–37
- Hanks TD, Ditterich J, Shadlen MN. 2006. Microstimulation of macaque area LIP affects decision-making in a motion discrimination task. Nat. Neurosci. 9:682–89
- Haynes JD, Sakai K, Rees G, Gilbert S, Frith C, Passingham RE. 2007. Reading hidden intentions in the human brain. Curr. Biol. 17:323–28
- Heekeren HR, Marrett S, Ruff DA, Bandettini PA, Ungerleider LG. 2006. Involvement of human left dorsolateral prefrontal cortex in perceptual decision making is independent of response modality. Proc. Natl. Acad. Sci. USA 103:10023–28
- Heekeren HR, Marrett S, Ungerleider LG. 2008. The neural systems that mediate human perceptual decision making. Nat. Rev. Neurosci. 9:467–79
- Hesse MD, Thiel CM, Stephan KE, Fink GR. 2006. The left parietal cortex and motor intention: an eventrelated functional magnetic resonance imaging study. Neuroscience 140:1209–21
- Huk AC, Shadlen MN. 2005. Neural activity in macaque parietal cortex reflects temporal integration of visual motion signals during perceptual decision making. J. Neurosci. 25:10420–36
- Hyder F, Phelps EA, Wiggins CJ, Labar KS, Blamire AM, Shulman RG. 1997. “Willed action”: a functional MRI study of the human prefrontal cortex during a sensorimotor task. Proc. Natl. Acad. Sci. USA 94:6989–94
I
- Iacoboni M, Molnar-Szakacs I, Gallese V, Buccino G, Mazziotta JC, Rizzolatti G. 2005. Grasping the intentions of others with one’s own mirror neuron system. PLoS Biol. 3:e79
J
- Jahanshahi M, Jenkins IH, Brown RG, Marsden CD, Passingham RE, Brooks DJ. 1995. Self-initiated versus externally triggered movements: I. An investigation using measurement of regional cerebral blood flow with PET and movement-related potentials in normal and Parkinson’s disease subjects. Brain 118:913–33
- Jenkins IH, Jahanshahi M, Jueptner M, Passingham RE, Brooks DJ. 2000. Self-initiated versus externally triggered movements: II. The effect of movement predictability on regional cerebral blood flow. Brain 123:1216–28
K
- Kennerley SW, Sakai K, Rushworth MFS. 2004. Organization of action sequences and the role of the Pre-SMA. J. Neurophysiol. 91:978–93
- Kerns JG, Cohen JD, MacDonald AWI, Cho RY, Stenger VA, Carter CS. 2004. Anterior cingulate conflict monitoring and adjustments in control. Science 303:1023–26
- Kiani R, Shadlen MN. 2009. Representation of confidence associated with a decision by neurons in the parietal cortex. Science 324:759–64
- Kim J-N, Shadlen MN. 1999. Neural correlates of a decision in the dorsolateral prefrontal cortex of the macaque. Nat. Neurosci. 2:176–85
- Knoch D, Fehr E. 2007. Resisting the power of temptations: the right prefrontal cortex and self-control. Ann. NY Acad. Sci. 1104:123–34
- Kühn S, Brass M. 2009a. Retrospective construction of the judgement of free choice. Conscious Cogn. 18:12–21
- Kühn S, Brass M. 2009b. When doing nothing is an option: the neural correlates of deciding whether to act or not. NeuroImage 46:1187–93
- Kühn S, Gevers W, Brass M. 2009a. The neural correlates of intending not to do something. J. Neurophysiol. 101:1913–20
- Kühn S, Haggard P, Brass M. 2009b. Intentional inhibition: how the “veto-area” exerts control. Hum. Brain Mapp. 30:2834–43
L
- Lamm C, Fischer MH, Decety J. 2007. Predicting the actions of others taps into one’s own somatosensory representations—a functional MRI study. Neuropsychologia 45:2480–91
- Lau HC, Passingham RE. 2007. Unconscious activation of the cognitive control system in the human prefrontal cortex. J. Neurosci. 27:5805–11
- Lau HC, Rogers RD, Haggard P, Passingham RE. 2004a. Attention to intention. Science 303:1208–10
- Lau HC, Rogers RD, Passingham RE. 2006. On measuring the perceived onsets of spontaneous actions. J. Neurosci. 26:7265–71
- Lau HC, Rogers RD, Passingham RE. 2007. Manipulating the experienced onset of intention after action execution. J. Cogn. Neurosci. 19:81–90
- Lau HC, Rogers RD, Ramnani N, Passingham RE. 2004b. Willed action and attention to the selection of action. NeuroImage 21:1407–15
- Leon MI, Shadlen MN. 1999. Effect of expected reward magnitude on the response of neurons in the dorsolateral prefrontal cortex of the macaque. Neuron 24:415–25
- Libet B. 1985. Unconscious cerebral initiative and the role of conscious will in voluntary action. Behav. Brain Sci. 8:529–66
- Libet B, Gleason CA, Wright EW, Pearl DK. 1983a. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential): the unconscious initation of a freely voluntary act. Brain 106:623–42
- Libet B, Wright EW Jr, Gleason CA. 1982. Readiness-potentials preceding unrestricted “spontaneous” vs preplanned voluntary acts. Electroencephalogr. Clin. Neurophysiol. 54:322–35
- Libet B, Wright EW Jr, Gleason CA. 1983c. Preparation or intention-to-act, in relation to pre-event potentials recorded at the vertex. Electroencephalogr. Clin. Neurophysiol. 56:367–72
- Linser K, Goschke T. 2007. Unconscious modulation of the conscious experience of voluntary control. Cognition 104:459–75
M
- Mainen ZF, Sejnowski TJ. 1995. Reliability of spike timing in neocortical neurons. Science 268:1503–6
- Matsuhashi M, Hallett M. 2008. The timing of the conscious intention to move. Eur. J. Neurosci. 28:2344–51
- Matsumoto K, Suzuki W, Tanaka K. 2003. Neuronal correlates of goal-based motor selection in the prefrontal cortex. Science 301:229–32
- Mazurek ME, Roitman JD, Ditterich J, Shadlen MN. 2003. A role for neural integrators in perceptual decision making. Cereb. Cortex 13:1257–69
- Mele AR. 2006. Free Will and Luck. Oxford: Oxford Univ. Press
- Mele AR. 2009. Effective Intentions: The Power of Conscious Will. New York: Oxford Univ. Press
- Monterosso J, Royzman EB, Schwartz B. 2005. Explaining away responsibility: effects of scientific explanation on perceived culpability. Ethics Behav. 15:139–58
- Moore J, Haggard P. 2008. Awareness of action: Inference and prediction. Conscious Cogn. 17:136–44
- Moore JW, Lagnado D, Deal DC, Haggard P. 2009. Feelings of control: Contingency determines experience of action. Cognition 110:279–83
- Mueller VA, Brass M, Waszak F, Prinz W. 2007. The role of the preSMA and the rostral cingulate zone in internally selected actions. NeuroImage 37:1354–61
N
- Nachev P, Kennard C, Husain M. 2008. Functional role of the supplementary and presupplementary motor areas. Nat. Rev. Neurosci. 9:856–69
- Nachev P, Rees G, Parton A, Kennard C, Husain M. 2005. Volition and conflict in human medial frontal cortex. Curr. Biol. 15:122–28
- Nahmias E, Coates DJ, Kvaran T. 2007. Free will, moral responsibility, and mechanism: experiments on folk intuitions. Midwest Stud. Philos. 31:215–42
- Newsome WT, Britten KH, Movshon JA. 1989. Neuronal correlates of a perceptual decision. Nature 341:52–54
O
- O’Craven KM, Rosen BR, Kwong KK, Treisman AM, Savoy RL. 1997. Voluntary attention modulates fMRI activity in human MT-MST. Neuron 18:591–98
- O’Doherty JEA. 2001. Abstract reward and punishment representations in the human orbitofrontal cortex. Nat. Neurosci. 4:95–102
P
- Pacherie E. 2006. Toward a dynamic theory of intentions. In Does Consciousness Cause Behavior? An Investigation of the Nature of Volition, ed. S Pockett, WP Banks, S Gallagher, pp. 145–67. Cambridge, MA: MIT Press
- Pacherie E. 2008. The phenomenology of action: a conceptual framework. Cognition 107:179–217
- Palmer J, Huk AC, Shadlen MN. 2005. The effect of stimulus strength on the speed and accuracy of a perceptual decision. J. Vis. 5:376–404
- Pesaran B, Nelson MJ, Andersen RA. 2008. Free choice activates a decision circuit between frontal and parietal cortex. Nature 453:406–9
- Picard N, Strick PL. 1996. Motor areas of the medial wall: a review of their location and functional activation. Cereb. Cortex 6:342–53
- Picard N, Strick PL. 2001. Imaging the premotor areas. Curr. Opin. Neurobiol. 11:663–72
- Platt ML, Glimcher PW. 1999. Neural correlates of decision variables in parietal cortex. Nature 400:233–38
- Praamstra P, Boutsen L, Humphreys GW. 2005. Frontoparietal control of spatial attention and motor intention in human EEG. J. Neurophysiol. 94:764–74
Q
- Quian Quiroga R, Snyder LH, Batista AP, Cui H, Andersen RA. 2006. Movement intention is better predicted than attention in the posterior parietal cortex. J. Neurosci. 26:3615–20
R
- Ramnani N, Miall RC. 2004. A system in the human brain for predicting the actions of others. Nat. Neurosci. 7:85–90
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. 2004. The role of the medial frontal cortex in cognitive control. Science 306:443–47
- Rizzolatti G, Sinigaglia C. 2007. Mirror neurons and motor intentionality. Funct. Neurol. 22:205–10
- Roitman JD, Shadlen MN. 2002. Response of neurons in the lateral intraparietal area during a combined visual discrimination reaction time task. J. Neurosci. 22:9475–89
- Roskies AL. 2006. Neuroscientific challenges to free will and responsibility. Trends Cogn. Sci. 10:419–23
- Roskies AL. 2010. Why Libet’s studies don’t pose a threat to free will. In Conscious Will and Responsibility, ed. W Sinnott-Armstrong, L Nadel. New York: Oxford Univ. Press. In press
- Rossi AF, Pessoa L, Desimone R, Ungerleider LG. 2009. The prefrontal cortex and the executive control of attention. Exp. Brain Res. 192:489–97
- Rowe J, Hughes L, Eckstein D, Owen AM. 2008. Rule-selection and action-selection have a shared neuroanatomical basis in the human prefrontal and parietal cortex. Cereb. Cortex 18:2275–85
- Rushworth MF. 2008. Intention, choice, and the medial frontal cortex. Ann. NY Acad. Sci. 1124:181–207
S
- Schall JD, Boucher L. 2007. Executive control of gaze by the frontal lobes. Cogn. Affect. Behav. Neurosci. 7:396–412
- Schall JD, Stuphorn V, Brown JW. 2002. Monitoring and control of action by the frontal lobes. Neuron 36:309–22
- Schultz W, Tremblya L, Hollerman JR. 2000. Reward processing in primate orbitofrontal cortex and basal ganglia. Cereb. Cortex 10:272–83
- Searle JR. 1983. Intentionality: An Essay in the Philosophy of Mind. Cambridge, UK: Cambridge Univ. Press
- Serences JT, Yantis S. 2007. Spatially selective representations of voluntary and stimulus-driven attentional priority in human occipital, parietal, and frontal cortex. Cereb. Cortex 17:284–93
- Shadlen MN, Britten KH, Newsome WT, Movshon JA. 1996. A computational analysis of the relationship between neuronal and behavioral responses to visual motion. J. Neurosci. 16:1486–510
- Shadlen MN, Newsome WT. 1996. Motion perception: seeing and deciding. Proc. Natl. Acad. Sci. USA 93:628–33
- Shadlen MN, Newsome WT. 2001. Neural basis of a perceptual decision in the parietal cortex (area LIP) of the rhesus monkey. J. Neurophysiol. 86:1916–36
- Shibasaki H, Hallett M. 2006. What is the Bereitschaftspotential? Clin. Neurophysiol. 117:2341–56
- Sinnott-Armstrong W, Nadel L, eds. 2010. Conscious Will and Responsibility. New York: Oxford Univ. Press. In press
- Sirigu A, Daprati E, Ciancia S, Giraux P, Nighoghossian N, et al. 2004. Altered awareness of voluntary action after damage to the parietal cortex. Nat. Neurosci. 7:80–84
- Soon CS, Brass M, Heinze H-J, Haynes J-D. 2008. Unconscious determinants of free decisions in the human brain. Nat. Neurosci. 11:543–45
- Sumner P, Nachev P, Morris P, Peters AM, Jackson SR, et al. 2007. Human medial frontal cortex mediates unconscious inhibition of voluntary action. Neuron 54:697–711
T
- Thaler D, Chen YC, Nixon PD, Stern CE, Passingham RE. 1995. The functions of the medial premotor cortex. I. Simple learned movements. Exp. Brain Res. 102:445–60
- Thoenissen D, Zilles K, Toni I. 2002. Differential involvement of parietal and precentral regions in movement preparation and motor intention. J. Neurosci. 22:9024–34
- Trevena JA, Miller J. 2002. Cortical movement preparation before and after a conscious decision to move. Conscious Cogn. 11:162–90
- Tsakiris M, Haggard P, Franck N, Mainy N, Sirigu A. 2005. A specific role for efferent information in self-recognition. Cognition 96:215–31
V
- Voss M, Ingram JN, Wolpert DM, Haggard P. 2008. Mere expectation to move causes attenuation of sensory signals. PloS ONE 3:e2866
W
- Wegner D. 2002. The Illusion of Conscious Will. Cambridge, MA: MIT Press
- Wegner D, Wheatley T. 1999. Apparent mental causation: sources of the experience of will. Am. Psychol. 54:480–92
Y
- Yang T, Shadlen MN. 2007. Probabilistic reasoning by neurons. Nature 447:1075–80
- Young G. 2006. Preserving the role of conscious decision making in the initiation of intentional action. J. Conscious Stud. 13:51–68
Z
- Zhu J. 2004a. Intention and volition. Can. J. Philos. 34:175–93
- Zhu J. 2004b. Understanding volition. Philos. Psychol. 17:247–73
