
4.ヒト脳拡大におけるアロメトリー成長曲線と制約
脳の大きさは種によって大きく異なり、哺乳類の中でも違いがあります。
最も小さな哺乳類の脳は体重に対して非常に軽く、最大の哺乳類(例えばクジラ)では脳の重さが10kgにも達することがあります。
ただし、哺乳類において脳の重さは一般に体重の関数(function of body weight)であり、体重が増えるに伴い脳の重さも増加します。
この関係は、基礎代謝エネルギー消費(resting energy turnover)に似ており、体重の約3/4乗に比例して増加します。
しかし、体重と脳重量を対数グラフにプロットすると、種間で相対的な脳の重さや体積に大きなばらつきがあることが分かります。
このとき使用される方法はアロメトリー(allometry)と呼ばれます。
アロメトリーでは、ある種の脳重量を、その種が最も近縁であると考えられる祖先種に似た現存種の脳重量で割ることで得られる進化的傾向(evolutionary trend)を示します。
この比率(progression index, PI)は、種間比較における特定の脳領域の機能的重要性を示す指標となります。
最も原始的な霊長類は昆虫食性(insectivores)に由来しているため、現存する種を比較する際には、ツパイ(tree shrews)、原猿類(prosimians、例:ブッシュベイビーやキツネザル)および類人猿(monkeys and apes)を含めたサンプルを使うのが適切です。
霊長類では、体重に対する脳の重さが、非霊長類の哺乳類と比べて約2~3倍大きいことが示されています(ただし歯のないクジラ類は例外)。
この傾向は、特に果実食(frugivory)を行う種で著しく、果実食に伴う季節的・空間的な資源の分散に対応するため、より多くの計算資源(computational resources)が必要だったと考えられます。
注目すべきことに、脳の重さと脳のサイズの間には強い相関があることが明らかになっています。
すなわち、脳サイズは基本的に体重に比例して増加します。
これは、脳が体の他の部分と同様にエネルギー消費の観点から設計されていることを示唆しています。
ただし、ヒトは比較的小さな体重にもかかわらず、安静時エネルギー消費量が約15Wと低いにもかかわらず、脳が消費するエネルギーは総エネルギーの15~20%と高率です。
これは、ヒトの脳が体重のわずか2%しか占めないにもかかわらず、非常にエネルギー集約的な器官であることを示しています。
特定の脳領域のサイズを、脳全体のサイズではなく、体重に関連づけて比較すると、ヒト進化の過程でどの領域が相対的に拡大し、どの領域が縮小したかの印象を得ることができます。
たとえば、嗅球(olfactory bulbs)の重要性は、霊長類の進化過程で一貫して低下してきました。
ツパイでは脳重量の7%、ガラゴでは2%、サルでは0.2%、ヒトでは0.01%しか嗅球が占めていません。
PI(progression index)で見ると、ヒトの嗅球は、体重補正後の基準値の1/140~1/150にしか達しないのに対して、ネオコルテックス(neocortex)は非常に拡大しており、ヒトではPIが58にもなっています。
これらの観察結果から、ヒトとチンパンジーの祖先が約600万年前に分岐した後、脳の構造的拡張が加速されたことが示唆されます。
この過程で、ヒトのネオコルテックスは、体重に対して期待される大きさの3倍にまで拡大しました。
そのため、体サイズにおける差異(現在のヒトと類人猿の間に見られる)を考慮しても、ヒト脳の相対的な大きさはチンパンジーの脳よりもはるかに大きいと結論できます。
また、もしヒトの体重が現代よりも増減したとしても、脳サイズと体重の関係は変わらなかったであろうと推測されます。
ヒトは通常、体重約450kgで身長3mに達するか、あるいは現状とほぼ同じ大きさだったでしょう。
注釈
(訳)
哺乳類では、脳重量は体重に対して増加します。これは、基礎代謝エネルギー消費と同様に、体重の3/4乗に比例しています。
(訳)
アロメトリー曲線を用いることで、体重の違いを補正しながら、近縁種および遠縁種の脳の比較が可能になります。
(訳)
ヒトの脳は、体重に対して期待される大きさの平均で約3倍ですが、嗅球の大きさは原始的霊長類の期待値の約1/150しかありません。
ヒトの新皮質は他の霊長類のそれよりも三倍以上大きい(ヒトの体サイズに対する脳サイズの比率は、霊長類のなかでも特に大きい)。また、ヒトの脳は出生後も高い割合で成長を続けるが、これはヒトの出産時の未熟性と関係している。この特徴はヒトの骨盤構造に由来する設計上の妥協を反映している(詳細は第3章、図2.3を参照)。
アロメトリーの話題に戻る前に、ヒトの脳において進化的により古い部分――特に扁桃体、海馬、帯状皮質――も不均衡に大きく成長している点を指摘しておくことが重要である。例えば海馬では、CA1領域(特に新皮質と密接に結びついていない部分)がヒトの海馬体積全体の44%を占めている。この領域の成長指数(PI)はヒトでは6.6であり、線条体においても同様に進化的なサイズ増加が見られる。線条体のPIは、ヒトでは16.4、キツネザル類では9.0、プロミシアン類では4.7である。これらの変化は、感情処理、恐怖や攻撃の制御、記憶形成に関わる辺縁系構造の機能的重要性の増大を反映している。なお、扁桃体の領域も一様ではなく、たとえば、ある領域は食物探索や性行動への影響に関与する一方で、別の領域は防衛行動や攻撃行動に関連している。
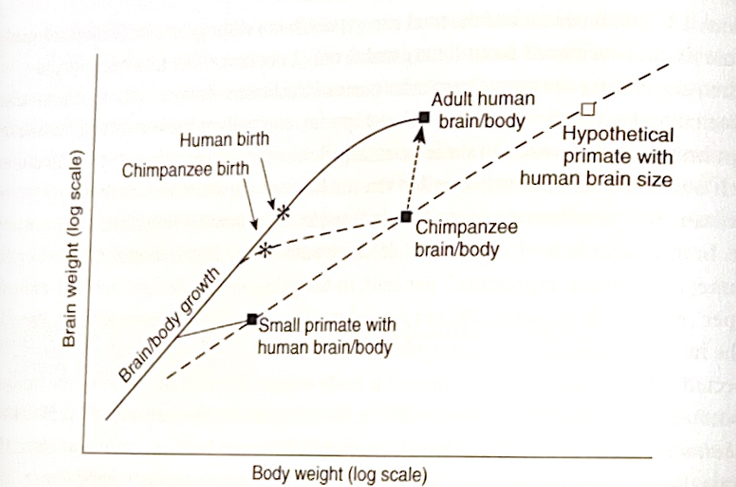
図2.3の解説(日本語版)
図2.3は、「ヒトの脳と身体の成長パターン」が、チンパンジーなど他の霊長類とどう違うかを示したものです。
- 通常、脳と身体はバランスよく成長します。
- しかしヒトでは、出生時にすでに脳が非常に大きく、しかも出生後も急速に脳が成長します。
- これは、ヒトが「未熟な状態で」生まれてくるからです(胎児の段階で脳を十分に成長させることができず、出生後に成長を続ける必要がある)。
- 仮にヒトがチンパンジーなどと同じような脳-身体成長比率を保ったまま脳をここまで大きくしたなら、体もそれに合わせて巨大化していたはずです。
- 具体的には、体重約450kg、身長約3mという、「キングコングモデル」と呼ばれる想定になります。
つまり、ヒトは脳を大きくするために、未熟なまま生まれて、出生後に脳を急成長させるという「特殊な進化戦略」を取ったわけです。
図の線の意味を簡単に言うと:
- 実線(人間の成長パターン)→ 脳がどんどん大きくなる。
- 破線(他の霊長類や仮想の霊長類)→ 脳と身体がもっとバランスよく成長。
また、ヒトでは自伝的記憶のための海馬形成体が拡大しており、これは社会的相互作用において重要な意味をもつ。このなかで特に注目すべきは、帯状回であり、これは辺縁系に分類されるが、進化的変化を遂げてきた。より前方の帯状皮質は、より複雑な意思決定や注目の分配に関わる機能的分化を示している。
線条体の拡大は、モチベーションや報酬予測、予期行動に関わる機能の進化的増大を示している可能性がある。しかし小脳については、ヒトでは比較的小さな増大にとどまっており、これは新皮質の急速な進化についていくことができなかった可能性を示唆している。特に小脳の新皮質領域(小脳新皮質)は、視空間的スキル、人間特有の道具使用や複雑な動作系列の計画、運動学習、注意配分に関与する領域であり、ヒトの進化に伴う高度な行動様式の発展にも関わっている可能性がある。
最初に目立つ発見は、ヒトの前頭葉――特に新皮質に対する相対的サイズ――は、体サイズに対して予想されるほどには大きくないという事実である。ただし、ヒトの前頭前野皮質は、たとえば後方の頭頂葉や側頭葉、運動領野や一次体性感覚領野に比べると、より高度に折りたたまれ(高い脳回形成)ている。この違いは、ヒトの前頭葉と頭頂葉の間の発達タイミングの違いとも関係しており、頭頂葉の一部(たとえばブロードマン領域BA13)はヒトにおいても霊長類においても、比較的後期に発達する。
機能的には、前頭葉の領域は未来の行動計画や意図の表現、社会的刺激に対する情動的意義の評価に関与している。前頭前野の内側部の損傷は動機づけの欠如を引き起こし、背外側前頭前野の損傷は思考と動作の分離をもたらす。特に、内側前頭前野皮質は、自己と他者の意図や感情の表象に関わっているとされ、ヒトの社会的適応の中核をなしている。
また、側坐核も注目すべき領域であり、ヒト個体の50%未満でしか発見されないが、生涯を通じて適応的変容に関与していると考えられている。
注釈(本文中の斜体部分)
- ヒトの新皮質は、他の霊長類に比べて三倍以上大きい。ヒトの脳の高い成長率は、出産時の未熟性に関係しており、これが出生後の脳の急速な成長を可能にしている。
- 扁桃体の領域には、食物探索や性行動に関与するものと、防衛行動や攻撃行動に関与するものとがある。
- ヒトでは線条体と小脳が、他の霊長類に比べて相対的により大きなサイズ増加を示している。特に線条体の成長は、より高度な道具使用や複雑な行動の発展と関係している可能性がある。
- ヒトの前頭葉の一部(特に側坐核)は、個体の約半数でしか発見されず、継続的な適応的変容に関与している可能性がある。
側頭葉は、他の領域と比較して人類系統において不均衡に増大しており、とりわけ白質の量において顕著である。これは、その機能的特殊性、すなわち音声認識、言語生成、顔認識および意図の認識に関連する可能性がある。側頭極は自伝的記憶の貯蔵に寄与している。側頭葉下部の紡錘状回は、個人が顔を識別するために顔の不変的な側面を認識するために重要であり、顔がどの角度から見られても認識可能にする。
何らかの理由で、頭頂葉の進化史は依然としてある程度不明瞭である。しかし、下頭頂小葉は、おそらく進化的再編成を遂げている。下頭頂小葉を構成する縁上回と角回は、前頭葉および側頭葉皮質との重要な連絡を持ち、統合感覚、自己認識、自己と他者の区別などに重要な役割を果たしている。この領域への損傷は、半側空間無視(特に右頭頂葉の損傷後)、洞察力および自己反省の欠如、行動および信念の誤帰属(いわゆる受動症状)を引き起こす可能性がある。新皮質のより内側の領域に対して、一次視覚野は後方へ移動し、類人猿に比べて面積が相対的に小さくなっているが、これは長距離の結合繊維の増加によるものであると考えられる。
特筆すべきは、霊長類における相対的な脳の拡大が、種の個体寿命の延長とも関連している点である。これは、食糧不足や予測不能な環境変動といった出来事をより多く経験するため、長寿命の種ほどこれらの困難に対処するための認知的資源を多く必要とすることによって説明できる。脳の重量から推定される最大ヒト寿命は約101.5年であり、これは実際の最大寿命に非常に近い。人類では、相対的な長寿は、新皮質、扁桃体、視床下部、脳幹など脳のさまざまな部位の類似した方法による発達と相関していることがわかっている。
ポイント
側頭葉は、音声の理解と生成、顔の認識および意図的な動きの認知において機能的に重要である。側頭紡錘状回は、異なる視角においても不変な顔の特徴の認識に寄与している。
