
- 第6章 自閉症とその他の広汎性発達障害
- 🔍 概要:自閉症と「心の理論」の障害
- 🧠 ASDにおける「心の理論」障害の特徴
- 🚫 他の認知機能障害とは異なる
- 👥 社会的情報処理の障害の具体例
- ⚖️ 社会的規範(公平性・互恵性)への理解の困難
- ✅ まとめ
- 🔹 自閉症の「メンタライジングの障害」は知能とは別の問題
- 🔹 知能が高くても「社会的認知の発達の遅れ」が顕著
第6章 自閉症とその他の広汎性発達障害
自閉症の基礎知識
原因と特徴
自閉症の特定の環境リスク要因は現在のところ不明です。
自閉症は、コミュニケーション障害、社会的交流の障害、そして限定的・常同的な行動という3つの症状を特徴としています。この障害は幼児期早期に発症します。
自閉症の症状の重症度はかなり異なります。アスペルガー症候群では言語によるコミュニケーションは保たれていますが、カナー型自閉症では知的障害やてんかんを伴うことがあります。知的能力が保たれている自閉症の人々は、しばしば優れた技術的知能や特異な才能を持っていることがありますが、これは社会的領域における困難さと対照的です。
疫学
一般人口における自閉症の有病率は1,000人あたり3〜6人です。男女比は約3〜4:1で、男性に多く見られます。
遺伝的要因
自閉症は高い遺伝性を示します。一卵性双生児の一致率は40〜90パーセントです。遺伝形式は多遺伝子性ですが、個々の遺伝子の正確な寄与度は不明です。ゲノムインプリンティングなどのエピジェネティック因子も関与している可能性があります。
行動観察と愛着
自閉症児の行動観察から、養育者に近づくことと回避する動機の間の葛藤が示唆されています。密接な接触の回避により、子どもと養育者の両方が情緒的な絆を確立することが困難になります。その結果、自閉症児では健常対照と比較して不安定な愛着スタイルがより多く見られます。
脳機能
自閉症は、他者の心的状態を理解する能力を含む、社会的情報処理の選択的障害と関連しています。模倣や視線の追跡、共同注意など、心の理論の発達に先立つ他の発達段階も自閉症では機能不全を示します。これはミラーニューロンシステムの障害と関連している可能性があります。
自閉症の脳が「男性脳」の極端な変異である可能性があるという観察から、胎児期のテストステロン曝露が自閉症の病因に関与しているという仮説が生まれました。
ゲノムインプリンティングに基づく別の仮説では、父親または母親からインプリントされた遺伝子が自閉症の表現型特性に寄与していると示唆されています。
鑑別診断と合併症
自閉症の最も重要な鑑別診断は、小児期統合失調症と知的障害です。注意欠如・多動性障害(ADHD)、ジル・ド・ラ・トゥレット症候群(GTS)、緊張病などとの併存も見られます。
経過と治療
自閉症スペクトラム障害は慢性的であり、自然寛解は知られていません。年齢と発達が進むにつれて、自閉症に見られる社会的な困難さには変化が現れることがあります。高機能自閉症であっても、皮肉やフォーパス(社会的失敗)状況などの微妙な心的状態の理解には困難が残ります。
自閉症には現在のところ根本的な治療法はありません。社会的・コミュニケーション能力のトレーニングや職業療法が重要な非薬物的介入戦略です。薬物治療は一般的に推奨されておらず、深刻な行動上の問題がある場合に限って用いられます。
1. 症状と診断基準
自閉症および自閉症スペクトラム障害は臨床的に、コミュニケーション障害、社会的相互作用の障害、そして限定的・反復的な行動や興味という3つの症状(三徴候)を特徴とし、乳幼児期早期に発症します。社会的・コミュニケーション上の障害には、顔の探索や他のコミュニケーション信号の理解の欠如、視線回避、他者の視線方向の追跡や共同注意の形成ができないことなどが含まれます。また、信念、欲求、知識、意図、気質といった他者の心的状態を理解する能力の著しい欠如や発達の遅れも見られます。
【ポイント】 社会的信号の認識・処理や模倣における障害は非常に顕著です。
自閉症の人々はまた、顕著な共感能力の欠如を示し、しばしば他者の存在を無視します。さらに、自閉症の子どもたちは観察した行動を模倣(エミュレート)したり、象徴的な遊びをしたりする能力に障害があります。その一方で、彼らはしばしば常同行動に従事し、時には他者の無意味な行動の模倣(エコプラクシア)や言葉の模倣(エコラリア)の形をとることがあります。エコラリアの場合、イントネーションさえも保持されることがあります。
自閉症スペクトラム障害の重症度は、言語コミュニケーションが保たれているアスペルガー症候群や高機能自閉症(その境界はあいまいに定義されています)のような軽度の形態から、知的障害(カナー型自閉症の約3分の2のケース)やてんかん(3分の1)を伴う重度の形態(カナー型自閉症)まで様々です。自閉症者の約20%は知的機能が正常です。
社会的認知の障害の重症度とは対照的に、知的機能が正常な自閉症の人々は優れた技術的知能や視覚的識別能力を持っていることがあり、技術系の職業に就いている親族が対照群の家族よりも自閉症の人々の家族に多く見られることがあります。さらに、自閉症の人々の中には、カレンダー計算、音楽、絵画など様々な分野で並外れた才能を持つ人々がいます。このいわゆるサヴァン(特異的才能者)はしばしば知的に重度の障害を持っています。
上記の症状三徴候は自閉症に典型的と考えられていますが、集団ベースの研究では、これら3つの中核領域の相関は中程度にすぎず、これらの領域のいずれかにおける単独の困難さが子どもに生じることがあります。そのような場合、自閉症の診断基準を満たさないものの、より広い自閉症の表現型スペクトルに分類されます。
行動の異常は生後間もなく認識できることがありますが、通常、診断は生後15〜24ヶ月の年齢でなされます。
2. 疫学
狭義の自閉症は、10,000人あたり平均5.2人に影響する稀な障害です。過去25年間で症例定義と診断感度の変化により、10,000人あたり7.2人へと相対的に増加しています。軽度の症例を含む自閉症スペクトラム障害の推定有病率は1,000人あたり3〜6人で、すべての広汎性発達障害(PDD)は10,000人あたり約19人の範囲です。
表6.1 自閉性障害のDSM-IV-TR診断基準
A. 以下の(1)、(2)、(3)から合計6項目(またはそれ以上)を満たし、そのうち(1)から少なくとも2項目、(2)と(3)からそれぞれ1項目ずつ満たす: (1) 社会的相互作用における質的障害(以下のうち少なくとも2つ): (a) 目と目を合わせる視線、表情、姿勢、身振りなど、複数の非言語的行動の使用における著しい障害 (b) 発達水準に適した仲間関係の形成の失敗 (c) 楽しみ、興味、達成感を他者と自発的に共有しようとすることの欠如(例:見せる、持ってくる、興味のある物を指さすことの欠如) (d) 社会的または感情的な相互性の欠如
(2) コミュニケーションにおける質的障害(以下のうち少なくとも1つ): (a) 話し言葉の発達の遅れまたは完全な欠如(身振りやマイムなどの代替コミュニケーション手段で補おうとする試みを伴わない) (b) 適切な発話を持つ個人における、他者との会話を始めたり続けたりする能力の著しい障害 (c) 常同的で反復的な言語使用または特異的な言語 (d) 発達水準に適した多様で自発的なごっこ遊びや社会的模倣遊びの欠如
(3) 行動、興味、活動の限定的・反復的・常同的パターン(以下のうち少なくとも1つ): (a) 強度や焦点において異常な、1つ以上の常同的で限定された興味パターンへの没頭 (b) 特定の非機能的な習慣や儀式への明らかに柔軟性のない固執 (c) 常同的で反復的な運動常同症(例:手や指のひらひら運動やねじり、または複雑な全身運動) (d) 物体の一部への持続的なこだわり
B. 3歳未満の発症を伴う、以下の領域のうち少なくとも1つにおける遅れまたは異常な機能:(1) 社会的相互作用、(2) 社会的コミュニケーションに使用される言語、または (3) 象徴的または想像的遊び。
C. この障害はレット障害または小児期崩壊性障害ではうまく説明できない。
表6.2 アスペルガー障害のDSM-IV-TR診断基準
A. 社会的相互作用における質的障害(以下のうち少なくとも2つ): (1) 目と目を合わせる視線、表情、姿勢、身振りなど、複数の非言語的行動の使用における著しい障害 (2) 発達水準に適した仲間関係の形成の失敗 (3) 楽しみ、興味、達成感を他者と自発的に共有しようとすることの欠如(例:見せる、持ってくる、他者に興味のある物を指さすことの欠如) (4) 社会的または感情的な相互性の欠如
B. 行動、興味、活動の限定的・反復的・常同的パターン(以下のうち少なくとも1つ): (1) 強度や焦点において異常な、1つ以上の常同的で限定された興味パターンへの没頭 (2) 特定の非機能的な習慣や儀式への明らかに柔軟性のない固執 (3) 常同的で反復的な運動常同症(例:手や指のひらひら運動やねじり、または複雑な全身運動) (4) 物体の一部への持続的なこだわり
C. この障害は社会的、職業的、またはその他の重要な機能領域において臨床的に著しい障害を引き起こす。
D. 言語の臨床的に著しい全般的な遅れはない(例:2歳までに単語を使用し、3歳までにコミュニケーション的な句を使用)。
E. 認知発達、年齢相応の自助スキル、適応行動(社会的相互作用以外)、および幼少期の環境に対する好奇心の発達において臨床的に著しい遅れはない。
F. 他の特定の広汎性発達障害または統合失調症の基準を満たさない。
世界的な有病率はある程度異なりますが、様々な文化的背景から症例が報告されています。男女比は約3〜4:1です。
3. 遺伝的リスク要因
自閉症は高い遺伝性を持ち、自閉症の子どものきょうだいが障害を発症するリスクは、一般集団と比較して30〜120倍高くなっています。一卵性双生児(MZ)の一致率は40〜90パーセント(狭義の定義基準を適用すると約60パーセント)であり、二卵性双生児(DZ)では0〜10パーセントです。
自閉症は単一の疾患単位ではなく、少なくとも10〜20の異なる対立遺伝子が関与する症候群です。染色体1q、2q、3p、4、7q、15q、17q、および性染色体上に位置する遺伝子が自閉症スペクトラム障害に役割を果たしているという証拠があります。これらの遺伝子座の機能的意義は部分的にしか分かっていません。特に興味深いのは、推定される言語と発話の領域(FOXP2)の近くに位置する染色体7qの対立遺伝子、染色体3p上のオキシトシン受容体の対立遺伝子変異、染色体15qの重複を含む細胞遺伝学的異常、および染色体17q上のセロトニントランスポーター遺伝子変異です。最近、染色体1q上にあり神経発生に関与していると考えられる遺伝子DISC1(「統合失調症において破壊された」)と自閉症およびアスペルガー症候群との関連が明らかになりました。
ゲノムインプリンティングなどのエピジェネティック要因が自閉症の病因に関与しているかどうかはまだ不明ですが、理論モデルでは、父性または母性インプリンティングがそれぞれ脳における男性的特性の過剰発現や社会的認知障害につながる可能性があることが示唆されています(以下参照)。しかし、双生児研究により、症状三徴候の三領域は高い遺伝性を持つものの、遺伝的に部分的にしか重複していないことが示唆されているため、生物学的マーカーを使用してエンドフェノタイプ(中間表現型)を特徴づけることが、異なる症状側面に対する遺伝子の寄与を明らかにするのに役立つ可能性があることを強調すべきでしょう。
4. 環境的リスク要因
自閉症の特定の環境的リスク要因は不明です。結節性硬化症、脆弱X症候群、出生前の風疹やサイトメガロウイルス感染、または出生前の毒物や催奇形性物質への曝露を持つ個人では、自閉症を発症する相対的リスクが高まります。しかし、これらの要因は症例の10〜15パーセント未満にしか当てはまりません。親の年齢が高いことが自閉症を含む広汎性発達障害(PDD)の有病率の増加と関連している可能性があるという証拠があります。
5. 病態生理学的メカニズム
自閉症における病態生理学的メカニズムは部分的にしか理解されていません。自閉症の人々ではエンドルフィンや脳由来神経栄養因子(BDNF)のレベルが上昇しているという証拠があります。一方、セロトニンに関する相反する知見では、血中レベルの上昇が見られますが、中枢神経系での利用可能性は低下しています。社会的認知、絆形成、性行動の重要な媒介物質であるオキシトシンは、自閉症では発現が低下しています。アポトーシス(プログラムされた選択的細胞死)は明らかに減少しており、これが自閉症の脳の一部における小さく密に詰まったニューロンの数の増加の原因となっている可能性がありますが、これらのニューロンは脳の他の部分との連結が不十分です。
胎児のテストステロン濃度の上昇や発達中の脳におけるアンドロゲン受容体数の増加によって、神経伝達物質レベルでのいくつかの変化が媒介されるという推測がありますが、現時点ではこれは実証されていません。同様に、自閉症の子どもを持つ母親のセロトニンレベルの低下が、脳の発達と成熟の変化に関連していることが報告されています。しかし、自閉症障害の異質性を考慮すると、全体的な知見は決定的ではありません。
模倣や他者の心的状態を推測することの困難さから、模倣学習にとって重要であり、おそらく他者の心的状態のシミュレーションに寄与すると考えられているミラーニューロンシステムが、自閉症の人々では機能的に障害されているという仮説が立てられています(後述)。
6. 進化論的統合
自閉症スペクトラム障害は非常に多様な性質を持ちますが、行動、認知、感情のレベルで社会的脳機能に関連する心理的メカニズムに選択的に影響を及ぼすようです。自閉症の子どもの行動学的観察から、少なくともこの障害のいくつかの軽度の形態では、おそらく高まった臆病さと恐怖のために、養育者に近づくことと回避することの間の動機づけの葛藤があることが示唆されています。
近接接触の回避は非常に早い段階から明らかであり、子どもと養育者の両方が情緒的絆を確立することを困難にします。それに応じて、健常対照群と比較して、自閉症の子どもでは不安定な愛着スタイルがより一般的であることが見出されています。近因レベルでは、これは自閉症の子どものオキシトシンとセロトニンの代謝回転の変化に関連している可能性があります。子どもの臆病さに対する養育者の反応は、乳児を社会的相互作用に強制しないことによって絆の形成を促進するか、あるいは視線を合わせようとする繰り返しの試みや物理的な近接を強引に確立しようとすることによって状況を悪化させる可能性があります。しかし、自閉症の子どもの親は平均して、正常に発達している子どもの親よりも敏感さが低いわけではありません。したがって、安定した母子二者関係を形成する問題は、自閉症の子どもの側でより顕著である可能性があり、重度の自閉症の場合、子どもと養育者の間の情緒的絆はおそらく決して確立されないかもしれません。
高機能自閉症とアスペルガー症候群を含む学齢期の自閉症の子どもたちは、同年齢の子どもたちとの社会的遊びに参加することが少なく、社会的孤立をより頻繁に経験します。成人として、高機能自閉症やアスペルガー症候群の人々を含む自閉症の人々は、親密な社会的関係や親密さを確立することに依然として深刻な困難を抱えています。
過去20年間、多くの研究により、自閉症の人々は他者の心的状態について推論することによって、思考、意図、感情、欲求、気質を理解することに深刻な問題を抱えていることが一貫して示されてきました。この「心の理論」または「メンタライジング」の障害は、他の認知機能障害の直接的な結果ではなく(実行機能の計画障害や自伝的出来事の再構成の乏しさを伴いますが)、メンタライジング、感情認識、顔の処理を含む社会的情報処理の選択的な障害を反映しています。これに沿って、自閉症の人々は公平性や互恵性などの社会的規則や規範を理解することに困難を示すことが明らかになっています。
しかし、自閉症におけるメンタライジングの障害は知能と直接結びついているわけではありません。むしろ、平均以上の知能を持つ自閉症の人々では、社会的認知の発達の遅れは顕著です。年齢が上がるにつれて他者の心に関する基本的な理解が発達する可能性がある自閉症の軽度の形態であっても、自閉症は単に成長するとともに解消される障害ではありません。
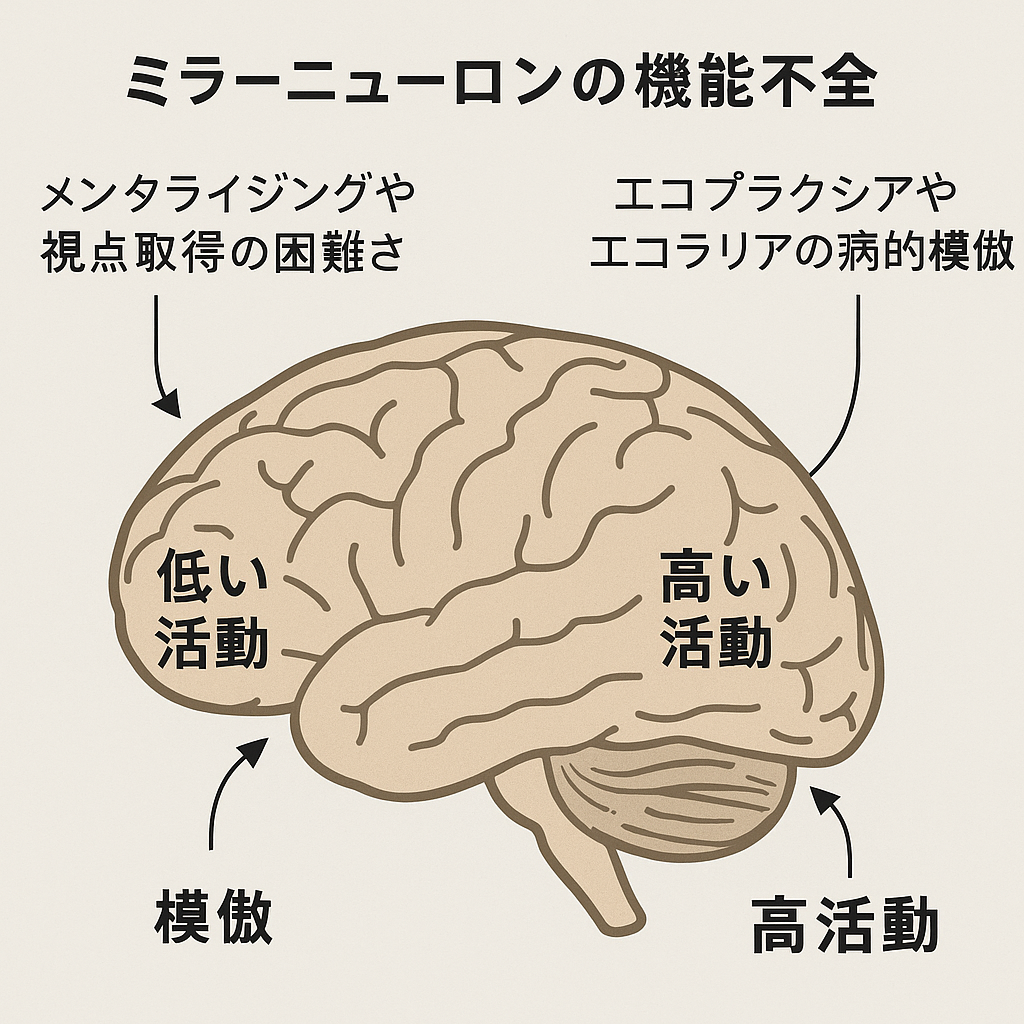
メンタライジングに関わる社会的認知機能は、子どもたちにおいて明確な発達段階で現れます(第2章の後記と比較してください)。しかし、メンタライジングの障害だけでは自閉症における社会的孤立を説明するには不十分です。なぜなら、自閉症における行動異常の発症は、通常発達している子どもがメンタライジング能力を獲得する年齢よりも明らかに早いからです。ミラーニューロンシステムの機能不全は、初期の行動異常と自閉症における欠陥のあるメンタライジングシステムを結びつける可能性があります。
種間比較やヒト以外の霊長類の研究から豊富な証拠があり、模倣による学習はミラーニューロンと呼ばれる特定の神経細胞集団の活動と関連していることが示唆されています。これらのニューロンは、手の動きの観察と同じ動きの実行の両方で顕著に発火することからその名が付けられています。ミラーニューロンは、動きの結果が観察から隠されている場合でも活性化します。したがって、それらは複雑な視覚的認知と予測の認知プロセスを結びつけます。
行動の模倣と観察された行動とその行動の実行との間の関連を確立することは、初期の学習経験にとって重要であるだけでなく、模倣能力はメンタライジング能力の前駆体でもあります。行動を模倣し、心的状態をシミュレーションすることは、想像力豊かに他の個人の視点を取ることができるという点で共通しています。しかし、メンタライジングには、自己と他者の区別や現実と見かけの区別など、追加のメカニズムが必要です。いずれにせよ、視線の追跡や共同注意などのメンタライジングに先行する模倣やその他の発達段階はすでに自閉症では発達不全です。ミラーニューロンシステムの機能不全は、神経活動の抑制によるメンタライジングや視点取得の困難さと、神経活動の脱抑制によるエコプラクシアやエコラリアの形での病的模倣の存在の両方を説明する可能性があります。
自閉症におけるミラーニューロンシステムの欠陥の仮説は、自閉症では皮質の厚さがミラーニューロンを含むと推定される前頭前皮質と側頭領域で減少しているという観察によってさらに支持されており、これは自閉症における他の部位での皮質厚の増加とは対照的です。メンタライジングに一貫して関与している他の脳領域は、機能的脳イメージング中に活動が低下しており、そのような異常な活性化パターンには前帯状皮質(ACC)と傍帯状皮質が含まれています。これらの領域はセロトニン神経が密に分布しており、前頭前皮質におけるセロトニン機能不全があると、おそらく自閉症では機能的に抑制されています。
共感的な視点取得やメンタライジング、社会的感受性や言語流暢性は、おそらく女性が男性よりも発達しており、これは女性の親の投資量がより大きいことによるものかもしれません。対照的に、心的回転、空間定位、物理的問題解決は平均して男性の方が優れています。自閉症における顕著な(そして選択的な)社会的認知の障害と、しばしば保存されているか、あるいは優れている技術的理解との対比から、自閉症の脳は「男性脳」特性の極端な変異である可能性があるという仮説が生まれました。これは過剰な胎児テストステロンの影響下で発達し、おそらく父性インプリント遺伝子の結果として生じる可能性があります。
この仮説を支持する間接的な証拠として、先天性副腎過形成の女児は、影響を受けていない姉妹と比較して、より男の子らしい行動や自閉症のような特性を示すという観察があります。胎児テストステロンは第2指と第4指の長さの比率と逆相関しており、このような低い比率が自閉症で見られます。さらに、自閉症の男児では思春期の開始が早熟であるという証拠もあります。
正常に発達している子どもでは、胎児テストステロンのレベルは、目と目を合わせる接触、共同注意、言語発達、社会的機能、そして極端な形で自閉症の症状に似るような限定された興味の範囲の量と逆相関しています。自閉症が胎児テストステロンレベルの上昇や性的二形性脳領域におけるアンドロゲン受容体数の増加と関連しているかどうかは不明です。
いずれにせよ、解剖学的データは、自閉症が脳の肥大、出生時の扁桃体のサイズの増大(思春期後に相対的なサイズが正常以下の体積に低下)、神経細胞密度の増加(ただし連結が不十分)、そして生後数年間の白質量の増加と関連していることを示唆しています。逆に、自閉症における半球内および半球間の連結に関するデータは、半球間の連結の減少と半球内の連結の減少を示す知見と矛盾しています。しかし、半球間の連結の減少は、「中心的統合性」の欠如、つまり部分よりも全体(ゲシュタルト)を認識する能力のような認知プロセスの機能不全を説明できるかもしれません。
この知見と一致して、自閉症のサヴァンは世界を文字通りに見ていると提案されています。なぜなら彼らは概念や心的テンプレートの正常な獲得を欠いており、したがってゲシュタルトではなく詳細の知覚に依存しているからです。これはまた、自閉症の人の話し言葉の解釈における文字通りさを部分的に説明することもできます。このような違いは、正常な男性と女性の間にも存在します(ただし、はるかに程度が低い)。したがって、男性的特性と女性的特性の差異的な遺伝や発現が自閉症の病因に関与していると考えられます。
進化的包括適応度理論は、母方と父方から受け継がれた遺伝子の間でゲノム内の葛藤が生じる可能性があることを示唆しています。父親からの遺伝子は、母方から受け継がれた遺伝子よりも母親からより多くの資源を引き出すように設計されています。ゲノムインプリンティングの場合のように、もう一方の親からの遺伝子が沈黙させられると、父方または母方由来の遺伝子が子孫で過剰に発現される可能性があります。
この段階では、ゲノムインプリンティングが自閉症において主要な役割を果たしているというのは推測的です。しかし、行動的にも解剖学的にも男性的特性の過剰発現、胎盤のサイズの増大(増殖性成長の兆候を伴う)、および自閉症の子どもの出生時体重の増加は、父性インプリンティングが少なくとも自閉症スペクトラム障害の一部に関与しているという仮定を強く支持しています。
さらに、染色体15上の父性遺伝子の過剰発現によって引き起こされる疾患であるアンジェルマン症候群(第1章参照)は、言語獲得障害、視線回避、常同行動など自閉症症状の高い有病率を特徴としており、これは父性遺伝に向けた遺伝子発現の不均衡のさらなる証拠と解釈できるかもしれません。
自閉症との関連については、これらの子どもたちは、剥奪の期間と発達年齢によって、豊かな環境でしばしば急速に改善します。
自閉症におけるゲノムインプリンティングに関するやや異なる仮説は、X染色体の部分的または完全な欠失を特徴とするターナー症候群の研究から生まれています。X染色体を1つだけ持つ女性は、父親または母親からX染色体を受け継ぐかによって、社会的認知能力と実行機能に違いがあります。X染色体が母親由来である場合(70%のケース)、社会的認知障害や自閉症様の特徴は、X染色体が父親由来であるターナー症候群の個人と比較してより一般的です。
これは、社会的認知に関与するX染色体上に遺伝子座が存在する可能性を示唆しており、正常な女性(46、XX)では父親のX染色体からのみ発現し、母親から受け継がれた遺伝子座は沈黙しています。これは、なぜ女性が一般的に男性と比較して社会的認知が優れているか(男性は常に母親からX染色体を受け継ぐため、母性X染色体の不活性化が起こらない)だけでなく、なぜ男性が自閉症スペクトラム障害を含む発達障害に対してはるかに脆弱であるか、そしてなぜターナー症候群(45、X0)の個人が父親のX染色体を持つX0女性と比較して母親由来のX染色体を持つ場合により多くの自閉症症状を示すかについても説明できます。
両方のインプリンティング仮説は部分的な実証的支持を受けていますが、おそらくどちらも自閉症スペクトラムのすべての特徴を説明するには不十分です。母親または父親の染色体から発現されるインプリント遺伝子は、実際の表現型を生成するためにゲノムの他の場所にある感受性遺伝子座と相互作用する可能性が高いです。したがって、両方のインプリンティングシナリオが自閉症スペクトラムの異なるサブタイプに適用される可能性があると考えられ、この推測は将来の研究の対象となるかもしれません。
7. 鑑別診断と併存症
自閉症の最も重要な鑑別診断は、小児期統合失調症と知的障害です。小児期統合失調症は12歳未満ではまれであり、通常、妄想と幻覚を伴います。行動症状を伴う知的障害は、対象者があまり社会的に引きこもらず、通常は家族や仲間と交流するという点で自閉症とは異なります。心理社会的剥奪のケースは、注意欠陥・多動性障害(ADHD)、ジル・ド・ラ・トゥレット症候群(GTS)、および緊張病と混同される可能性があります。
自閉症の主な併存障害は、注意欠陥・多動性障害(ADHD)、ジル・ド・ラ・トゥレット症候群(GTS)、そして時々緊張病です。併存症に関しては、精神医学的障害は離散的な疾患単位というよりも特性の次元的表現であることを念頭に置くべきです。したがって、他の障害との症状の重複が頻繁に観察されることがあります。
8. 経過と予後
自閉症スペクトラム障害は、自然に寛解することが知られていない慢性的な障害です。しかし、年齢と発達が進むにつれて、自閉症に見られる社会的な困難さには変化が現れることがあります。
乳児期には、自閉症のある個人はしばしば社会的な無関心さを示し、時には他者を完全に無視することもあります。この行動パターンは、子どもから働きかけるイニシアチブがほとんどない、受け身の社会的交流スタイルへと発展することがあります。
思春期から成人期にかけて、自閉症のある人々はより積極的に社会的交流に関わるようになることがありますが、その様子は奇妙であったり独特であったりします。特に軽度の自閉症のある年長の人々は、他者の心を基本的に理解できる発達段階に達することがあります。しかし、皮肉や「ある特定の人の前で言うべきではないことを言ってしまう」といったフォーパス状況のような、より微妙な心の状態を理解することには依然として困難を抱えています。
高機能自閉症やアスペルガー症候群の人々でさえ、通常は自己中心的な視点を持ち続け、場合によっては被害妄想的な思考が生じることもあります。IQが70以上で言語能力がある場合は、比較的良好な予後と関連しています。しかし、自閉症のある人々のうち、独立した、あるいは半独立した生活を送ることができる発達段階に達する人はごく一部です。
9. 治療
自閉症障害や広汎性発達障害(PDD)に対する原因療法はありません。心理教育的介入には、社会的交流を促進することや、子どもの個々のニーズに合わせた行動介入が含まれます。
さらに、社会的・コミュニケーション能力のトレーニングや職業療法も重要な非薬物的介入戦略です。自閉症の人々を無理に社会的交流に強制することは有用ではないことが証明されています。なぜなら、これはストレス反応や社会的引きこもりをかえって増加させるからです。したがって、過剰な刺激を避けることは、特に持続的な注意の問題を考慮すると、自閉症の子どもたちに適切かもしれません。
また、自閉症の人々が他者の心を理解することの困難さを考慮すると、二重の意味を持つ会話を避けるのが有益かもしれません。さらに、他の人々は自分とは異なる期待、知識、意図を持っていることを慎重に教育することも役立つでしょう。
家族への介入とサポートも、評価と治療手順の不可欠な部分であるべきです。自閉症の心理療法は、高機能自閉症やアスペルガー症候群における抑うつや不安などの併存障害を除いて、成功していません。
薬物治療は一般的に推奨されておらず、深刻な行動上の問題に限られています。セロトニン再取り込み阻害薬(SSRI)が常同行動や自傷行為を減らすのに効果的である可能性を示す証拠があります。過度の攻撃性や衝動性は、第二世代の抗精神病薬やナルトレキソンに反応することもあります。オキシトシンの実験的投与により、自閉症やアスペルガー症候群の患者の音声からの感情認識(韻律)が改善されることが明らかになっています。
Selected further reading
Asperger, H. 1944, ‘Die autistischen Psychopathen im Kindesalter’, Archiv für Psychiatrie und
Nervenkrankheiten, vol. 117, pp. 76-136.
Badcock, C. & Crespi, B. 2006, ‘Imbalanced genomic imprinting in brain development: an evolutionary
basis for the aetiology of autism’, Journal of Evolutionary Biology, vol. 19, pp. 1007-1032.
Baron-Cohen, S. 1991, ‘The theory of mind deficit in autism: how specific is it?’, Journal of
Developmental Psychology, vol. 9, pp. 301-314.
Baron-Cohen, S., Knickmeyer. R. & Belmonte, M. K. 2005, ‘Sex differences in the brain: implications
for explaining autism’, Science, vol. 310, pp. 819-823.
Baron-Cohen, S., Wheelwright, S., Spong, A., Scahill, V. & Lawson, J. 2001, ‘Studies of the theory of
mind: are intuitive physics and intuitive psychology independent?’, Journal of Developmental
Learning Disabilities, vol. 5, pp. 47-78.
Buitelaar, J. K., van der Wees, M., Swaab-Barneveld, H. & van der Gaag, R. J. 1999, ‘Verbal memory and
performance IQ predict theory of mind and emotion recognition ability in children with autistic
spectrum disorders and in psychiatric control children’, Journal of Child Psychology and Psychiatry,
vol. 40, pp. 869-881.
Critchley, H. D., Daly, E. M., Bullmore, E. T., et al. 2000,’The functional neuroanatomy of social
behaviour. Changes in cerebral blood flow when people with autistic disorder process facial
expressions’, Brain, vol. 123, pp. 2203-2212.
Donnelly, S. L., Wolpert, C. M., Menold, M. M., et al. 2000, ‘Female with autistic disorder and
monosomy X (Turner syndrome): parent-of-origin effect of the X chromosome’, American
Journal of Medical Genetics, vol. 96, pp. 312-316.
Fombonne, E. 1999, ‘The epidemiology of autism: a review’, Psychological Medicine, vol. 29, pp. 769-786.
Happé, F., Ronald, A. & Plomin, R. 2006, ‘Time to give up on a single explanation for autism’, Nature
Neuroscience, vol. 9, pp. 1218-1220.
Hardan, A.Y., Muddasani, S., Vemulapalli, M., Keshavan, M. S. & Minshew, N. J. 2006, ‘An MRI study of
increased cortical thickness in autism’, American Journal of Psychiatry, vol. 163, pp. 1290-1292.
Hollander, E., Bartz, J., Chaplin, W., et al. 2007, ‘Oxytocin increases retention of social cognition in
autism’, Biological Psychiatry, vol. 61, pp. 498-503.
Jacob, S., Brune, C. W., Carter, C. S., Leventhal, B. L., Lord, C. & Cook, E. H. 2007, ‘Association of
the oxytocin receptor gene (OXTR) in Caucasian children adolescents with autism’, Neuroscience
Letters, vol. 417, pp. 6-9.
Just, M. A., Cherkassky, V. L., Keller, T.A., Kana, R. K. & Minshew, N. J. 2007,’Functional and
anatomical cortical underconnectivity in autism: evidence from an fMRI study of an executive
function task and corpus callosum morphometry’, Cerebral Cortex, vol. 17, pp. 951-961.
Kanner, L. 1972, Child Psychiatry, 4th edn, Charles C. Thomas, Springfield IL.
Keller, T. A., Kana, R. K. & Just, M. A. 2007,’A developmental study of the structural integrity of
white matter in autism’, Neuroreport, vol. 18, pp. 23-27.
Kilpinen, H., Ylisaukko-Oja, T., Hennah, W., et al. 2008, ‘Association of DISCI with autism and
Asperger syndrome’, Molecular Psychiatry, vol. 13, pp. 187-196.
Macintosh, K. & Dissanayake, C. 2006, ‘A comparative study of the spontaneous social interactions
of children with high-functioning autism and children with Asperger’s disorder’, Autism,
vol. 10, pp. 199-220.
McCracken, J. T., McGough, J., Shah, B., et al. 2002, ‘Risperidone in children with autism and serious
behavioral problems’, New England Journal of Medicine, vol. 347, pp. 314-321.
Muhle, R., Trentacoste, S. V. & Rapin, I. 2004, ‘The genetics of autism’, Pediatrics, vol. 113, pp. 472-486.
Perner, J., Frith, U., Leslie, A. M. & Leekam, S. R. 1989, ‘Exploration of the autistic child’s theory of
mind: knowledge, belief, and communication’, Child Development, vol. 60, pp. 689-700.
Sally, D. & Hill, E. 2006, ‘The development of interpersonal strategy: autism, theory-of-mind,
cooperation and fairness’, Journal of Economic Psychology, vol. 27, pp. 73-97.
Skuse, D. H., James, R. S., Bishop, D. V. M., et al. 1997, ‘Evidence from Turner’s syndrome of an
imprinted X-linked locus affecting cognitive function’, Nature, vol. 387, pp. 705-708.
Snyder, A. W. & Thomas, M. 1997, ‘Autistic artists give clues to cognition’, Perception, vol. 26, pp. 93-96.
Tinbergen, E. A. & Tinbergen, N. 1972, Early Childhood Autism-an Ethological Approach,
Paul Parey, Berlin.
Van Ijzendoorn, M. H., Rutgers, A. H., Bakermans-Kranenburg, M. J., et al. 2007, ‘Parental sensitivity
and attachment in children with autism spectrum disorders: comparison with children with
mental retardation, with language delays, and with typical development, Child Development,
vol. 78, pp. 597-608.
Williams, J. H. G., Whiten, A., Suddendorf, T. & Perrett, D. I. 2001, ‘Imitation, mirror neurons and
autism’, Neuroscience and Biobehavioral Reviews, vol. 25, pp. 287-295.
この文章は自閉症スペクトラム障害(ASD)の基本的な特性と、特に社会的行動における特定の見解について説明しています。詳しく解説します:
自閉症スペクトラム障害の多様性
文章の冒頭で述べられているように、自閉症スペクトラム障害は非常に多様な症状と表現型を持ちます。「スペクトラム」という用語自体が、軽度から重度まで幅広い症状の連続体があることを示しています。
社会的脳機能への選択的影響
自閉症は以下の3つのレベルで社会的脳機能に特異的な影響を与えるとされています:
- 行動レベル:社会的相互作用や他者との関わり方に影響
- 認知レベル:社会的情報の処理や理解の仕方に影響
- 感情レベル:社会的状況における感情の処理や表出に影響
これは、自閉症が一般的な認知機能全体に影響するのではなく、特に「社会的脳」と呼ばれる神経回路に関連する機能に選択的に影響することを示唆しています。
動機づけの葛藤モデル
この文章で提示されている特に興味深い観点は、軽度の自閉症における「動機づけの葛藤」の存在です:
- 観察に基づく仮説:行動学的観察から得られた知見
- 葛藤の性質:
- 接近動機:養育者に近づきたい、社会的つながりを求める欲求
- 回避動機:同時に社会的相互作用を回避したい欲求
- 葛藤の原因:この回避行動は「高まった臆病さと恐怖」に起因する可能性がある
この仮説の重要性
このモデルは従来の自閉症理解に重要な視点を追加します:
- 自閉症の人々が社会的関わりを「望んでいない」のではなく、それを「恐れている」可能性がある
- 社会的相互作用への関心の欠如ではなく、不安や恐怖が社会的行動を妨げている可能性がある
- この理解は、特に軽度の自閉症の治療アプローチに影響を与える可能性がある
この「接近-回避葛藤」モデルは、治療介入において社会的不安の軽減が重要である可能性を示唆しています。安全で予測可能な社会的環境を提供することで、自閉症の人々の社会的関わりを促進できる可能性があります。
この視点は、自閉症を単に「社会的関心の欠如」として捉えるのではなく、より複雑な動機づけの問題として理解することを促し、より効果的な支援方法の開発につながる可能性があります。
この文章は自閉症児の社会的接触と愛着形成の課題、およびその生物学的・関係的基盤について説明しています。詳しく解説します:
近接接触の回避と早期徴候
- 自閉症児は非常に早い発達段階から近接接触の回避を示す
- これは人との物理的な近さや触れ合いを避ける傾向を指します
- この早期の回避行動が、子どもと養育者の間の情緒的絆の確立を困難にする
愛着スタイルへの影響
- 研究結果:自閉症児は健常児と比較して不安定な愛着スタイルが一般的
- 愛着理論では、安定型、回避型、両価型、無秩序型などの愛着パターンがあり、自閉症児では安定型以外の愛着パターンが多く見られる
生物学的基盤(近因レベル)
この現象に関連する可能性のある生物学的要因:
- オキシトシンの代謝回転の変化
- オキシトシンは「絆形成ホルモン」とも呼ばれ、社会的結合に重要
- セロトニンの代謝回転の変化
- セロトニンは気分や社会的行動の調節に関与する神経伝達物質
養育者の反応の影響
養育者の対応は状況を良くも悪くもしうる:
有効な対応
- 子どもを社会的相互作用に強制しない
- 子どものペースを尊重することで絆の形成を促進
悪化させうる対応
- 視線を合わせようとする繰り返しの試み
- 物理的な近接を強引に確立しようとする行動
- これらは子どもの不安や回避傾向を強める可能性がある
親の敏感さについての誤解を正す
- 重要な指摘:自閉症児の親は、平均して健常児の親よりも敏感さが低いわけではない
- これは、自閉症の原因を「冷淡な母親」に求めた歴史的に誤った「冷蔵庫マザー」理論を否定する知見
問題の所在
- 愛着形成の困難さは主に子ども側の要因による可能性が高い
- 子どもの社会的シグナルの発信・反応の特異性が関係している
重度の自閉症の場合
- 最も深刻なケースでは、子どもと養育者の間の情緒的絆が決して確立されない可能性もある
- これは自閉症の症状の重さによって、相互的な社会的関係の基盤そのものが形成されにくい場合があることを示唆
この内容は、自閉症における社会的関係の困難さが単なる「無関心」ではなく、生物学的基盤を持つ複雑な現象であること、そして養育者のアプローチが重要でありながらも、問題の本質は子どもの神経発達的特性にあることを示しています。適切な介入は、子どもの特性を理解し、その不安や恐怖を悪化させない配慮が必要であることが示唆されています。
この文章は、自閉スペクトラム症(ASD)の中でも「高機能自閉症」や「アスペルガー症候群」と診断される子どもたちや成人が、社会的相互作用や人間関係の構築において直面する困難について述べています。以下に内容を詳しく解説します。
1. 学齢期における社会的遊びと孤立
◆ 高機能自閉症・アスペルガー症候群の特徴:
- 一般的に知的発達には遅れがない(平均〜平均以上のIQ)ため、見た目や学力だけでは「障害」とは認識されにくい。
- しかし、社会的コミュニケーションや対人関係のスキルに顕著な困難があります。
◆ 社会的遊びの参加が少ない理由:
- 子ども同士の**非言語的なやり取り(表情・ジェスチャー・視線の読み取りなど)**が苦手。
- 「空気を読む」「相手の気持ちを推測する」といった暗黙のルールを理解することが難しい。
- 興味や関心が限定的で、同年齢の子と関心を共有しにくい。
- 自分のやり方に強いこだわりがあるため、共同作業やルールのある遊びが苦手なことも多い。
◆ 結果として:
- 他の子どもたちとの自然な遊びの輪に入りにくくなる。
- 遊びに誘われてもどう振る舞えばよいか分からず、回避することが多くなる。
- 周囲から**「変わった子」と見なされ、孤立する**ことも少なくない。
2. 成人期における親密な人間関係の困難
◆ 成長しても残る対人関係の困難:
- 自閉スペクトラム症の特性は、年齢を重ねても完全には消えない。
- 成人になると、対人関係の質がより深く複雑になる(友人関係から恋愛関係、職場での人間関係など)。
◆ 特に難しいとされるのは「親密さの形成」:
- 会話のキャッチボールや相手への配慮など、双方向的な関係性の維持が難しい。
- 感情表現が乏しい、または過剰で、誤解されることがある。
- 自他の境界が曖昧で、距離感をつかむのが苦手なこともある。
- 人との関わりそのものが負担やストレスになる場合もある。
◆ 社会的孤立や孤独感につながる:
- 社交の場に出ることが減り、孤独感を抱えやすい。
- 親密なパートナー関係(恋愛・結婚など)を築くのが困難で、生活上のサポートが得にくくなる。
- 一部の人は、自ら人間関係を避ける傾向もあり、社会的に孤立するケースもあります。
このように、自閉スペクトラム症の人々(とくに高機能・アスペルガー型)の社会的困難は、子ども時代から大人まで一貫して見られるものであり、支援や理解が長期的に必要です。
この文章は、自閉スペクトラム症(ASD)の人々が持つ**「心の理論(Theory of Mind)」の障害**について、近年の研究成果をもとに詳細に説明しています。それを分かりやすく噛み砕いて解説します。
🔍 概要:自閉症と「心の理論」の障害
◆ 「心の理論(Theory of Mind)」とは?
- 他人には自分とは異なる考えや感情、意図、信念があると理解する能力。
- つまり、「あの人はこう思っているんじゃないか」「こう感じているかもしれない」と他者の内面を推測する能力です。
- 心の理論は、人間関係を築いたり、社会的に適切に振る舞うために必要不可欠なスキルです。
🧠 ASDにおける「心の理論」障害の特徴
◆ 多くの研究が一貫して示す問題点:
- 自閉症の人々は、他人の考えや感情を推測するのが苦手です。
- 相手の表情や態度を手がかりに、「今どう思っているのか」「何を望んでいるのか」などを理解するのが難しい。
- これにより、思考・意図・感情・欲求・気質などを把握するのが困難になります。
🚫 他の認知機能障害とは異なる
◆ 重要なポイント:
「この困難は単なる知的障害や記憶力の問題ではない」
- 実行機能(計画性や注意のコントロールなど)に課題がある場合もありますが、「心の理論」の障害はそれとは独立した、より“選択的な”障害です。
- たとえば:
- 自伝的記憶(過去の経験を物語として再構成する力)や
- 実行機能の障害(計画の立案・遂行)が見られることもあるが、
- それらが主因ではなく、特に社会的な情報処理能力に限って障害があるという点が注目されています。
👥 社会的情報処理の障害の具体例
以下の能力が選択的に障害されていることが報告されています:
- メンタライジング
→ 他人の内面を推測する能力の障害 - 感情認識
→ 表情や声のトーンから感情を読み取る力の障害 - 顔の処理
→ 顔を見分けたり、視線の方向から相手の関心を把握する力の障害
⚖️ 社会的規範(公平性・互恵性)への理解の困難
◆ 社会的規則や価値観の理解にも影響:
- 公平性(「みんなに平等に分けるべき」など)
- 互恵性(「自分がしてもらったことを他人にも返すべき」)
こういった社会の中で共有されている“ルール”や“暗黙の了解”を直感的に理解するのが難しいとされています。
例:
- 誕生日パーティーでプレゼントをもらったら、「ありがとう」と返すのが当たり前とされる社会的期待。
- 自閉症の人は、それがなぜ必要かが分からなかったり、気づかなかったりする。
✅ まとめ
この文章で述べられていることは以下のように整理できます:
| 項目 | 内容 |
|---|---|
| 問題の中心 | 他人の心を読む「心の理論」能力の障害 |
| 影響を受ける能力 | 思考・感情・意図・欲求などの理解、感情認識、顔の処理 |
| 他の認知機能との関係 | 記憶力や実行機能とは別の、選択的な社会的障害 |
| 結果として起きること | 公平性や互恵性など、社会的なルールの理解が困難になる |
この文章は、自閉スペクトラム症(ASD)におけるメンタライジング(=他者の心の状態を理解する能力)の障害が、知的能力とは必ずしも関係していないこと、また、たとえ軽度であっても自閉症の特性が自然に「成長とともに治る」ものではないということを強調しています。
以下に、ポイントごとに詳しく説明します。
🔹 自閉症の「メンタライジングの障害」は知能とは別の問題
◆ 知能(IQ)が高くてもメンタライジングが苦手
- 一般には、「頭が良ければ人の気持ちもよく分かる」と思われがちですが、自閉症の場合はそうとは限りません。
- 自閉症の中には、平均以上のIQや優れた学力を持つ人も多くいます(特に「高機能自閉症」や「アスペルガー症候群」など)。
- しかし、そうした人々でも、他者の考え・感情・意図を読み取る社会的認知は未発達なことが多いです。
➤ なぜか?
- 知能検査は、記憶力・計算力・言語理解など「論理的な認知能力」を測ります。
- 一方でメンタライジングは、**社会的な文脈や人間関係に関する「暗黙の理解」や「感情の読解」**といった、別の能力です。
- つまり、知的に優秀でも「心の理論」のスキルは育たないことがあるのです。
🔹 知能が高くても「社会的認知の発達の遅れ」が顕著
- 特にIQが高い軽度の自閉症の人では、学校の勉強や知識の獲得には問題がない一方で、
- 他者の感情を読み取る
- 空気を読む
- 状況に合わせた柔軟な対人行動をとる
といった、「社会的知性(social intelligence)」にあたる部分が著しく発達しにくいことがしばしば報告されています。
➤ 結果として:
- 「勉強はできるけど、友だちと関係を築けない」
- 「職場での雑談やチームワークが苦手」
- 「周囲に誤解されやすい」
などの社会的困難を、知能に関係なく経験することになります。
🔹 「軽度の自閉症」でも自然に解消されるわけではない
◆ 年齢とともに改善するケースもあるが…
- 年齢が上がると、訓練や経験を通して表面的な社会的スキルを学ぶことは可能です。
- たとえば、よく使われるフレーズを覚えたり、相手の表情をある程度パターン化して理解したり。
◆ しかし根本的な「社会的な直感」や「共感性」は自動的には発達しない
- **多くの場合、社会的スキルは“意識して努力して身につけるもの”**となり、直感的・自然なものではありません。
- また、ストレスがかかった場面では、学習したスキルがうまく使えなくなることもあります。
➤ つまり:
- 自閉症の特性は、単なる「子どもらしい未熟さ」ではなく、生涯にわたる神経発達の違いに基づくもの。
- 軽度であっても、「放っておけば大人になれば自然に治る」という見方は誤解です。
✅ まとめ
| ポイント | 説明 |
|---|---|
| 知能とメンタライジングの関係 | IQが高くても心の理論は発達しにくい |
| 社会的認知の遅れ | 特に他人の気持ちや視点を理解する力が育ちにくい |
| 自然な成長での解消 | 年齢とともに表面的な改善はあり得るが、本質的な困難は残る |
| 支援の必要性 | 成長とともに「治る」ものではなく、継続的な理解と支援が必要 |
この部分は、自閉症スペクトラム症(ASD)における社会的孤立や行動異常の根本原因を説明しようとする議論です。キーワードは「メンタライジングの障害」と「ミラーニューロンシステムの機能不全」です。それぞれを詳しく解説し、なぜメンタライジングの障害だけでは説明が不十分なのかを明らかにします。
🧠 1. メンタライジングの発達段階とその限界
◆ メンタライジング(心の理論)の発達
- 前述のように、通常の発達では3〜5歳頃にメンタライジング能力(他者の信念や意図の理解)が発達します。
- これにより、他人との関係で「なぜそう感じたか」「何を考えているか」を理解できるようになります。
◆ 自閉症における障害
- 自閉症ではこの能力の発達が遅れたり、質的に異なっていたりするため、社会的なズレや孤立が生じます。
◆ しかし、問題はここからです:
メンタライジングが通常3〜5歳で発達するにもかかわらず、自閉症の子どもたちはそれ以前、つまり乳幼児期(1〜2歳)からすでに社会的行動の異常を示している。
🔍 つまり:
- **メンタライジングの障害は結果の一部にすぎず、もっと早期の脳や神経の異常が背景にあるのでは?**という疑問が生まれます。
🧠 2. ミラーニューロンシステム(Mirror Neuron System: MNS)の役割
◆ ミラーニューロンとは?
- 他人の動作を見ているだけで、自分の脳内で同じ神経活動が起きるというニューロン。
- 例:誰かがコップを持ち上げるのを見ると、自分も同じ動作をするつもりになったかのような脳活動が起こる。
◆ ミラーニューロンシステムの主な働き
- 模倣(他人の行動をまねる)
- 意図の理解(なぜその行動をしたのか、目的を推測する)
- 感情の共感(他人の表情や仕草に共鳴する)
- 社会的学習(直接体験せずに他者の行動から学ぶ)
◆ 自閉症との関連
- ASDでは、このミラーニューロンシステムの活動が低下しているという研究報告が多数あります。
- その結果、模倣の困難・表情の理解の弱さ・共感の乏しさが生じると考えられています。
🔄 3. 結論:なぜメンタライジング障害だけでは説明できないのか
| 観点 | 説明 |
|---|---|
| 🧩 時期の不一致 | メンタライジング能力は通常3〜5歳に発達するが、ASDでは1歳台から社会的行動の異常が出現する(例:アイコンタクトや共同注意の欠如)。 |
| 🔍 根本原因の仮説 | より初期に機能するミラーニューロンシステムの異常が、社会的情報処理の土台を揺るがしている可能性がある。 |
| 🔗 連続性の理解 | ミラーニューロンの異常 → 模倣・共感・意図理解の困難 → メンタライジングの発達障害 → 社会的孤立や対人関係の困難、という発達的連鎖があると考えられる。 |
📌 補足:ミラーニューロン仮説の現在の位置づけ
- 一時期、「ミラーニューロン仮説=自閉症の原因」として注目を浴びました。
- しかし現在は、「原因の一つの要素であり、複数の脳機能の異常の中の一部」という位置づけにあります。
- ただし、早期の社会的情報処理能力の低下や感覚過敏といった、行動異常の始まりに大きく関与している可能性は強く支持されています。
✅ まとめ
- メンタライジング能力の障害だけでは、自閉症における社会的困難の早期出現(1〜2歳から)を説明できない。
- より根本的な神経システム、特にミラーニューロンシステムの機能不全が、その原因として関与している可能性がある。
- これは、自閉症の社会的障害を発達の流れ全体で捉える視点を提供する重要な知見です。
以下に、ご提示の内容をわかりやすく詳しく解説し、見出しに H4(####) を使用して整理しました。
ミラーニューロンの発見と特徴
ミラーニューロン(mirror neurons)は、1990年代初頭にイタリアのパルマ大学の研究チーム(リッツォラッティら)によって、**マカクザルの脳(運動前野F5)**で発見された神経細胞です。驚くべきことに、サルが自分で物をつかむ動作を行うときと、他者(人間や他のサル)が同じ動作をしているのを見るときの両方で、同じニューロンが活動したのです。
この性質により、「鏡のように相手の行動を脳内で写し取る」ニューロン=ミラーニューロンと名付けられました。
模倣と学習におけるミラーニューロンの役割
ミラーニューロンは、単なる動作の感知ではなく、「理解」や「学習」と深く結びついていると考えられています。具体的には:
- 他者の動作を見ただけで、自分自身がそれを実行する準備をするような神経活動が起こる。
- この働きによって、**模倣による学習(観察学習)**が成立する。
つまり、ミラーニューロンの働きは、人間や動物が他者の行動を観察して、それを自分の行動に取り入れる(模倣する)ことを可能にしているという点で極めて重要です。
視覚的な手がかりがなくても活動する特性
ミラーニューロンは、実際の動作を視覚的に完全に見えなくても発火することが知られています。たとえば:
- サルが「実験者が箱の中に手を入れて何かを取る」という行動を見えない状態(箱の中)で観察しても、ミラーニューロンは活性化します。
これは、ミラーニューロンが単なる視覚的模倣ではなく、「他者の意図」や「行動の予測」を行っていることを示しています。
種間比較とヒト以外の霊長類からの証拠
ミラーニューロンの存在や働きは、ヒトだけでなく、サルや類人猿にも確認されており、「社会的学習や文化の伝達がなぜ可能になるのか」という問いに対する神経学的な根拠を提供しています。
種間比較(ヒト vs 他の霊長類)の研究では:
- チンパンジーやオランウータンも、道具の使い方や社会的ルールを観察学習によって学ぶことが示されています。
- これらの能力は、模倣・予測・共感的理解といった、ミラーニューロンが関与すると考えられる高度な認知機能と関係している可能性が高いです。
ミラーニューロンと認知予測の統合
このように、ミラーニューロンは単に「見た動作を真似る」だけではなく、動作の「意味」や「目的」を理解し、行動を予測する脳の回路と深くつながっています。この点で、以下の2つの認知機能をつなぐハブのような役割を果たしていると考えられます:
- 視覚的認知(他者の行動の観察)
- 予測と理解(他者の意図・結果の推測)
この働きが、人間の社会性や文化形成、さらにはメンタライジング能力の基礎になっているという見方が広がっています。
以下では、ミラーニューロンに関連するヒトの脳領域について詳しく解説し、対応する図解の内容も含めて説明します。
🧠 ミラーニューロンに関わる主な脳部位(ヒトの場合)
ミラーニューロンが最初に発見されたのはサルの脳(F5野)でしたが、ヒトにおいては以下の脳領域が対応すると考えられています:
#### 1. 下前頭回(Inferior Frontal Gyrus, IFG)
- 位置:前頭葉の下部、左半球でブローカ野に近い領域。
- 役割:言語、運動の模倣、行動意図の理解に関与。
- ミラーニューロンの主要な活動部位とされ、他者の動作や意図を理解する際に強く活性化します。
- ヒトの社会的コミュニケーションにおいて重要な場所でもあります。
#### 2. 下頭頂小葉(Inferior Parietal Lobule, IPL)
- 位置:頭頂葉の下側で、後頭葉と前頭葉の間に位置。
- 役割:
- 自他の動作の統合。
- 他者の動作の模倣と予測。
- 他者の行動を「意味づけ」し、「その動作が何を意図しているか」を推測する処理を担っています。
#### 3. 上側頭溝(Superior Temporal Sulcus, STS)
- 位置:側頭葉の上部の溝部分。
- 役割:
- 他者の視線、顔の向き、身体の動きなどの生物的運動の知覚に関与。
- IFGやIPLと連携し、動作の理解の補助的な情報処理を行う。
🧩 この3領域で構成される「ミラーニューロンシステム」
これら3つの領域(IFG・IPL・STS)は密接に結びついており、人間の模倣・共感・行動理解の神経基盤として「ミラーニューロンシステム(Mirror Neuron System, MNS)」を構成しています。
🧭 図解イメージの説明(※図が必要であれば生成・提供可能)
🧠 左側面から見た脳の図(簡易的に説明):
┌────────────────────────────┐
│ 脳の左側面(側面図) │
│ │
│ ┌────────────┐ │
│ │ 上側頭溝(STS) │ ◀ 生物的運動の知覚 │
│ └────────────┘ │
│ ▲ │
│ │ │
│ ┌────────────┐ │
│ │ 下頭頂小葉(IPL) │ ◀ 意図理解・予測 │
│ └────────────┘ │
│ ▲ │
│ │ │
│ ┌────────────┐ │
│ │ 下前頭回(IFG) │ ◀ 模倣・動作理解 │
│ └────────────┘ │
└────────────────────────────┘
🔄 これらの部位の連携の例
例:誰かがコップを持ち上げるのを見ると…
- STSがその動作を視覚的に検出。
- IPLがその動作の目的(飲む?移動?)を推測。
- IFGがその行動を模倣・内的に再現し、理解しようとする。
この一連の処理が「他者の行動を自分の脳でシミュレートする」メカニズムと考えられています。
🧠 追加ポイント:左半球優位 vs 右半球の役割
- 通常、左半球のIFGやIPLが言語や道具の使用に関連する模倣に強く関わります。
- 一方で、右半球の対応部位は表情や感情などの非言語的コミュニケーションの模倣や共感に関与する傾向があります。
✅ まとめ
| 脳領域 | 主な機能 | ミラーニューロンとの関係 |
|---|---|---|
| 下前頭回 (IFG) | 模倣、行動理解 | 他者の動作を再現・理解する中心 |
| 下頭頂小葉 (IPL) | 意図の予測、模倣 | 動作の目的を推論する |
| 上側頭溝 (STS) | 生物的運動の認知 | 視覚的な社会情報を提供 |
以下に、あなたが提示された内容を、専門的かつわかりやすく詳しく解説します。
🧠 メンタライジングの前駆体としての模倣
### 🔹 行動の模倣と初期学習
- **行動の模倣(imitation)**とは、他者が行っている行為を観察し、それを再現する能力です。
- この模倣行動は、乳児期から始まる初期の社会的学習にとって極めて重要です。たとえば、赤ちゃんは親の表情や動作をまねることで、言語やジェスチャーを学びます。
### 🔹 模倣能力は「心の理論(メンタライジング)」の前段階
- 模倣は単に「真似する」だけでなく、他者の行為の意味や意図を理解する基盤でもあります。
- 特に重要なのは、「他者の行動を自分の中で再現(シミュレート)すること」が、後のメンタライジング能力の発達に必要だという点です。
🧠 メンタライジングとは?
- メンタライジング(Mentalizing)とは、他者の信念・意図・感情・欲求といった心的状態を推論する能力です。
- たとえば、誰かが困っている様子を見て、「この人は助けを求めているかもしれない」と理解することがメンタライジングの一例です。
🧭 メンタライジングには追加的な認知処理が必要
模倣とメンタライジングは連続的な発達段階と考えられていますが、メンタライジングには模倣以上に高度な認知メカニズムが必要です:
| 認知メカニズム | 説明 |
|---|---|
| 自己と他者の区別 | 他者の感情・行動を想像しても、それが「自分のものではない」と理解する力。 |
| 現実と見かけの区別 | 他者が持っている誤った信念(たとえば「箱の中にチョコがあると思っている」)を、自分の正しい知識と切り分ける力。 |
こうした**視点取得(perspective-taking)**や、**誤信念理解(false belief understanding)**が、模倣ではなくメンタライジング特有の高度な機能です。
🧠 自閉症における模倣とメンタライジングの発達不全
自閉スペクトラム症(ASD)の人々においては、以下のような模倣とその前提能力の発達障害が見られます:
- 視線の追跡(gaze following):
- 他者がどこを見ているかに注意を向ける能力。
- 共同注意(joint attention):
- 他者と一緒に何かに注意を向ける、という初期の社会的相互作用。
これらは通常、生後6~12か月の間に発達しますが、自閉症の子どもではこの段階からつまずきが見られます。
⚠ ミラーニューロンの機能不全とその二面性
🔹 神経活動の「抑制」:
- ミラーニューロンシステムが**十分に働かない(抑制)**場合、他者の行動の理解・共感・意図の推測が難しくなり、メンタライジングや視点取得が困難になると考えられます。
- これは、自閉症の「社会的無関心」や「心の理論の障害」と一致します。
🔹 神経活動の「脱抑制」:
- 一方で、ミラーニューロンシステムが**過剰に働く(脱抑制)**場合、病的な模倣行動が現れることがあります。
- エコプラクシア(echopraxia):他人の動作を自動的に真似してしまう。
- エコラリア(echolalia):他人の言葉を繰り返してしまう。
- これは、自閉症や発達障害に見られる現象で、模倣が制御不能になっている状態です。
✅ まとめ
| 項目 | 内容 |
|---|---|
| 模倣 | 行動の再現。社会的学習の基本で、メンタライジングの前提。 |
| メンタライジング | 他者の心的状態を理解・予測する高度な認知能力。 |
| 自閉症との関係 | 早期の模倣・共同注意の発達不全 → メンタライジング能力の障害へとつながる。 |
| ミラーニューロンの役割 | 模倣と意図理解を神経的に仲介する。機能不全が両方向の症状を説明しうる。 |
この部分は、自閉症の特徴的な症状である他者の意図や感情の理解の困難さ(メンタライジングの障害)が、脳の構造的および機能的な異常と関連している可能性を示唆しています。具体的には、以下の3つのポイントが挙げられます。
1. ミラーニューロンを含む脳領域の皮質厚の減少:
- ミラーニューロンシステムとは: ミラーニューロンは、自身が行動を行うときと、他者が同様の行動を行うのを見たときに活動する特殊な神経細胞群です。このシステムは、他者の行動の意図を理解したり、共感したりする上で重要な役割を果たすと考えられています。
- 自閉症における皮質厚の減少: ご提示の記述によれば、自閉症の方では、ミラーニューロンが多く存在すると推定される前頭前皮質と側頭領域において、皮質の厚さが減少しているという観察があります。皮質の厚さは、その領域の神経細胞の密度や神経線維の量と関連すると考えられており、この減少はミラーニューロンシステムの機能低下を示唆する可能性があります。
- 他の部位との対比: 興味深いことに、自閉症ではこれらの領域とは対照的に、他の脳部位では皮質の厚さが増加しているという報告もあります。このことは、自閉症が脳全体に一律の影響を与えるのではなく、特定の領域に選択的な影響を与える可能性を示唆しています。
2. メンタライジングに関わる脳領域の活動低下:
- メンタライジングに関わる脳領域: メンタライジング、つまり「心の理論」とも呼ばれる、他者の心的な状態(意図、感情、信念など)を推測する能力は、社会的な相互作用において非常に重要です。ご提示の記述では、機能的脳イメージング研究によって、メンタライジングに一貫して関与している脳領域(具体的には前帯状皮質(ACC)と傍帯状皮質)において、活動の低下が観察されていると述べられています。
- 異常な活性化パターン: これらの領域の活動低下は、自閉症の方々が他者の心を理解する際に困難を抱える神経基盤の一つであると考えられます。
3. セロトニン神経系の関与:
- セロトニン神経の分布: 前帯状皮質(ACC)と傍帯状皮質は、セロトニンという神経伝達物質を放出する神経細胞(セロトニン神経)が密に分布している領域です。セロトニンは、気分、感情、社会行動など、様々な脳機能に影響を与えることが知られています。
- 前頭前皮質におけるセロトニン機能不全: ご提示の記述は、前頭前皮質におけるセロトニンの機能不全が、自閉症においてこれらのメンタライジングに関わる脳領域の機能を抑制している可能性を示唆しています。セロトニンの異常が、ミラーニューロンシステムを含む前頭前皮質の機能低下、ひいてはメンタライジング能力の障害につながるという仮説です。
まとめ:
この部分は、自閉症における社会性の困難さの神経基盤として、ミラーニューロンシステムの機能低下とメンタライジングに関わる脳領域の活動異常に焦点を当てています。さらに、これらの異常には、前頭前皮質におけるセロトニン神経系の機能不全が関与している可能性が示唆されています。これは、自閉症の複雑な病態を理解する上で、脳の構造、機能、神経化学的な側面を統合的に捉えることの重要性を示しています。
この文章は、自閉症の脳が「男性脳」の極端な変異であるとする仮説(極端男性脳仮説、Extreme Male Brain Theory)を説明しており、性差、認知特性、胎児期のテストステロン、および遺伝的要因がどのように関与するかを議論しています。以下で、内容を詳しく解説します。
1. 性差と認知特性
文章では、男性と女性の間で認知能力や行動特性に平均的な違いがあると指摘しています。これらの違いは、進化的な視点や脳の発達に関連すると考えられています。
- 女性の優位性:
- 共感的な視点取得(Empathetic Perspective-Taking):他者の感情や視点を理解する能力。
- メンタライジング(Mentalizing):他者の意図や信念を推測する能力(心の理論)。
- 社会的感受性(Social Sensitivity):社会的状況や非言語的手がかりを敏感に察知する能力。
- 言語流暢性(Verbal Fluency):言葉を流暢に使い、コミュニケーションを行う能力。
- これらの特性は、女性が子育てや社会的な関係構築において多くの「親の投資」(時間、エネルギー、ケア)を行う役割を進化的に担ってきたため、発達した可能性があるとされています。
- 男性の優位性:
- 心的回転(Mental Rotation):3次元オブジェクトを頭の中で回転させる空間的思考能力。
- 空間定位(Spatial Orientation):空間内での方向感覚や位置把握の能力。
- 物理的問題解決(Physical Problem-Solving):機械や物理的なシステムを理解し、操作する能力。
- これらの特性は、狩猟や道具使用、競争などの役割に関連して進化した可能性があります。
これらの性差は、脳の構造やホルモン環境の違いによるものと考えられています。
2. 自閉症と「極端男性脳仮説」
自閉症スペクトラム障害(ASD)は、社会的コミュニケーションの困難や、限定的・反復的な行動パターンを特徴とします。文章では、自閉症の脳が「男性脳」の特性を極端に強調した形態であるとする仮説を提示しています。この仮説は、サイモン・バロン=コーエン(Simon Baron-Cohen)によって提唱されたものです。
- 自閉症の認知特性:
- 社会的認知の障害:共感やメンタライジングが特に困難。たとえば、他者の感情や意図を理解するのが難しい。
- 技術的理解の保存または優位性:一方で、システム化(物事の規則性やパターンを理解する能力)や空間的・機械的問題解決において、平均以上の能力を示す場合がある。
- 例:自閉症の人は、数学、エンジニアリング、コンピュータプログラミング、または細かいパターン認識(例:時刻表や地図)に強い興味や能力を持つことがあります。
- 「男性脳」との関連:
- 男性脳は、共感(Empathizing)よりもシステム化(Systemizing)に優れる傾向があるとされています(バロン=コーエンのE-S理論)。
- 自閉症の人は、システム化が極端に強く、共感が極端に弱い傾向があるため、「男性脳」の特性が極端に強調された状態と考えられる。
- 例:自閉症の人は、機械や数字のパターンに強い関心を持つ一方で、社会的相互作用や感情の共有に困難を抱えることが多い。
3. 胎児テストステロンの影響
この仮説では、胎児期のテストステロン(男性ホルモン)への過剰な曝露が、自閉症の脳の発達に影響を与える可能性があるとされています。
- テストステロンの役割:
- 胎児期にテストステロンが多く分泌されると、脳の構造や機能に影響を与える。特に、大脳皮質や扁桃体、海馬などの領域に影響を及ぼす。
- テストステロンは、共感や社会的認知に関連する脳領域の発達を抑制し、空間的・システム的思考に関連する領域を強化する可能性がある。
- 自閉症の人は、胎児期にテストステロンへの曝露が過剰だったため、「男性脳」の特性が極端に現れると仮説づけられている。
- 証拠:
- 研究では、自閉症の人は胎児期のテストステロンレベルが高い可能性が示唆されている(例:羊水中のテストステロン濃度と自閉症の関連)。
- 性差の大きい特性(例:空間能力や共感の欠如)が自閉症で強調されることも、この仮説を支持する間接的証拠とされる。
4. 父性インプリント遺伝子の関与
文章では、自閉症の脳の発達が「父性インプリント遺伝子」の影響を受ける可能性にも触れています。
- インプリント遺伝子とは:
- インプリント遺伝子は、親のどちらか(父親または母親)からのみ発現する遺伝子。遺伝子の発現が親の性別に依存する。
- 父性インプリント遺伝子は、父親由来の遺伝子が優先的に発現するもので、胎児の成長や脳の発達に影響を与える。
- 自閉症との関連:
- 父性インプリント遺伝子が過剰に発現すると、テストステロン関連の脳発達が促進され、「男性脳」の特性が強調される可能性がある。
- 一部の研究では、自閉症に関連する遺伝子変異が、父性インプリント遺伝子と関連している可能性が示唆されている。
- 例:特定の遺伝子(例:UBE3Aや他のインプリント遺伝子)が自閉症やその特性に影響を与える可能性。
5. 仮説の意義と限界
- 意義:
- この仮説は、自閉症の性比(男性に多く、4:1程度)の説明や、認知特性の理解に役立つ。
- 自閉症の多様な症状を、進化的な性差やホルモン環境の観点から統合的に説明する枠組みを提供する。
- 治療や支援の開発において、性差やホルモン環境を考慮する重要性を示唆する。
- 限界:
- 自閉症は非常に多様で、全員が「極端男性脳」の特性を示すわけではない(例:女性の自閉症者は異なる表現型を示すことが多い)。
- 胎児テストステロンの影響やインプリント遺伝子の役割は、仮説の域を出ておらず、因果関係の証明にはさらなる研究が必要。
- 社会的・環境的要因(例:育児環境や教育)が自閉症の症状に与える影響も無視できない。
まとめ
極端男性脳仮説は、自閉症の脳が「男性脳」の特性(システム化の優位性、共感の低下)を極端に強調した形態であると提案しています。この特性は、胎児期のテストステロンの過剰な影響や父性インプリント遺伝子の発現によって引き起こされる可能性があります。性差に基づく認知特性(女性の共感力、男性の空間能力など)を背景に、自閉症の社会的認知の障害と技術的理解の保存を説明する枠組みを提供します。ただし、仮説には限界があり、自閉症の多様性や環境的要因を完全に説明するにはさらなる研究が必要です。
極端男性脳仮説(Extreme Male Brain Theory)を支持する間接的な証拠として、以下の3つの観察を挙げています:
- 先天性副腎過形成(CAH)の女児が、影響を受けていない姉妹と比較して、より男性的な行動や自閉症様の特性を示す。
- 胎児テストステロンと**第2指と第4指の長さの比率(2D:4D比)**の逆相関、および自閉症での低い2D:4D比。
- 自閉症の男児における思春期の早熟。
以下で、これらのポイントを詳しく解説します。
1. 先天性副腎過形成(CAH)の女児と男性的行動・自閉症様特性
- 先天性副腎過形成(CAH)とは:
- CAHは、副腎がホルモンを正常に生成できない遺伝性疾患で、特に21-ヒドロキシラーゼ欠損症が一般的です。この結果、コルチゾールやアルドステロンの生成が減少し、代わりにアンドロゲン(テストステロンを含む男性ホルモン)の過剰産生が起こります。
- 女児の場合、胎児期に高レベルのアンドロゲンに曝露することで、性分化や脳の発達に影響が出ます。
- CAH女児の行動特性:
- CAHの女児は、胎児期の高アンドロゲン曝露により、典型的には女児よりも「男性的」な行動を示すことが観察されています。具体的には:
- 遊びの好み:女児らしいお人形遊びよりも、車やブロックなどの「男児らしい」玩具を好む。
- 社会的行動:攻撃性や競争心が高まる傾向。
- 空間的認知:空間的タスク(例:心的回転)でのパフォーマンスが、典型的な女児よりも優れている場合がある。
- これらの行動は、テストステロンが脳の「男性化」を促進し、極端男性脳仮説で述べられる「システム化(Systemizing)」の傾向を強めることを示唆します。
- CAHの女児は、胎児期の高アンドロゲン曝露により、典型的には女児よりも「男性的」な行動を示すことが観察されています。具体的には:
- 自閉症様特性との関連:
- CAHの女児は、影響を受けていない姉妹(対照群)と比較して、自閉症スペクトラム障害(ASD)に似た特性を示すことが報告されています。たとえば:
- 社会的コミュニケーションの困難:他者との感情的なつながりや視点取得(メンタライジング)の能力が低下する。
- 限定的な興味:特定のトピックや活動に対する強いこだわり。
- 感覚過敏:自閉症でよく見られる感覚処理の異常。
- これらの特性は、自閉症の「共感(Empathizing)の低下」と「システム化の強化」に一致し、胎児期のテストステロン過剰が自閉症様の脳発達を引き起こす可能性を支持します。
- CAHの女児は、影響を受けていない姉妹(対照群)と比較して、自閉症スペクトラム障害(ASD)に似た特性を示すことが報告されています。たとえば:
- 証拠の意義:
- CAH女児の観察は、テストステロンが脳の性分化や認知特性に直接影響を与える証拠を提供します。
- 自閉症の性比(男性に多い、約4:1)が、テストステロンの関与を反映している可能性を示唆します。
- ただし、CAH女児全員が自閉症を発症するわけではなく、あくまで「自閉症様」の特性が高まる傾向がある点に注意が必要です。
2. 胎児テストステロンと2D:4D比の逆相関
- 2D:4D比とは:
- 2D:4D比は、第2指(人差し指)の長さを第4指(薬指)の長さで割った比率です。この比率は、胎児期のテストステロンとエストロゲン(女性ホルモン)のバランスを間接的に反映するとされています。
- 低い2D:4D比(人差し指が薬指より短い)は、胎児期にテストステロンへの曝露が多かったことを示します。これは男性で一般的に見られ、女性では2D:4D比がより高い(人差し指と薬指の長さが近い)傾向があります。
2. 胎児期テストステロンと2D:4D比(続き)
2D:4D比と胎児期ホルモン: 2D:4D比(人差し指の長さと薬指の長さの比率)は、妊娠中の胎児期のテストステロンとエストロゲンのバランスを反映すると考えられているバイオマーカーです。
低い2D:4D比(薬指に比べて人差し指が短い)は、出生前テストステロンへのより高い曝露と関連しています。このパターンは男性に多く見られ、一方、女性はより高い2D:4D比(人差し指と薬指の長さがより類似している)を持つ傾向があります。
この比率は胎児の発達の初期に確立され、生涯を通じて比較的安定しているため、出生前のホルモンが脳と行動に与える影響を研究するための有用な指標となります。
自閉症における2D:4D比: 研究により、自閉スペクトラム症(ASD)の個人、特に男性は、神経典型的な個人と比較して低い2D:4D比を持つ傾向があることがわかっています。これは、彼らが子宮内でより高いレベルのテストステロンに曝露されていた可能性を示唆しています。
例えば、Manningら(2001年)などの研究では、自閉症の個人だけでなく、彼らの第一度親族(例えば、両親や兄弟姉妹)もしばしば低い2D:4D比を示すことが示されており、出生前テストステロンが自閉症のリスクに寄与する可能性を支持しています。
この発見は、自閉症が高い出生前テストステロンによって引き起こされる「男性的な」認知特性(例えば、強いシステム化、弱い共感)の誇張によって特徴づけられると仮定する極端な男性脳理論と一致しています。
重要性: 自閉症における低い2D:4D比は、胎児期のテストステロンの上昇が、自閉症の特性の可能性を高めるような方法で脳の発達を形成した可能性を示す間接的な証拠となります。
また、これは自閉症の有病率における男性の偏り(男性対女性の比率がおよそ4対1)を説明するのにも役立ちます。男性は子宮内で自然により高いレベルのテストステロンに曝露されるためです。
しかし、2D:4D比は自閉症の決定的な診断ツールではありません。個人間でばらつきがあり、他の要因(遺伝的要因、環境要因)も比率と自閉症のリスクに影響を与えるためです。
限界: 低い2D:4D比と自閉症の関連性は一部の研究で一貫していますが、普遍的ではなく、効果量も小さいことが多いです。
2D:4D比は間接的な指標であり、他の要因(例えば、遺伝、母親のストレス)が比率と自閉症のリスクの両方に影響を与える可能性があり、因果関係の解釈を複雑にしています。
3. 自閉症の男の子の早期の思春期
思春期とテストステロン: 思春期は、視床下部-下垂体-性腺軸の活性化によって引き起こされ、男性におけるテストステロンを含む性ホルモンの産生増加につながります。
早期思春期(性早熟症)とは、典型的な年齢範囲よりも前に思春期の変化が始まることを指します(例えば、男の子で9歳より前)。遺伝的要因、ホルモン要因、または環境要因によって影響を受ける可能性があります。
自閉症の男の子における証拠: 一部の研究では、自閉症の男の子は神経典型的な同年代の男の子と比較して、より早く思春期を迎える可能性があると報告されています。例えば、研究では、一部の自閉症の男性において、早期の精巣の成長や二次性徴などの性早熟の兆候が認められています。
この観察は、胎児期のテストステロン曝露に影響を与えるのと同じホルモンメカニズムに関連すると考えられています。発達期のテストステロンレベルの上昇(またはテストステロンに対する感受性の亢進)は、思春期のプロセスのより早期の活性化のために身体を「準備」する可能性があります。
例えば、Tordjmanら(1997年)の研究では、一部の自閉症の男の子が早期の思春期の変化の兆候を示しており、それは神経内分泌系の調節不全と関連している可能性が示唆されました。
極端な男性脳理論との関連: 自閉症の男の子の早期の思春期は、出生前と出生後の発達全体にわたるテストステロンの影響の亢進というより広範なパターンを反映している可能性があります。
テストステロンは、脳の組織化と行動に影響を与え、「男性脳」に関連する特性(例えば、共感よりもシステム化)を促進することが知られています。もし自閉症の男の子が高いテストステロンプロファイルを持っている場合、これはシステムへの強い関心や社会的認知の困難さなど、自閉症の認知および行動特性に寄与する可能性があります。
思春期の早期の発症は、青年期のホルモンの変化が社会的行動、感情調節、および感覚過敏に影響を与える可能性があるため、特定の自閉症の特性を悪化させる可能性もあります。
重要性: 自閉症の男の子における早期の思春期の証拠は、テストステロンが自閉症の発達の軌跡を形成する上で役割を果たしているという考えを支持しています。
これは、出生前のホルモン影響(例えば、高い胎児期テストステロン)とその後の発達の結果との関連性を示し、極端な男性脳理論を強化します。
この観察は、早期の思春期が場合によっては医学的なモニタリングや介入を必要とする可能性があるため、臨床的な意味合いも持ちます。
限界: すべての自閉症の男の子が早期の思春期を経験するわけではなく、証拠は再現が必要な小規模な研究に基づいています。
思春期のタイミングは複数の要因(例えば、遺伝、栄養、ストレス)によって影響を受けるため、テストステロンだけでは観察されたパターンを完全に説明できない可能性があります。
早期の思春期と自閉症特有の特性(例えば、社会的な困難さ)との関連性は完全には確立されておらず、さらなる研究が必要です。
全体的な考察とこれらの観察の全体的な重要性
先天性副腎過形成症(CAH)の女の子: CAH(出生前のアンドロゲンへの曝露が多い)の女の子が、より男性的な行動(例えば、「男の子らしい」おもちゃの好み、より高い攻撃性)や自閉症に似た特性(例えば、社会性の低下、限定された興味)を示すという観察は、出生前のテストステロンが脳の発達と行動を男性化する可能性があることを示唆しています。
これは自閉症の認知プロファイル(強いシステム化、弱い共感)と一致しており、高いテストステロンが自閉症の特性に寄与する可能性があるという考えを支持しています。
2D:4D比: 自閉症の個人における低い2D:4D比は、より高い出生前テストステロン曝露を示しており、自閉症の男性に偏った有病率と、自閉症が男性脳の組織の極端なバージョンを反映しているという理論と一致しています。
早期の思春期: 自閉症の男の子の早期の思春期は、青年期へのテストステロンの影響の亢進の継続を示唆しており、極端な男性脳理論に関連する特性を潜在的に増幅させています。
仮説への総合的な支持: これらの発見は、上昇したテストステロン(出生前および出生後)を自閉症に特徴的な行動や特性と結びつけることで、極端な男性脳理論の間接的な証拠を総合的に提供します。
これらは、テストステロンが脳の発達の「彫刻家」として作用し、システム化能力(例えば、パターン認識、機械的理解)を高めながら、共感能力(例えば、社会的認知、感情理解)を低下させる可能性を示唆しています。
自閉症の有病率における男性の偏り、CAHの女の子における男性化された行動、低い2D:4D比、そして早期の思春期はすべて、自閉症において男性的な脳の特性を誇張する可能性のあるホルモンメカニズムを示唆しています。
より広範な意味合い: これらの観察は、自閉症のような神経発達障害におけるホルモン影響の重要性を強調しています。
これらは、ホルモン経路の調節が自閉症の特性に影響を与えるかどうかを調査するなど、研究の潜在的な方向性を示唆しています(ただし、これは推測的であり、倫理的に複雑です)。
臨床的には、これらのパターンを理解することで、自閉症の男の子の思春期の発達のモニタリングや、CAHの影響を受けた女の子の社会的な課題への対処など、個別化された介入に役立つ可能性があります。
限界と今後の方向性
証拠の間接性: これら3つの観察(CAH、2D:4D比、早期の思春期)はすべて相関関係であり、因果関係ではありません。これらはテストステロンの役割を示唆していますが、他の要因(遺伝的要因、環境要因)も自閉症に寄与しています。
2D:4D比と思春期のタイミングは複数の変数によって影響を受けるため、テストステロンの特定の役割を分離することは困難です。
自閉症の異質性: 自閉症は非常に多様であり、すべての個人が「極端な男性脳」のプロファイルに当てはまるわけではありません。例えば、自閉症の女性は異なる認知および行動パターンを示す可能性があり、この理論はそれを完全には扱っていません。
一部の自閉症の個人は共感課題に優れていたり、システム化に特別な強みを示さなかったりすることがあり、よりニュアンスのある理解の必要性を強調しています。
さらなる研究の必要性: 自閉症の男の子における早期の思春期や、多様な集団における2D:4D比の一貫性などの発見を確認するためには、より大規模な縦断的研究が必要です。
出生前テストステロンの直接的な測定(例えば、羊水による)と、その後の自閉症の特性との相関関係は、証拠を強化する可能性があります。
テストステロンと遺伝的要因(例えば、父性刷り込み遺伝子)の相互作用を調査することで、自閉症におけるそれらの複合的な役割を明らかにすることができます。
まとめ
極端な男性脳理論は、自閉症が高い出生前テストステロンによって引き起こされる男性的な脳の特性の誇張を反映していると仮定しています。支持する証拠には以下が含まれます。
- CAHの女の子が男性化された行動と自閉症に似た特性を示し、脳の男性化におけるテストステロンの役割を示唆している。
- 自閉症の個人における低い2D:4D比は、より高い出生前テストステロン曝露を示している。
- 自閉症の男の子の早期の思春期は、自閉症の特性を増幅させる可能性のある持続的なテストステロンの影響を示唆している。
これらの発見は、間接的ではあるものの、説得力のある理論の裏付けとなり、テストステロンを自閉症の認知および行動プロファイルに関連付けています。ただし、証拠は相関関係であり、自閉症の複雑さを完全に理解するためには、これらのメカニズムに関するさらなる研究が必要です。
この段落では、正常発達の子どもにおける胎児期テストステロンの影響と、自閉症の脳に見られる可能性のある特徴、そしてそれらの関連性について議論しています。
正常発達における胎児期テストステロンの影響
冒頭の部分では、正常に発達している子どもにおいて、胎児期のテストステロンレベルが高いほど、以下の能力や行動が低い傾向にあると指摘しています。
- 目と目を合わせる接触: 他者と視線を合わせる行動は、社会的なコミュニケーションや感情の共有において非常に重要です。胎児期テストステロンが高いと、この行動が少なくなる可能性があるとされています。
- 共同注意: 他者と関心のある対象を共有する能力です。例えば、誰かが指さしたものを一緒に見たり、同じ遊びに一緒に参加したりする行動を指します。これも社会的な相互作用の基礎となる重要な能力であり、胎児期テストステロンが高いと発達が遅れる可能性があるとされています。
- 言語発達: コミュニケーションの主要な手段である言語の発達も、胎児期テストステロンの影響を受ける可能性が示唆されています。
- 社会的機能: 他者との関係を築き、維持する能力、社会的なルールや規範を理解し、適切に行動する能力など、社会生活を送る上で必要な様々な機能です。胎児期テストステロンが高いと、これらの機能の発達が遅れる可能性があるとされています。
- 限定された興味の範囲: 特定の対象や活動に非常に強い関心を持ち、他のことへの関心が狭くなる傾向です。これは自閉症の主要な特徴の一つですが、正常発達の子どもにおいても、胎児期テストステロンが高いと、このような傾向が見られる可能性があるとされています。
自閉症と胎児期テストステロン、アンドロゲン受容体
次に、自閉症が胎児期テストステロンレベルの上昇や、性的二形性を示す脳領域(男女間で構造や機能に差がある領域)におけるアンドロゲン受容体(テストステロンなどのアンドロゲンを受け取るタンパク質)の数の増加と関連しているかどうかは不明であると述べています。これは、現時点ではまだ明確な結論が出ていない研究課題であることを示しています。もし関連性があれば、胎児期の高いテストステロン曝露が、後の自閉症の発症や特性の発現に影響を与えるメカニズムの一端を説明できる可能性があります。
自閉症の脳の解剖学的特徴
続いて、自閉症の脳に見られる可能性のある解剖学的データがいくつか挙げられています。
- 脳の肥大: 全体的な脳の体積が出生時から増加している傾向があることが示唆されています。
- 出生時の扁桃体のサイズの増大(思春期後に相対的なサイズが正常以下の体積に低下): 扁桃体は感情処理に関わる脳領域です。出生時には大きいものの、思春期以降には相対的に小さくなるという、特異な発達パターンが示唆されています。
- 神経細胞密度の増加(ただし連結が不十分): 脳の神経細胞の数は多いものの、それらの間の連結(シナプス)が十分に発達していない可能性があるとされています。これは、情報処理の効率性や統合性に影響を与える可能性があります。
- 生後数年間の白質量の増加: 白質は神経細胞の軸索が集まった部分で、脳の異なる領域間の情報伝達を担います。自閉症では、生後数年間でこの白質量が増加する傾向があることが示唆されています。
自閉症における脳の連結
最後に、自閉症における脳の連結に関するデータについて触れています。
- 半球間の連結の減少: 左右の大脳半球を結ぶ神経線維の束(脳梁など)の機能や構造に異常があり、左右の脳の情報伝達がうまくいかない可能性が示唆されています。
- 半球内の連結の減少を示す知見と矛盾: 一方で、同じ半球内の異なる領域間の連結が減少しているという知見もあり、これらは互いに矛盾していると指摘されています。脳の連結パターンは複雑であり、自閉症においても一様ではない可能性を示唆しています。
半球間の連結の減少と「中心的統合性」の欠如
しかし、半球間の連結の減少は、「中心的統合性」の欠如、つまり個々の部分よりも全体像(ゲシュタルト)を認識する能力のような認知プロセスの機能不全を説明できるかもしれないと述べています。これは、自閉症の人が細部に注意を払いやすい一方で、全体的な状況や文脈を把握することが苦手な傾向があることと関連付けられる可能性があります。左右の脳が連携して情報を統合する能力の低下が、このような認知特性の一因となっている可能性が考えられます。
まとめ
この部分は、胎児期テストステロンが正常発達における社会性やコミュニケーション能力の発達に負の影響を与える可能性を示唆するとともに、自閉症の脳の構造や連結の異常について概説しています。特に、脳の肥大、扁桃体の発達の特異性、神経細胞密度と連結の不均衡、白質量の増加、そして半球間の連結の減少などが指摘されています。そして、この半球間の連結の減少が、自閉症に見られる「中心的統合性」の欠如という認知特性を説明する可能性についても言及しています。ただし、自閉症と胎児期テストステロンやアンドロゲン受容体の関連性については、まだ明確な結論が出ていないことが強調されています。
この段落では、前述の「中心的統合性」の欠如という考え方をさらに発展させ、自閉症のサヴァン症候群の認知スタイルや、自閉症の人の言語理解における文字通りの解釈と関連付け、さらに性差との関連性を示唆しています。
自閉症のサヴァンと文字通りの世界の見方
冒頭で、「この知見と一致して、自閉症のサヴァンは世界を文字通りに見ていると提案されています」と述べています。ここでいう「この知見」とは、前の段落で説明された「中心的統合性」の欠如、つまり全体像(ゲシュタルト)を捉えるのが苦手で、細部に注意を払いやすいという自閉症の認知特性を指しています。
サヴァン症候群とは、自閉症などの発達障害を持つ人の中で、特定の分野において非常に 뛰어난、常識では考えられないほどの能力を発揮する状態を指します。例えば、一度見た風景を細部まで正確に記憶する、複雑な計算を一瞬で行う、聞いた音楽を完璧に再現するなどです。
この段落では、自閉症のサヴァンが驚異的な能力を発揮する背景には、「概念や心的テンプレートの正常な獲得を欠いている」可能性があると指摘しています。「概念」とは、複数の具体的な事物や事柄に共通する性質を抽象化したものです。「心的テンプレート」とは、私たちが経験を整理し、予測するために脳内に持っている、ある種の枠組みやパターン認識のシステムのようなものです。
もし、これらの概念や心的テンプレートの獲得がうまくいかない場合、世界を経験する際に、既存の枠組みに当てはめて全体像を理解することが難しくなります。その結果、彼らは「ゲシュタルトではなく詳細の知覚に依存している」と考えられます。つまり、物事を全体として捉えるのではなく、個々の具体的な細部に注意を集中して世界を認識しているということです。サヴァンの驚異的な記憶力や細部への注意深さは、このような認知スタイルによって説明される可能性があります。
自閉症の人の話し言葉の文字通りの解釈
次に、「これはまた、自閉症の人の話し言葉の解釈における文字通りさを部分的に説明することもできます」と述べています。自閉症の人は、比喩表現、冗談、皮肉など、言葉の文字通りの意味とは異なる意図が含まれている場合、その解釈に困難を抱えることがあります。
「中心的統合性」の欠如によって全体的な文脈や意図を捉えることが難しい場合、言葉を個々の要素として、文字通りの意味で解釈する傾向が強くなるのは自然な流れと言えます。例えば、「それは君のボールパークだ」という比喩表現を、文字通り「それは君の野球場だ」と解釈してしまう可能性があります。
正常な男性と女性の間の違い
さらに、「このような違いは、正常な男性と女性の間にも存在します(ただし、はるかに程度が低い)」と指摘しています。これは、男性は女性と比較して、細部に注意を払いやすく、全体的な文脈よりも具体的な情報に焦点を当てる傾向があるという、心理学的な研究における一般的な知見と一致しています。また、言語理解においても、男性の方が女性よりもやや文字通りの解釈をする傾向があるという研究もあります。
ただし、重要なのは、この性差は自閉症に見られる特徴と比較して「はるかに程度が低い」ということです。自閉症の人は、健常な男性よりもさらに強く、この文字通りの解釈や細部への注目の傾向を示すと考えられます。
男性的特性と女性的特性の差異的な遺伝や発現と自閉症の病因
最後に、「したがって、男性的特性と女性的特性の差異的な遺伝や発現が自閉症の病因に関与していると考えられます」と結論付けています。
これまでの議論を踏まえると、自閉症に見られる認知スタイル(細部への注目、全体像の把握の困難さ、文字通りの解釈など)は、一般的に男性に多く見られる特性が極端な形で現れたものと捉えることができます。
したがって、自閉症の発症には、男性的な特性に関連する遺伝子やその発現の仕方の異常、あるいは女性的な特性に関連する遺伝子やその発現の抑制などが関与している可能性があるという考え方が提示されています。これは、極端な男性脳理論の根幹となる考え方であり、自閉症の病因を理解する上で、遺伝子レベルでの性差に着目することの重要性を示唆しています。
まとめ
この部分は、自閉症の中核的な認知特性である「中心的統合性」の欠如が、サヴァンの特異な能力や言語理解における文字通りの解釈とどのように関連しているかを説明しています。さらに、これらの特性が正常な男性にも程度は低いながらも見られることを指摘し、自閉症の病因には、男性的な特性と女性的な特性の差異的な遺伝や発現が関与している可能性を示唆しています。これは、自閉症を「極端な男性脳」として捉える理論を支持する重要な議論の一つと言えるでしょう。
この段落では、進化的包括適応度理論とゲノムインプリンティングの概念を紹介し、親から受け継いだ遺伝子間に潜在的な「ゲノム内の葛藤」が存在する可能性と、それが子孫における遺伝子発現にどのように影響するかについて説明しています。
進化的包括適応度理論
まず、「進化的包括適応度理論」についてです。この理論は、個体の適応度(繁殖成功度)だけでなく、その個体の遺伝子を共有する親族(例えば、兄弟姉妹、甥姪など)の繁殖成功度も考慮に入れることで、利他的な行動などがどのように進化してきたかを説明しようとするものです。
この理論の重要な側面の一つは、遺伝子は自身のコピーを次世代に伝えることを「目的」としていると考える視点です。したがって、同じ個体内に存在する遺伝子であっても、その起源(母親由来か父親由来か)によって、次世代への伝わり方を最大化するための「戦略」が異なる可能性があります。
ゲノム内の葛藤(ゲノムインプリンティングの文脈で)
次に、「母方と父方から受け継がれた遺伝子の間でゲノム内の葛藤が生じる可能性がある」という点です。これは、特に母親が複数の子どもを産むことができる哺乳類において顕著になると考えられています。
- 父親由来の遺伝子の「利己性」: 父親にとって、自分の遺伝子を次世代に伝える最も直接的な方法は、自分の子どもが生存し、繁殖することです。父親は、母親が他の男性との間に別の子どもを持つ可能性があるため、現在の子どもにできるだけ多くの資源(母親の栄養など)を分配し、その生存と成長を最大化しようとする遺伝子を進化させる可能性があります。これは、現在の子どもの生存を高める一方で、母親の将来の繁殖能力を低下させる可能性も孕んでいます。
- 母親由来の遺伝子の「利他的性」: 一方、母親は自身の子どもたち全員に遺伝子を伝えます。したがって、現在の子どもに過剰な資源を分配することで自身の将来の繁殖能力が低下したり、他の子どもへの資源が不足したりすることは、母親の包括適応度にとっては不利になる可能性があります。そのため、母親由来の遺伝子は、資源の分配をより公平に、あるいは母親自身の生存と将来の繁殖を優先するように働く可能性があります。
このように、父親由来の遺伝子は現在の子どもの利益を最大化しようとする傾向があるのに対し、母親由来の遺伝子はより長期的な視点や、すべての子どもの利益を考慮する傾向があるため、遺伝子レベルでの「葛藤」が生じる可能性があると考えられています。
ゲノムインプリンティング
ここで重要なのが「ゲノムインプリンティング」という現象です。これは、親から子へ遺伝子が伝わる際に、特定の遺伝子について、父親由来のコピーか母親由来のコピーかによって、その活性が異なる(一方のコピーが沈黙させられる)現象です。
「ゲノムインプリンティングの場合のように、もう一方の親からの遺伝子が沈黙させられると、父方または母方由来の遺伝子が子孫で過剰に発現される可能性があります」と述べられています。
もし、ある遺伝子が父親由来のコピーのみが活性を持ち、母親由来のコピーが沈黙させられている場合(父性刷り込み)、その遺伝子の効果は子孫において通常よりも強く現れる可能性があります。これは、母親由来の抑制的な効果が失われているためです。同様に、母親由来のコピーのみが活性を持つ場合(母性刷り込み)、その遺伝子の効果が過剰に現れる可能性があります。
自閉症との関連性(文脈から推測)
この段落は直接的に自閉症について述べていませんが、前の議論の流れから考えると、ゲノムインプリンティングの異常が、父方または母方由来の特定の遺伝子の過剰発現を引き起こし、それが脳の発達や機能に影響を与え、自閉症の病因に関与している可能性を示唆していると考えられます。
例えば、父性刷り込み遺伝子の過剰発現が、父親由来の「より多くを求める」傾向を増幅させ、それが社会的な相互作用やコミュニケーションの発達に影響を与える、といったシナリオが考えられます。
まとめ
この部分は、進化的包括適応度理論に基づき、親由来の遺伝子間に資源分配に関する潜在的な葛藤が存在すること、そしてゲノムインプリンティングという現象によって、一方の親由来の遺伝子が沈黙することで、もう一方の親由来の遺伝子が子孫で過剰に発現する可能性があることを説明しています。これは、遺伝子レベルでの親の起源が、子どもの発達に異なる影響を与える可能性を示唆しており、自閉症の病因を遺伝子レベルで理解する上で重要な視点となります。
はい、承知いたしました。この部分では、ゲノムインプリンティングが自閉症の病因に関与している可能性について、いくつかの観察事実とアンジェルマン症候群との関連性を示しながら、より具体的な議論を展開しています。
ゲノムインプリンティングと自閉症の関連性の推測
冒頭で、「この段階では、ゲノムインプリンティングが自閉症において主要な役割を果たしているというのは推測的です」と注意を促しています。これは、ゲノムインプリンティングと自閉症の直接的な因果関係が完全に解明されているわけではないことを示唆しています。しかし、以下の観察事実は、父性インプリンティングが自閉症スペクトラム障害(ASD)の少なくとも一部に関与しているという仮説を強く支持すると述べています。
父性インプリンティングを示唆する観察事実
- 行動的および解剖学的な男性的特性の過剰発現: 前の議論で触れられたように、自閉症は「極端な男性脳」として捉えられることがあります。もし父性インプリンティングが特定の遺伝子の過剰発現を引き起こし、それが男性的な特性の発達を促進するならば、自閉症に見られる男性的特性の偏りは、父性インプリンティングの異常と関連付けられる可能性があります。
- 胎盤のサイズの増大(増殖性成長の兆候を伴う): 胎盤は胎児に栄養や酸素を供給する重要な器官です。父性由来の遺伝子は、胎児の成長を促進するために、胎盤からの資源供給を最大化する傾向があると進化的に考えられています。もし父性インプリンティングの異常により、胎盤の成長を促進する遺伝子が過剰に発現すると、胎盤が異常に大きくなる可能性があります。これは、父性遺伝子の影響が強まっている兆候と解釈できます。
- 自閉症の子どもの出生時体重の増加: 同様に、父性由来の遺伝子は胎児の成長を促進する傾向があるため、父性インプリンティングの異常により成長促進遺伝子が過剰に発現すると、出生時の体重が増加する可能性があります。これも、父性遺伝子の影響が強まっている可能性を示唆する所見です。
これらの観察事実は、直接的な証拠ではないものの、父性由来の遺伝子の影響が自閉症の特徴と関連している可能性を示唆しており、父性インプリンティングが自閉症の病因の一端を担っているという仮説を支持する根拠となります。
アンジェルマン症候群との関連性
次に、染色体15上の父性遺伝子の過剰発現によって引き起こされる疾患であるアンジェルマン症候群との関連性が指摘されています。(第1章でこの症候群について触れられていることが示されています。)
アンジェルマン症候群は、以下の自閉症に類似した症状を高頻度で示すことが知られています。
- 言語獲得障害: 言葉の発達が遅れる、またはほとんど言葉を話さない。
- 視線回避: 他者の目を見ることを避ける傾向がある。
- 常同行動: 同じ行動を繰り返す(例:手をひらひらさせる、体を揺らすなど)。
アンジェルマン症候群は、通常、母親由来の染色体15q11-q13領域にある UBE3A 遺伝子の機能喪失によって引き起こされます。しかし、この段落では、父性遺伝子の過剰発現がアンジェルマン症候群を引き起こす状況にも言及している可能性があります(例えば、父性ダイソミーと呼ばれる、父親から2コピーの染色体15を受け継ぎ、母親から受け継がない場合など)。
いずれにせよ、アンジェルマン症候群において、父性遺伝子に偏った遺伝子発現の不均衡が見られ、それが自閉症と類似した症状を引き起こすという事実は、「父性遺伝に向けた遺伝子発現の不均衡のさらなる証拠」と解釈できると述べています。これは、遺伝子発現のバランスの崩れ、特に父性遺伝子の過剰な影響が、自閉症スペクトラムの症状発現に関与している可能性を強く示唆しています。
まとめ
この部分は、ゲノムインプリンティング、特に父性インプリンティングの異常が、自閉症の病因に関与している可能性について、具体的な観察事実とアンジェルマン症候群との関連性を用いて議論を展開しています。行動的・解剖学的な男性的特性の過剰発現、胎盤のサイズの増大、出生時体重の増加といった所見が父性インプリンティングの関与を示唆する一方、父性遺伝子の過剰発現が自閉症類似の症状を引き起こすアンジェルマン症候群の存在は、遺伝子発現の不均衡が自閉症スペクトラム障害の一因となる可能性を強く示唆しています。ただし、この時点ではまだ推測の域を出ないことも強調されています。
はい、承知いたしました。この部分では、自閉症におけるゲノムインプリンティングに関する別の視点として、ターナー症候群の研究から得られた知見を紹介し、X染色体の起源が社会認知能力や実行機能に与える影響、そしてそれが自閉症との関連を示唆する可能性について説明しています。
ターナー症候群とX染色体の起源
ターナー症候群は、女性においてX染色体が部分的または完全に欠失している染色体異常です。通常の女性は2本のX染色体(XX)を持ちますが、ターナー症候群の女性は1本のX染色体(X0)のみを持ちます。
この段落で重要なのは、その1本のX染色体が母親由来であるか父親由来であるかによって、ターナー症候群の女性の認知機能に違いが見られるという点です。
- 母親由来のX染色体を持つ場合(約70%のケース): この場合、社会的認知障害や自閉症様の特性が、父親由来のX染色体を持つターナー症候群の女性と比較して、より一般的に見られるとされています。社会的認知とは、他者の感情や意図を理解する能力、社会的な状況を適切に判断する能力などを指します。
- 父親由来のX染色体を持つ場合: こちらの場合、母親由来のX染色体を持つ女性と比較して、社会的認知障害や自閉症様の特性の頻度が低いとされています。
ゲノムインプリンティングとの関連性
この観察は、X染色体上にはゲノムインプリンティングを受ける遺伝子が存在する可能性を示唆しています。つまり、同じ遺伝子であっても、母親から受け継がれたか父親から受け継がれたかによって、その発現や機能が異なるということです。
もし、社会認知能力の発達に関わるX染色体上の遺伝子に母性刷り込み(母親由来の遺伝子の発現が抑制される)が存在する場合、母親由来のX染色体しか持たないターナー症候群の女性は、その遺伝子の機能が十分に発揮されず、社会認知障害のリスクが高まる可能性があります。
逆に、もし社会認知能力に関わる別のX染色体上の遺伝子に父性刷り込み(父親由来の遺伝子の発現が抑制される)が存在する場合、父親由来のX染色体しか持たないターナー症候群の女性は、その遺伝子の機能が十分に発揮され、社会認知障害のリスクが低くなる可能性があります。
自閉症との関連性
このターナー症候群の研究結果は、自閉症におけるゲノムインプリンティングの役割について、これまでとはやや異なる視点を提供します。
自閉症は男性に多いという性差が見られますが、女性の自閉症者も存在します。ターナー症候群の研究は、X染色体上の遺伝子の発現の違いが、社会認知能力の発達に影響を与える可能性を示唆しており、これが自閉症の性差や、女性の自閉症の特性の現れ方の違いを理解する上で手がかりとなるかもしれません。
例えば、特定のX染色体上の遺伝子の母性刷り込みが、女性において社会認知能力の発達を阻害しやすく、それが他の遺伝的要因や環境要因と組み合わさることで、自閉症の発症リスクを高める可能性がある、といった仮説が考えられます。
まとめ
この部分は、ターナー症候群の研究から、X染色体の起源が社会認知能力や実行機能に影響を与えるという興味深い知見を紹介しています。これは、X染色体上にもゲノムインプリンティングを受ける遺伝子が存在する可能性を示唆しており、特定の遺伝子の母性由来または父性由来の活性の違いが、社会認知の発達、ひいては自閉症の感受性に影響を与える可能性を示唆しています。自閉症の性差や、女性の自閉症の特性を理解するための新たな研究の方向性を示唆する知見と言えるでしょう。
はい、承知いたしました。この部分は、前述のターナー症候群の研究結果に基づいて、社会的認知に関与するX染色体上の特定の遺伝子座のインプリンティングの可能性を提唱し、それが男女間の社会的認知能力の差、男性の自閉症への脆弱性、そしてターナー症候群における自閉症症状の現れ方の違いを説明できるかもしれないと論じています。
社会的認知に関与するX染色体上の遺伝子座の父性発現
この段落は、社会的認知に関与するX染色体上の特定の遺伝子座について、以下のような仮説を立てています。
- 正常な女性(46, XX)の場合: 父親から受け継いだX染色体上のその遺伝子座のみが発現し、母親から受け継いだ同じ遺伝子座は沈黙している(父性刷り込み)。
- 正常な男性(46, XY)の場合: 母親から受け継いだX染色体上のその遺伝子座が発現する。
女性の社会的認知の優位性の説明
この仮説は、なぜ女性が一般的に男性と比較して社会的認知が優れている可能性があるかを説明できるとしています。女性は父親由来のX染色体から活性のある社会的認知関連遺伝子を受け継ぎますが、母親由来のX染色体上の同じ遺伝子は沈黙しています。一方、男性は常に母親からX染色体を受け継ぎ、もしその遺伝子座が父性刷り込みを受けるならば、男性の持つその遺伝子は活性化されない可能性があります。これにより、女性の方が社会的認知に関連する遺伝子を活性化された状態で持つ可能性が高くなり、それが社会的認知能力の優位性につながるかもしれません。
男性の自閉症への脆弱性の説明
次に、なぜ男性が自閉症スペクトラム障害を含む発達障害に対してはるかに脆弱であるかを説明できるとしています。もし社会的認知に関わる重要な遺伝子がX染色体上に存在し、父性刷り込みを受ける場合、男性は母親から受け継いだその遺伝子を唯一のX染色体として持つことになります。もしその遺伝子の発現量が不十分であったり、機能に何らかの変異があったりした場合、男性はそれを補うためのもう一つのX染色体を持たないため、発達障害のリスクが高まる可能性があります。一方、女性はもう一つのX染色体を持っているため、片方のX染色体に問題があっても、もう片方のX染色体がその機能を補完できる可能性があります。これは、X染色体に関連する遺伝子の異常が男性に顕著に現れやすいという「X連鎖遺伝」の考え方とも一部関連します。
ターナー症候群における自閉症症状の違いの説明
最後に、ターナー症候群(45, X0)の個人が、父親のX染色体を持つX0女性と比較して母親由来のX染色体を持つ場合に、より多くの自閉症症状を示す理由についても説明できるとしています。
- 母親由来のX染色体を持つX0女性: この場合、活性化されない(父性刷り込みを受けている)可能性のある社会的認知関連遺伝子しか持たないため、社会的認知障害や自閉症様の症状が現れやすくなる可能性があります。
- 父親由来のX染色体を持つX0女性: この場合、活性化される社会的認知関連遺伝子を持つため、母親由来のX染色体を持つ女性と比較して、社会的認知能力が比較的保たれ、自閉症症状も少ない可能性があります。
まとめ
この部分は、社会的認知に関わるX染色体上の特定の遺伝子座に父性刷り込みが存在するという仮説を提唱し、それが男女間の社会的認知能力の差、男性の自閉症への脆弱性、そしてターナー症候群における自閉症症状の現れ方の違いを統一的に説明できる可能性を示唆しています。この仮説は、ゲノムインプリンティングが神経発達障害の性差や症状の多様性を理解する上で重要な役割を果たしている可能性を強調するものです。
はい、承知いたしました。この部分では、前述の父性インプリンティングと母性インプリンティングに関する二つの仮説について、その現状の支持状況と限界を述べ、自閉症スペクトラムの多様性を説明するための今後の研究の方向性を示唆しています。
両インプリンティング仮説の現状と限界
冒頭で、「両方のインプリンティング仮説は部分的な実証的支持を受けていますが、おそらくどちらも自閉症スペクトラムのすべての特徴を説明するには不十分です」と述べています。これは、前述の父性インプリンティング(胎盤や出生時体重の増加、アンジェルマン症候群との関連性など)と母性インプリンティング(ターナー症候群におけるX染色体の起源と社会的認知の関連性など)のどちらの仮説も、自閉症のいくつかの側面を説明する可能性はあるものの、自閉症の複雑で多様な症状全体を完全に解明するには至っていないという認識を示しています。
自閉症スペクトラム障害(ASD)は、社会的なコミュニケーションや相互作用の困難さ、限定された興味や反復的な行動を主な特徴とする、非常に多様な神経発達障害群です。個々の自閉症者は、症状の現れ方や重症度、合併する知的障害やその他の発達障害の有無など、様々な点で異なります。単一の遺伝的または発生学的メカニズムですべての自閉症の特徴を説明することは難しいと考えられています。
感受性遺伝子座との相互作用
次に、「母親または父親の染色体から発現されるインプリント遺伝子は、実際の表現型を生成するためにゲノムの他の場所にある感受性遺伝子座と相互作用する可能性が高いです」と述べています。
- インプリント遺伝子: 親由来によって発現が異なる遺伝子(父性刷り込みまたは母性刷り込みを受ける遺伝子)。
- 感受性遺伝子座: 特定の疾患や特性に対する感受性を高める遺伝子の存在する場所(ローカス)。
この考え方は、自閉症のような複雑な疾患の発症には、単一の遺伝子の異常だけでなく、複数の遺伝子の相互作用が関与する可能性が高いという「多因子遺伝」のモデルに基づいています。インプリント遺伝子の発現異常が、他の感受性遺伝子座の変異や機能異常と組み合わさることで、自閉症の様々な表現型(症状の現れ方)が生じると考えられます。例えば、あるインプリント遺伝子の異常が、特定の脳領域の発達に影響を与え、それが他の感受性遺伝子の影響を受けやすい状態を作り出す、といった相互作用が考えられます。
自閉症スペクトラムの異なるサブタイプへの適用可能性
そして、「したがって、両方のインプリンティングシナリオが自閉症スペクトラムの異なるサブタイプに適用される可能性があると考えられ、この推測は将来の研究の対象となるかもしれません」と結論付けています。
この部分は、自閉症の多様性を説明するための重要な視点を示唆しています。つまり、父性インプリンティングの異常が強く関与する自閉症のサブタイプと、母性インプリンティングの異常がより強く関与する別のサブタイプが存在する可能性があるということです。
例えば、胎盤や出生時体重の異常が見られる自閉症のサブグループでは父性インプリンティングの異常がより重要である一方、社会的認知の障害がより顕著な自閉症のサブグループでは母性インプリンティング(X染色体上の遺伝子を含む)の異常がより重要である、といった可能性が考えられます。
このような考え方は、自閉症を単一の疾患として捉えるのではなく、その多様性を考慮し、異なる病因メカニズムによって引き起こされる複数のサブタイプが存在する可能性を探求する、個別化医療への道を開くかもしれません。
将来の研究の方向性
この推測は、「将来の研究の対象となるかもしれません」と述べられており、今後の研究において、自閉症の異なる臨床的特徴を持つグループにおいて、父性および母性インプリンティングに関わる遺伝子の異常の頻度や種類を詳細に調査することが重要であることを示唆しています。ゲノムワイドな関連解析(GWAS)や、特定の候補遺伝子の詳細な機能解析、そして臨床情報との統合などが、この方向性の研究を進める上で役立つと考えられます。
まとめ
この部分は、現在のゲノムインプリンティングに関する二つの主要な仮説が、自閉症の複雑な全体像を説明するには不十分である可能性を指摘しつつ、インプリント遺伝子と他の感受性遺伝子座との相互作用の重要性を強調しています。そして、自閉症スペクトラムの多様性を理解するために、父性および母性インプリンティングの異常が、自閉症の異なるサブタイプにそれぞれ異なる形で関与している可能性を探求することが、今後の重要な研究課題であると結論付けています。
はい、承知いたしました。この部分では、自閉症に対する非薬物的介入戦略、コミュニケーション上の配慮、心理療法、薬物療法について詳しく説明しています。
非薬物的介入戦略
冒頭で、社会的・コミュニケーション能力のトレーニングや職業療法が重要な非薬物的介入戦略であると述べています。
- 社会的・コミュニケーション能力のトレーニング: これは、自閉症の人が他者との効果的なコミュニケーションや社会的な相互作用を学ぶための様々な技法やプログラムを指します。例えば、視覚的な支援(絵カードやスケジュール表)、ソーシャルストーリー(特定の社会的な状況を説明する物語)、ロールプレイングなどが用いられます。
- 職業療法: これは、日常生活を送る上で必要な様々なスキル(食事、着替え、入浴などの身辺自立、学校や職場での作業遂行能力など)を向上させるための療法です。感覚処理の問題や運動協調性の困難さなど、個々のニーズに合わせたプログラムが組まれます。
しかし、重要な注意点として、「自閉症の人々を無理に社会的交流に強制することは有用ではないことが証明されています」と強調しています。これは、無理強いがストレス反応や不安を高め、かえって社会的な引きこもりを悪化させる可能性があるためです。
したがって、「過剰な刺激を避けることは、特に持続的な注意の問題を考慮すると、自閉症の子どもたちに適切かもしれません」と述べています。自閉症の人は、感覚過敏を持つことが多く、騒がしい場所や多くの人がいる環境など、過剰な刺激に対して強い不快感や苦痛を感じることがあります。また、注意を持続させることも苦手な場合があるため、刺激の多い環境は集中をさらに困難にする可能性があります。そのため、落ち着いた環境を整え、刺激の量を調整することが、自閉症の子どもたちへの支援において重要となります。
コミュニケーション上の配慮
次に、自閉症の人々のコミュニケーションにおける困難さを考慮した配慮について述べています。
- 二重の意味を持つ会話を避ける: 比喩、冗談、皮肉など、言葉の文字通りの意味と異なる意味合いを持つ表現は、自閉症の人にとって理解が難しい場合があります。そのため、明確で直接的な言葉遣いを心がけることが推奨されます。
- 他の人々は自分とは異なる期待、知識、意図を持っていることを慎重に教育する: 自閉症の人は、他者の心の状態(考え、感情、意図など)を推測する「心の理論」の獲得に困難を抱えることがあります。そのため、他者が自分とは異なる視点を持っていることを具体的に教えることが、社会的な状況を理解し、適切な行動をとる上で役立ちます。例えば、「〇〇さんは、あなたがそれを触るのが好きではないかもしれません」のように、他者の感情や意図を明確に伝えることが重要です。
家族への介入とサポート
「家族への介入とサポートも、評価と治療手順の不可欠な部分であるべきです」と強調しています。自閉症の子どもを持つ家族は、診断、療育、日常生活の様々な課題に直面するため、心理的なサポートや情報提供、具体的な支援方法の指導などが不可欠となります。家族全体のwell-beingが、自閉症の人の発達にも大きな影響を与えるため、家族支援は治療の重要な側面です。
自閉症の心理療法
「自閉症の心理療法は、高機能自閉症やアスペルガー症候群における抑うつや不安などの併存障害を除いて、成功していません」と述べています。これは、自閉症そのものの特性(社会性の困難さ、コミュニケーションの特性など)に対する従来の心理療法のアプローチが、必ずしも有効ではないことを示唆しています。ただし、高機能自閉症やアスペルガー症候群の人々が併発しやすい抑うつや不安といった精神的な問題に対しては、認知行動療法(CBT)などの心理療法が有効な場合があります。
薬物治療
「薬物治療は一般的に推奨されておらず、深刻な行動上の問題に限られています」と述べています。自閉症の中核的な症状(社会性の困難さ、コミュニケーションの特性、常同行動)に対する直接的な薬物療法は存在しません。薬物療法は、自傷行為、激しい癇癪、重度の攻撃性など、日常生活に著しい支障をきたす行動問題に対して、他の介入が無効な場合に、慎重に検討されるべきものです。
ただし、いくつかの薬物が特定の症状の緩和に役立つ可能性が示唆されています。
- セロトニン再取り込み阻害薬(SSRI): 常同行動や自傷行為を減らす効果がある可能性を示す証拠があります。セロトニンは、気分や行動の調節に関わる神経伝達物質であり、SSRIはそのセロトニンの脳内濃度を高めることで効果を発揮すると考えられています。
- 第二世代の抗精神病薬やナルトレキソン: 過度の攻撃性や衝動性に対して反応することがあります。これらの薬物は、脳内のドーパミンなどの神経伝達物質の働きを調整することで、興奮や衝動性を抑える効果が期待されます。
- オキシトシンの実験的投与: オキシトシンは、「愛情ホルモン」とも呼ばれる神経ペプチドで、社会的 bonding や信頼感に関与するとされています。実験的な研究では、自閉症やアスペルガー症候群の患者において、音声からの感情認識(韻律、声のトーンなどから感情を読み取る能力)を改善する可能性が示されています。これは、オキシトシンが社会的な情報処理の一部を改善する可能性を示唆する興味深い知見ですが、まだ実験段階であり、一般的な治療法として確立されてはいません。
まとめ
この部分は、自閉症に対する包括的な支援として、非薬物的介入(社会的・コミュニケーション能力トレーニング、職業療法)、コミュニケーション上の配慮、家族支援の重要性を強調しています。心理療法は併存障害には有効な場合があるものの、自閉症そのものに対しては限界があることが示唆されています。薬物療法は、深刻な行動問題に対する最終的な手段として位置づけられており、特定の薬物が特定の症状の緩和に役立つ可能性が研究されています。特に、オキシトシンの実験的な効果は、今後の自閉症治療の新たな可能性を示唆するものです。
