
進化精神医学と心身医学の教科書:精神病理学の起源
マーチン・ブルーネ
精神医学と心身医学は、脳、心、行動に影響を与える医療条件に関心を持っています。従来のアプローチは、精神的および心身的な状態の潜在的な原因の限られた配列に焦点を当ててきました。これには、トラウマ、放置、虐待などの逆境体験、遺伝的脆弱性、遺伝子発現のエピジェネティックな調節が含まれます。これらは精神障害の理解にとって不可欠ですが、なぜ人間の心が機能不全に対して脆弱であるのかといった重要な問いを無視してきました。
「進化精神医学と心身医学の教科書」は、これらの問いに対する答えを提供するために、前版を更新し拡張しています。精神的および心身的な状態に対する進化的視点を強調しています。この教科書では、以下のことを説明しています:
- 人間の脳/心が自然選択と性的選択によってどのように形成されたか。
- 私たちの進化の過去における環境条件への適応が、現在どのように最適でない方法で機能する可能性があるか。
- 人間の認知、感情、行動がどのように科学的に枠付けられ、社会における地位、交配、ケアの提供と受け入れ、報酬のある関係を維持するための重要な生物社会的目標を達成しようとする理解を深めることができるか。
精神的および心身的な状態の理解に関連する進化的なトピックには、以下の概念が含まれます:
- 遺伝的可塑性
- 生活史理論
- ストレス調節
- 免疫学的側面
さらに、進化的な枠組みが、精神的および心身的障害を持つ患者の生活を改善するために、心理療法や精神薬理学がどのように機能するかを理解するためにも必要であると主張されています。
「進化精神医学と心身医学の教科書」は、健康と病気に関する進化的な問題を理解しようとする心理学、医学、心理療法のすべての学生にとって貴重なテキストです。
第2版の序文
この本の新しい版は、いくつかの顕著な変更と追加を示しています。1つは、臨床章が新しい(第5版の)精神障害の診断と統計マニュアル(DSM-5)に適応されたことです。DSM-5はその前の版と同様に進化的思考と互換性がありませんが、精神科医や研究者に精神的状態の進化的概念を親しんでもらう最良の方法は、最もよく知られ広く普及している既存の枠組みを利用することだと感じました。
内容的には、すべての章が広範に改訂されています。初版とは対照的に、章はすべて完全に引用されており、「選択されたさらなる読書」セクションは廃止されました。さらに、私の見解では精神障害の理解に深い影響を与えるいくつかの概念を導入または詳述しました。第1章には「差別的遺伝的脆弱性」の概念に関する新しいセクションが含まれており、同じ遺伝的変異(またはアレル)が逆境的な初期経験と関連している場合には障害に対する脆弱性を促進し、有利な環境条件と関連している場合には精神的疾患から保護する可能性があることを示唆しています。この基本的な概念を無視することは、現代精神医学における最も重要な科学的省略の1つかもしれません。第2章には、自律神経系(ANS)の進化に関する説明が含まれており、初期のストレスによるANSの持続的な変化が予後的な意味を持つ可能性があり、電気生理学的に容易にアクセスでき、治療の進捗を監視する窓口を提供するかもしれないことが述べられています。第3章では、生活史理論と生活史戦略のより詳細な説明が含まれています。この拡張は、ほとんどの精神的問題が生活史戦略の病理学的極端として見なされることを強調するために必要でした。読者が精神的状態に反映される「速い」および「遅い」生活史パターンを、複雑な遺伝子-環境相互作用から生じる生活史戦略の個人差に言及する際に、より容易に特定できることを期待しています。適切な場合には、臨床章でも生活史理論に触れています。さらに、第4章では(初期の)ストレスと免疫機能との関係、そしてこれが心理的健康にどのように影響するかについて詳述しました。免疫学的側面も臨床章に含まれています。
第I部は依然として5章で構成されていますが、臨床セクション(第II部)は現在14章で構成されています。具体的には、感情障害は現在、うつ病に関する章と双極性感情障害に関する章の2つに分割されています。同様に、強迫性障害(OCD)はDSM-5では不安障害の一部ではなくなったため、OCDおよび関連障害に関する別の章が含まれています。同様に、外傷およびストレス関連障害に関する新しい章が追加され、主に外傷後ストレス障害に焦点を当てています。さらに、新しいDSM-5のカテゴリーである身体症状および関連障害に関する新しい章も含まれています。同様に、性的機能障害に関する新しい章もあります。最後に、摂食および食事障害に関する章は、肥満に関する議論を追加することで拡張されました(肥満はDSM-5には含まれていませんが、「古典的」な摂食障害である神経性無食欲症や過食症と重要な特徴を共有しています)。第III部には、薬理学的治療の進化的考察に関する新しい章があります。
これらの多様な変更を考慮し、この本のタイトルを「進化精神医学と心身医学の教科書:精神病理学の起源」に変更することが適切であると考えました。これは、精神医学および心身医学のトピックに対する本のより広範な影響を認識するためです。
一般的に、私は初版の序文で説明した章の構造を保持しています。後者で言及し、感謝の意を表した学者に加えて、私が多くを学び、彼らの洞察を共有し、最新の文献を提供してくれた他の数人にも感謝の意を表したいと思います。進化的な健康と病気に関する記事の数が爆発的に増加しているため、見落としていたかもしれない文献が多くあります。したがって、私はリヤード・アベド、モハメド・アッバス、カール=ユルゲン・バー、ベルナール・クレスピ、ブルース・エリス、ホラシオ・ファブレガ、ジェイ・ファイアマン、世界精神医学協会(WPA)の進化精神医学者グループ、サー・ポール・ギルバート、シモーヌ・シャマイ=ツーリー、ダン・スタイン、ポール・セント・ジョン=スミス、ジェームズ・アンダーソン(「アンディ」)・トンプソン、アルフォンソ・トロイジ、ダン・ウィルソン、そして多くの他の人々に感謝しています。また、2010年に悲劇的な交通事故で亡くなった良き同僚で友人のエルウィン・ゲールツにも感謝の意を表します。彼の精神障害の行動学的側面に関する研究は、私の研究と臨床業務に深い影響を与えました。最後に、私の元博士課程の学生であるクリスティーネ・ハイニッシュ、エリオット・C・ブラウン、チュムフール・タス、クリスティーナ・ゴンザレス=リエンクレスに感謝します。彼らは進化的な精神医学のトピックに対する熱意と議論を提供してくれました。
ステファニー・チュガノフスキー、ベッティーナ・フィンガー、ペトラ・ネンゲルケン、ビルギット・ザンダーは、多くの新しい参考文献を収集し整理するのを手伝ってくれました。
最後に、私は私の患者たちに感謝したいと思います。彼らから「実生活」における生活史戦略や、過去と現在の逆境から生じた困難に対処するために何が役立つかについて多くを学びました。この本の新しい版が、研究者や臨床医にとって有用な洞察を提供できることを非常に期待しています。
マーチン・ブルーネ、2015年9月。
目次
はじめに
第I部 進化精神医学と心身医学の理論的背景
- 進化理論、進化的精神病理学、遺伝学の原則
- 人間の脳:解剖学、進化、機能
- 人間の生活史
- 精神病理学の原因
- 精神科評価
第II部 精神的および心身的障害
- 自閉症スペクトラム障害
- 注意欠陥/多動性障害
- 統合失調症スペクトラムおよびその他の精神病性障害
- 双極性および関連障害
- うつ病障害
- 不安障害
- 強迫性障害および関連障害
- 外傷およびストレス関連障害
- 身体症状および関連障害
- 摂食および食事障害
- 性的機能障害
- 物質関連および依存症障害
- 神経認知障害
- パーソナリティ障害
第III部 特別なトピック
- 自殺および自己傷害行動
- 精神障害の法医学的側面
- 心理療法
- 精神薬物療法 – 進化医学からの洞察
エピローグ:なぜ精神医学と心身医学は進化を必要とするのか
参考文献
著者索引
主題索引
はじめに
- 定義
精神医学と心身医学は、精神的および心身的状態の評価、記述、予防、治療に関心を持つ2つの関連する医療分野です。両者は社会科学と生物科学の接点に位置する学際的な分野です(Engel 1977; Fabrega 2007)。精神医学と心身医学は、心理学、哲学、倫理学、神経科学、生物学、薬理学、神経学、内科学、その他の医療専門分野からの洞察を利用しています。現代の精神医学は、生物精神医学、社会・地域精神医学、コンサルテーションおよびリエゾン精神医学、緊急精神医学、小児および青年精神医学、高齢者または老年精神医学、異文化精神医学、法医学精神医学など、いくつかのサブスペシャリティに分けることができます。さらに、心理療法は精神的および心身的治療の不可欠な部分です。さまざまな「流派」の心理療法が存在し、その中には学習理論に焦点を当てたもの(例:行動療法)や、患者の自伝的な資料に特に取り組むもの(例:精神力動的心理療法)があります。
- 認識論的問題
現代の精神医学と心身医学(およびほとんどの心理療法)の認識論的基盤は、明らかに自然主義に根ざしています。つまり、精神障害の科学的に価値のある理解は、認知、感情、行動の現象が中枢神経系の神経活動の結果であることを指します。より具体的には、人間は個人的な歴史(個体発生)だけでなく、系統発生の歴史も持っていることが暗黙のうちに認められています。言い換えれば、長い年月をかけて、自然選択と性的選択が、個体が内的および外的環境と効果的にコミュニケーションを取るための脳のメカニズムを形成してきました。精神的な生活は、そのすべての側面が、私たちの種や進化した祖先の遠い過去に存在した環境条件への適応であることを示しています。重要なのは、精神的状態は「正常性」と質的に異なるのではなく、変異の極端に位置しているということです。別の言い方をすれば、精神的状態を特徴づける可能性のあるほぼすべての兆候や症状は、一般集団において弱い表現で見られることができます(van Os et al. 1999, 2000)。
人間は本質的に群れを成す生き物であるため、人間の脳に表現される多くのメカニズムは社会的な問題に対処するために進化しました。したがって、人間の脳を「社会的脳」と呼ぶことができます(Brothers 1990a, 1990b; Dunbar 1998, 2003)。これは、精神医学に関して現在私たちが「生物学的」という用語を使用する方法が大いに貧弱であることを強く示唆しています。私たちの種の生物学的遺産は社会性と不可避的に結びついているため、ほぼすべての対人関係の問題には生物学的な次元があります。この視点は、精神的状態や治療アプローチを生物文化的文脈に置く重要性を軽視するものではなく、そうしなければ精神的状態や症候群の「意味」を完全に理解することはできません(Fabrega 2006)。いずれにせよ、さまざまな理由から、系統発生的視点は現代の精神医学によってまだ完全には認識されておらず、進化理論は医療カリキュラムに正式に取り入れられたことがなく、これは治療の進展に対する主要な障害と見なされるかもしれません(Nesse et al. 2006, 2008; Nesse et al. 2010; Stearns et al. 2010b)。現在の精神医学の概念化において進化を無視する理由のいくつかは、次のセクションで概説されます。
3. 歴史的ノート
「精神医学」という用語は、ヨハン・クリスティアン・ライル(Johann Christian Reil、1759-1813)によって1808年に造られました。当時、精神的に病んでいる人々の治療は医学の中で十分に統合されていませんでした。精神病は、脳の機能不全や不利な経験によるものではなく、個人的な精神的または道徳的失敗、あるいは神による罰の結果と見なされていました。そのため、多くの精神的に病んでいる人々は監禁され、残酷な扱いを受けていました。
フランスでは、フィリップ・ピネル(Philippe Pinel、1745-1826)と彼の弟子ジャン・エティエンヌ・ドミニク・エスキロール(Jean Etienne Dominique Esquirol、1772-1840)が、精神病は治癒不可能であり、精神的に病んでいる人々はその予測不可能な行動のために社会を守るために隔離されなければならないという一般的な見解に挑戦しました。代わりに、彼らは共感と慈悲を特徴とする「道徳的治療」(traitement morale)を導入し、科学的に根拠のある精神医学の分類法を発展させました。
ドイツでは、ヴィルヘルム・グリージンガー(Wilhelm Griesinger、1817-1868)が精神医学の主要な権威の一人となりました。1845年、彼は精神医学の最初の科学的教科書の一つ(『精神病の病理と治療』)を出版し、精神医学において自然主義的視点を採用し、精神病を「脳の障害」として特徴づける必要性を強調しました。グリージンガーは、たとえば、精神病性障害は認知機能の低下の異なる段階を経るが、精神的健康との連続体を形成すると仮定しました。この「単一精神病」の見解は、他の医学分野に倣って「自然な病気の実体」を現在の現象学や病気の「自然な」経過における症状の変化に基づいて区別しようとした多くの同時代の人々には満足されませんでした。
方法論的には、カール・ルートヴィヒ・カールバウム(Karl Ludwig Kahlbaum、1828-1899)が、偏見のない行動観察とすべての精神的および身体的(肉体的)兆候と症状の徹底的な記録と記述を含む「臨床的方法」を発展させました。カールバウムの意図は、経験的に得られた臨床資料を神経病理学的相関と結びつけることでしたが、彼の時代には成功しませんでした。カールバウムの「カタトニア」(「緊張精神病」; 1874)および「ヘベフレニア」(「青年精神病」; 1871; カールバウムの同僚で弟子のエヴァルト・ヘッカー(Ewald Hecker、1843-1909)がカールバウムの名のもとに執筆)に関する最も有名な出版物は、エミール・クレペリン(Emil Kraepelin)によって採用され、彼の「早発性痴呆」の概念に含まれました。カールバウムは、精神的な病気をカール・フォン・リンネ(Carl von Linné)の動植物の分類に従って分類しようとした試みも行い(『精神病の分類と精神障害の分類』; 1863)、このアイデアはクレペリンによって支持され、さらに発展されました。スイスの精神科医エウゲン・ブロイラー(Eugen Bleuler、1857-1939)は、後に「早発性痴呆」という用語を「統合失調症」に置き換えました。これは、すべての患者が避けられない認知の低下に関連する悪い予後を持っているわけではないことを強調するためのものでした。
興味深いことに、19世紀後半から20世紀初頭の多くの精神科医は、精神医学に対する進化論の強い含意を見出していました。これは部分的には、哲学的モニズムの受容が高まり、心身二元論が放棄されたことに基づいていました。一方で、生物学的進化と安定した進歩の混同は、精神病が自然選択の力の廃止の結果であるという見解につながりました(精神医学における生物学的進化の誤った解釈に関する批判は第1章の後書きを参照)。たとえば、クレペリンはキャリアの終わりに次のように書いています:
「人間の人格の発展(系統発生)は、無限に小さく、ほとんど知覚できない前進のステップによってのみ完成されてきました。後退のステップも発生しました。迂回が行われ、その後放棄されました。この予測不可能な進歩の最終結果は、さまざまな発展段階の痕跡や遺物を保持しています。たとえかつて形成され、次第に超越されたメカニズムの大部分が完全に失われたとしてもです。したがって、今日、私たちが精神病の表現を個々の人格発展の段階に適合させようとすると、必要な証拠はその欠如によって際立っています。このような試みが成功する場合、私たちの心理的生活の現れを子供の心理、原始人の心理、動物の心理に根ざして追跡する必要があります。このようにして、特定の病気が私たちの個人または系統発生的発展の歴史において隠されていた感情の再発をどの程度反映しているかを発見することができます。この点に関しては、私の知識が貧弱であるにもかかわらず、希望が持てると思います。この努力から、私たちの最も重要で最も困難な課題である病気の形態の臨床的理解に向けて助けを得ることができるかもしれません。」
(エミール・クレペリン(1920)『精神病の現れ』(ドミニク・ビアによる翻訳))
さらに以前に、ジェームズ・クライトン=ブラウン(James Crichton-Browne、1840-1938)は、「最後に組織され、最も高度に進化し、任意のものであり、脳の左側に位置する皮質中心が、精神病において最初に影響を受ける可能性が高いように思われる」と仮説を立てました(Crichton-Browne 1879, p.42)。若い頃、クライトン=ブラウンはチャールズ・ダーウィンを支持し、彼の著書『人間と動物における感情の表現』(The Expression of the Emotions in Man and Animals、1872)の出版に貢献し、いくつかの図を提供しました。クライトン=ブラウンは、神経系の病気に関連する解体の逆の進化的発展のアイデアを展開したジョン・ヒューリングス・ジャクソン(John Hughlings Jackson、1835-1911)と親交を結びました。ジャクソンは、脳の階層的な組織を提案し、自己反省の能力が脳の最も最近進化した部分である前頭前皮質に局在しているとしました。同様に、ヘンリー・モーズリー(Henry Maudsley、1835-1918)は、脳の進化を精神病理学を理解するための重要な側面として言及しましたが、精神障害の現れや治療に対する社会環境の影響を軽視することはありませんでした(Maudsley 1867, p.421)。
このように、精神医学の新しい医療分野で生まれた多くの重要な概念は、少なくとも部分的には進化的思考に根ざしていました(初期の精神分析も同様です;第22章を参照)。しかし、選択の単位が不明確であり、遺伝の法則が知られていなかったため、精神医学の分類法や「自然な病気の実体」の探求は依然として elusive(捉えどころのない)ものでした。さらに、20世紀の初めにメンデルの遺伝の法則がデ・フリース、ツェルマック、コレンスによって再発見されたにもかかわらず(それぞれ独立に)、多くの精神科医はこれらの重要な発見に気づかず、獲得した特性が遺伝するという見解を持ち続けました。同様に、その当時、選択は種レベルで行われるという見解が広まっており、個体生物レベルでの選択のメカニズムが発見されるまでにはさらに数十年を要しました。
しかし、より重要なのは、19世紀後半から20世紀初頭にかけて、精神医学は精神的入院患者の数が大規模に増加しているという観察に関心を持っていたことです。19世紀末には、ヨーロッパやアメリカ全土で何千人もの患者が大規模な精神病院で治療されるか、しばしば単に収容されていました。精神的に病んでいる人々の急増は、精神科医たちに自然選択の廃止が人口の退化を引き起こしたと結論づけさせました。この考えは、当時の文化的な悲観主義と奇妙に一致する人気のあるアイデアでした。
たとえば、クレペリンはジャワへの旅の途中で、精神障害が「原始的」な人種では比較的まれであり、精神病の予後が発展した世界よりも良好であることを観察しました。クレペリンはこれを、発展途上国の人々が病気に対してより大きな抵抗力を持っている結果だと解釈しました。精神病が家畜化によって引き起こされた退化の結果であるというクレペリンの意見は、精神医学の分類法に強い影響を与え、貧困、衛生状態の悪さ、教育の欠如といった精神病の可能な原因を大きく無視しました。
その結果、精神障害の効果的な薬物治療が利用できなかったため、多くの国が精神病の有病率のさらなる増加を防ぐために優生学的措置を導入しました。これには強制的な不妊手術が含まれていました。こうした手段を承認した多くの精神医学の権威は、個人を扱う医師ではなく、人口レベルでの精神衛生の擁護者として自らを位置づけていました(中には明らかに人種衛生を念頭に置いていた者もいました)。
精神医学における社会ダーウィニズムの考え方の導入と、精神病院における精神患者のケアの質の低さは、精神医学の悪評に寄与し、精神障害を理解するための強力なツールとしての進化論の拒絶につながりました。(余談として、「社会ダーウィニズム」という用語は、チャールズ・ダーウィンが進化論を社会政策に適用することを促進したと誤解されるべきではありません。逆に、ダーウィンはその問題に対して消極的でした。)さらに、特にナチス・ドイツや他の全体主義体制における政治的目的のための精神医学の悪用は、今日の公衆における精神医学の認識において依然として根本的な役割を果たしています。
4. 倫理的問題
ナチスによる人間実験の残虐行為の余波は、人間実験のための倫理規範、いわゆるニュルンベルク綱領を必要としました。1964年、世界医師会はヘルシンキ宣言を発表し、それ以来、個人の権利の認識、個人への尊重、自己決定、治療および研究に対するインフォームド・コンセントを明確に規制しています。
1977年、世界精神医学会はハワイ宣言を承認し、精神医学の実践に関する倫理的ガイドラインを設定しました。このガイドラインは、患者の診察、継続的な医学教育と知識の更新、人間の尊厳と患者の権利、機密保持、研究倫理を含んでいます。1996年のマドリード改訂版では、精神科医が安楽死、拷問、死刑、性選択、民族的または文化的差別に参加することを拒否する場合を含む特定の状況に関するガイドラインも含まれています。また、臓器移植、メディアとの関係、遺伝研究とカウンセリング、業界や第三者支払者との利益相反、精神療法に関する精神科医の行動規範も定義されています。これには、精神科医と患者の間の信頼と境界の侵害も含まれます。
全文は世界精神医学会のウェブサイトで入手可能です:http://www.wpanet.org/
5. 概念的問題
数十年にわたり、精神科医は精神疾患を概念化する際に、基盤となる焦点に応じて異なる方法に苦しんできました。指摘されているように、19世紀末から20世紀初頭には、精神障害を生物学的に捉える最初の波が見られました(Roelcke 1997)。これは、誤った生物学的前提に基づき、精神障害の原因として社会的要因をほとんど認識しないものでした。1950年代には、精神分析理論が精神医学の支配的な枠組みとなり、精神障害は抑圧された思考や感情、または不利な母子関係に根ざしているという見解が広まりました。これにより、「統合失調症を引き起こす母親」という概念が生まれました。
1980年代以降、新しい診断ツールの登場に伴い、精神障害の遺伝的基盤、脳の解剖学的異常、異常な神経伝達が異常な認知、感情、行動の相関として再び注目されるようになりました。それでも、精神分析(および行動主義)からの洞察を再定義し、精神障害の理解を不利な初期経験によって引き起こされるものとして新たな概念に再構築しようとする強い動きがありました。しかし、概念的な観点から見ると、生物学的原因(遺伝と神経伝達)と心理的原因(不利な対人関係の要因)は、長い間、精神障害の理解において対立するアプローチとして扱われてきました。
したがって、臨床的な観点だけでなく、研究の問題に関しても、これら二つの陣営は孤立した生活を送ってきました。これは、進化論に基づく包括的な概念的枠組みが十分に評価されていなかったためです(Nesse and Jackson 2006)。
精神障害の診断における困難を克服するために、精神医学は診断プロセスを形式化しました。最も広く使用されているマニュアルは、精神障害の診断と統計マニュアル(DSM)と国際疾病分類(ICD)です。これらのマニュアルは、診断の信頼性を向上させる利点がありますが、臨床現象の複雑さを減少させるという代償があります。これらは「非理論的」とされることが多いですが、さまざまな精神医学の「学派」間の妥協を表しています。さらに、これらは明示的にそう述べているにもかかわらず、明確な病気の実体の存在を示唆しているように見えますが、これは科学的証拠によって支持されていません。精神医学の学生や研修医にとって、精神医学における明確な診断カテゴリーが存在するという印象は、診断や治療に対する誤った確信を与えるかもしれませんが、精神障害は正常からの連続体として現れ、障害間の次元的または段階的な違いが規則であり、例外ではありません。
したがって、純粋に記述的なマニュアルは臨床医間での診断の合意を得るのに役立ちますが、その恣意性のために注意して使用する必要があります。DSMやICDが十分に反映していないのは、徴候や症状の現れにおける性別の違いや、異文化的な問題です。我々の診断システムは、有効な異文化比較には狭すぎます(Fabrega 2002)。これは、おそらくそれらが主に西ヨーロッパの小さな地理的地域に起源を持つためです(Burton-Bradley 1979)。
現在、精神医学の整合性は、そのサブスペシャリティが分離していることで危険にさらされています。生物学的精神医学、社会精神医学、精神療法は、臨床医と研究者の両方に適した統合的な枠組みを必要としています。たとえば、患者を診る臨床医は、患者の遺伝的構成や神経伝達物質受容体のプロファイル、機能的脳イメージング中の活性化パターンにアクセスできません。臨床医の仕事は、対面の設定で患者の認知、感情、行動を理解することです。一方、研究者は科学的進展を遂げるために、分類学的システムを放棄する必要が高まっています。個々の遺伝子の遺伝的変異や機能的脳イメージング中の活性化パターンの寄与は、せいぜい症状特異的であり、症候群(または障害)特異的ではありません。記述的アプローチに従い、障害はその症状、疫学、遺伝的および環境的要因、病態生理、遺伝子と環境の相互作用の観点から理解される必要があります。この際、「ライフヒストリー理論」がこれらの情報源を統合するためのガイドとして機能することができます(Ellis and Del Giudice 2014)。
これらすべては、個人の認知、感情、行動の機能を理解することを目指しています。これは、我々が「正常な機能」と呼ぶものにおいても十分に難しいですが、障害のある状態ではさらに困難です。精神障害は明らかに異常で不適応な機能状態を反映しており、徴候や症状の現れが時には機能的に無意味に見えることがあります。しかし、精神病理学的な徴候や症状が「正常性」とは種類ではなく程度によって異なるという次元的な視点を考慮すると、対応する適応メカニズムの分析は異常な精神現象のコミュニケーション的側面に光を当てるかもしれません。
精神障害の理解において重要なもう一つの側面は、多くの精神的状態が脳だけでなく、身体全体に影響を及ぼすという観察にあります。これは、摂食障害や身体症状障害において最も明白です。しかし、エピジェネティックなストレス生理学の調節異常や免疫反応が、すべての主要な精神的状態の重要な側面であることを示す証拠が増えています(McEwen 2002; Ottaviani 2011)。要約すると、精神的状態は他の医療状態と同様に、ダーウィン医学(Nesse and Williams 1994)や「ハミルトン医学」(Crespi et al. 2014)の視点から全体的に分析されることが不可欠です。これらの問題は次の章で扱われます。
Part I
Theoretical background to evolutionary psychiatry and psycho –
somatic medicine
第1章
進化理論、進化精神病理学、遺伝学の原則
要約
ダーウィンの自然選択と性的選択による進化に関する研究は、生命が変化する環境への適応を通じてどのように発展したかを説明する生物学における中心的な科学的枠組みです。進化は、人間の脳と心を形成する原動力であり、身体的特性と同様に形成されてきました。人間の認知、感情、行動に関連する多くの適応は、進化的適応環境において生じましたが、現代の生活条件は何らかの形でそれから逸脱しています。このような進化した特性と現在の環境との「ミスマッチ」は、認知、感情、行動特性の機能不全に対する脆弱性を引き起こす可能性があります。遺伝子と環境は多様な方法で相互作用しますが、遺伝的可塑性は機能不全に対する脆弱性をもたらすだけではありません。むしろ、環境の逆境と関連する場合に機能不全を引き起こす可能性のある同じ遺伝的変異が、より好ましい環境においては機能不全に対する保護的効果を発揮します。これらの洞察は、精神医学と心身医学によってまだ認識されていません。
キーワード
- 自然選択
- 性的選択
- 進化的適応環境
- 遺伝的可塑性
- 遺伝子と環境の相互作用
1.1 進化理論、進化精神病理学、遺伝学への序論
伝統的に、精神的および心身的障害の原因や、それに伴う精神病理学的および身体的徴候や症状は、胎児発達中の発達的混乱や、幼少期、初期の子供時代、または思春期の逆境的な出来事の観点から概念化されています。神経生物学的レベルでは、これらは神経回路の機能不全、神経伝達物質の不均衡、または遺伝的脆弱性の観点から説明されます。たとえば、うつ病は、愛着対象からの分離などの幼少期の逆境的な出来事の結果として、セロトニン欠乏症候群として、またはセロトニントランスポーター遺伝子のアレル変異と関連付けられることがあります。心理的機能不全の行動的および神経生物学的根源に関する研究は、精神病理学の理解を大いに向上させたことは間違いありませんが、より広い生物学的視点から見ると、これは物語の半分に過ぎません。
精神医学と心身医学における一般的な誤解は、個人の初期発達条件や脳機能の神経化学または分子レベルの理解が、認知・感情・行動システムや心身システムの病理を理解するのに十分であると仮定することです。これは、個体発生と生理学だけで動物行動の本質を理解できると仮定するのと同じくらい誤りです。20世紀の偉大な動物行動学者の一人であるニコラス・ティンバーゲンは、これらのレベルを行動の近接的または即時的な原因と呼びました(Tinbergen 1963)。行動の近接的原因は、個体発生的発達と生理的メカニズムを含みます。行動(および病理)の近接的原因は、個人の生涯の中で変化する可能性があります。しかし、ティンバーゲンが指摘したように、行動を完全に理解するためには、行動の最終的なレベルも認識することが不可欠です。最終的または進化的レベルは、行動の系統発生的発展の再構築を含みます。つまり、密接に関連する種を研究することによって行動の進化的根源を遡る試みと、特定の特性の選択的利点または適応的価値を分析することです。これらの4つの「なぜ」の質問、すなわち個体発生、生理的メカニズム、系統発生、適応的価値は、行動を完全に理解するための重要な要素を構成します。近接的および進化的(最終的)レベルは、決して相互排他的ではありません。むしろ、基本的に補完的です(Nesse 2013; Brüne 2014b)。
近接的メカニズム(個体発生と生理学)と最終的原因(系統発生と適応機能)は、行動の理解に不可欠と考えられる補完的な次元です。
この視点から見ると、人間の認知、感情、行動は、すべての4つの質問に対処した場合にのみ理解されることができ、適応不全の特性も同様に分析される必要があるという明示的な理解が、本書の主なテーマを構成しています(Nesse 2013; Brüne 2014b)。しかし、これは、精神病理学的な徴候や症状が適応を表すものであるということを意味するものではありません。逆に、徴候や症状は、用語の一般的な理解と進化的な意味の両方において、真に不適応的です。精神病理学的な徴候や症状は、現在の文脈における異常な頻度、強度、または不適切さのために機能不全となった適応特性の極端な変異を反映しています(Brüne 2002)。たとえば、恐怖は個体に対する脅威を示し、環境の危険を回避するのに役立つ古代の適応特性です。それに対して、病的な不安は「本当の」脅威をもたらさない状況で発生するため、不適応と見なされますが、影響を受けた個人は特定の状況を危険だと主観的に認識するかもしれません。恐怖反応は通常、引き出しやすく、進化的に見て引き出しの閾値を低く保つことが理にかなっています。もし閾値が高すぎれば、恐れを知らない個体は非常に早く命を落とし、したがって生存する子孫を残さなかったでしょう。
精神病理学的な徴候や症状は、適応特性の機能不全の極端な変異を構成するという理解をもって分析されるべきです。
進化の過程において、実際の危険に直面したときに恐れを知らないことよりも、時には不必要に恐れる方が「安上がり」であった(Nesse 2001)。これは、恐怖や不安が多くの精神病理学的状態の一部である理由の一つかもしれません。さらに、恐怖や不安のケースは、生理学と病理学、適応と不適応の間に明確な境界を引くことの難しさの典型例かもしれません。しかし、うつ病は単なる誇張された悲しみではなく、迫害妄想は単に極端な疑い深さに似ているわけではないことを強調する必要があります。加えて、精神病理学的な徴候や症状は、適応特性の極端な表れであり、その不適応性は、医療的および/または心理療法的支援なしに自発的に回復する能力の限界または失敗によって表現されます。自己治癒能力の障害は、早期の無視や虐待の経験(これはストレスシステムの機能不全と関連している可能性があります;Brody et al. 2013)から、精神作用物質の中毒効果、対人関係の葛藤に永続的に閉じ込められること、自然選択が人間の心が対処するのに十分に装備していない達成不可能な生物社会的目標まで、さまざまな要因によって引き起こされる可能性があります。
精神病理学的な徴候や症状は、適応特性の極端な表れであり、その不適応性は、医療的および/または心理療法的支援なしに自発的に回復する能力の限界または失敗によって表現されます。発熱や咳嗽と同様に、これらは病原体に対する自然な防御メカニズムですが、心理的防御は、システムが疲弊したり、因果イベントの「病原性」に圧倒されたりすると崩壊する可能性があります。
発熱や咳嗽と同様に、病原体に対する自然な防御メカニズムである心理的防御は、システムが疲弊したり、因果イベントの「病原性」に圧倒されたりすると崩壊する可能性があります。このアナロジーによれば、悲しみは発熱に似ていると見なされるかもしれませんが、うつ病は敗血症性ショックに例えられるかもしれません。これらの簡単な例は、次のような重要な質問を引き起こします。「私たちの種はどのような状況に適応したのか?」「人間の心を形成した環境条件は何だったのか?」「人間の状態の出現に伴う進化的コストは何だったのか?」「なぜ心は逆境に対処するのにより良く適応していないのか?」(Gilbert 1998; Brüne 2014b)。
したがって、複雑な認知・感情・行動システムの機能不全を理解したいのであれば、ティンバーゲンが提案したように、その機能をすべてのレベルで理解する必要があります。次のセクションでは、心理的メカニズムと病理的変異の進化的(最終的)原因を理解するために不可欠な基本的な進化概念を要約します。
1.2 進化の定義
進化は、直接観察することができない歴史的なプロセスです。観察から推測しなければなりませんが、進化的プロセスが人間の認知、感情、行動を解剖学や生理学と同様に形成してきたことに疑いはありません(Barkow et al. 1992; Garcia 1996)。共通の祖先からの進化は非常に保守的であり、かなり遅いプロセスです。たとえば、チンパンジーと人間の分子の類似性は、両系統が約500万〜700万年前に共通の祖先から分岐したことを示唆しています(Kaessmann and Pääbo 2002)。これは約3500万の単一ヌクレオチドの違いに相当します(Varki and Altheide 2005)が、酵母と人間にもいくつかの共通の遺伝子があります。
自然選択と性的選択による進化は、人間の認知、感情、行動を形成してきた歴史的なプロセスです。
進化は、種の個体間で交配可能な個体の集団レベルで起こり、個体の遺伝的変動を反映します。進化が非常に遅いのは、数百世代または数千世代にわたる安定化選択が、集団内の個体間の変異を減少させる傾向があるからです。安定化選択は、特定の生態的ニッチに適応した最適な遺伝子型に集団を近づける傾向があります(Mayr 2001)。しかし、集団内には個体間で遺伝的および表現型の変異が存在し、これは選択が作用するための「原材料」を生み出すために必要です(Abecasis et al. 2012)。定義上、成功裏に繁殖する個体は、現在の環境条件に最も適応していると見なされます(Darwin 1859; 誤解を招く形で「適者生存」と要約される)。突然変異によって生じる新しい変異のほとんどは不利であるため、選択は主に適応が不十分な変異の排除プロセスとして機能します(負の選択)。しかし、新たに発生した特性が個体の繁殖適応度を高め、集団の遺伝子プールに固定される場合、正の選択が起こることもあります。これにより、選択は個体の表現型のレベルで作用することが示唆されます(Mayr 2001)。より過激な見解では、選択は個々の遺伝子に作用し、遺伝子同士が競争することができるとされています(Dawkins 1976)。選択が作用するレベルについての議論は完全には解決されていませんが、以前の提案とは対照的に、選択が種レベルで行われるという仮定は捨てられ、特定の特性がその特性を欠く他の集団に対して競争優位をもたらす場合、集団選択が役割を果たす可能性があります(Boyd et al. 2003)。
- 安定化選択は、交配可能な集団内の個体間の違いを減少させる傾向があるが、個体間の小さな違いも生殖成功の差に寄与する。
ほとんどの突然変異は有害であるため、選択は主に排除のプロセスです。しかし、時折、正の選択が特定の特性の急速な普及をもたらすことがあります。
選択は主に個体レベルで行われます。ほとんどの適応は設計上最適ではなく、妥協を表しています。これは、なぜ一部の不適応的特性(精神病理学に関連するものを含む)が選択によって排除されないのかを部分的に説明します。
自然選択による進化は「倹約的」なプロセスです。これは、適応が設計上最適ではなく、適応特性の経済性と信頼性の観点から利益とコストの間の妥協を表すことを意味します(Williams 1966)。これは、なぜ病理(精神病理学を含む)が存在し、選択的力によって排除されていないのかという質問に取り組む際に重要です(Nesse and Williams 1994; 第4章を参照)。
個体の表現型は、その構造や生理学だけでなく、認知、感情、行動の構成要素からも成り立っていることに注意することが重要です。これは「拡張された」表現型と呼ばれます(Dawkins 1982)。拡張された表現型は選択の対象となる可能性があり、これは人間の心理的特性の研究にとって特に重要です。
人間においては、個体選択と集団選択の両方が協力、道徳、社会的認知能力の進化に寄与した可能性があります。
集団選択は、高度に協力的な種において発生するメカニズムであり、集団の純粋な適応度の利点がすべての個体の適応度の算術平均を上回る場合に起こります。人間においては、個体選択と集団選択は補完的です。たとえば、遺伝的に無関係な個体間の協力は、生存と繁殖の観点から集団に利益をもたらすことがあります(Boyd et al. 2003)。これは、個体の認知的および感情的適応を形成し、「フリーライダー」を検出し、集団基準の違反に対して道徳的な罰を課す能力が生まれることにもつながります(Boyd et al. 2003)。これは、社会環境からの情報を処理するための人間の脳の特化を理解する上で特に重要です(「社会脳仮説」;第2章を参照)および、欺瞞に対する過剰警戒などの特定の病理を理解する上で重要です。その極端な例は、精神医学用語で言うところの「迫害妄想」を伴う症候群を思い起こさせます(Zolotova and Brüne 2006)。
1.2.1 自然選択と性的選択
進化的プロセスは、理論的には自然選択から生じる特性と、性的選択から生じる特性に分けることができます(Darwin 1859, 1871)。一般的に言えば、自然選択は個体の生存を助ける特性を好みます。たとえば、捕食者から逃げるためには、速く走ること(またはカモフラージュすること)が有利であり、速いランナーは平均して遅いランナーよりも生存する子孫を残す可能性が高くなります。捕食者種もまた選択にさらされており、最も熟練したハンターが最も繁殖する可能性が高いのです。この自然界における競争の原則は、進化における複雑さと体の大きさの増加への傾向、すなわちアナゲネシスを観察できる理由の一つかもしれません(Mayr 2001)。進化的理論家は、これを共進化的な「軍拡競争」またはレッドクイーン原理(ルイス・キャロルの『不思議の国のアリス』に由来)と呼び、増加する複雑さが少なくとも同じ場所に留まるために必要である(絶滅しないために)という観察の比喩としています(Dawkins and Krebs 1979)。
- 自然選択と性的選択は、種の進化を促進する二つの別々のプロセスです。
- 種間および種内の「軍拡競争」は、適応的修正の速度に寄与します。
進化的「軍拡競争」は、種間および種内で発生します。たとえば、人間においては、遺伝的に無関係な個体間の協力を高める選択が、「心の理論」や「メンタライジング」、すなわち自分自身や他者の精神状態を認知的に表現する能力、他者への共感的関心など、多くの社会的認知能力の進化に対してかなりの圧力をかけた可能性があります(詳細は第2章を参照)。自然選択された特性は、基本的に両性において同じです。対照的に、同じ種の雄と雌の間で異なる特性は、性的選択の産物です。
チャールズ・ダーウィンが性的選択を第二の(そして少なくとも同等に重要な)進化的力として発見する前、彼は、主に雄に見られるいくつかの特性が明らかに生存上の不利を示すことに混乱していました(Darwin 1871; Hiraiwa-Hasegawa 2000)。たとえば、孔雀の尾は非常に目立ち重いため、特に最も大きな尾を持つ個体が捕食者から逃げるのを妨げます。しかし、これらの個体は雌によって交配相手として優先的に選ばれます。このような誇張された形態的特徴の進化を説明するために、二つの仮説があります。一つは「遺伝的浮動」と呼ばれるメカニズムで、これは小さな繁殖集団におけるアレル頻度のランダムな変化によって、いわゆるランナウェイ選択を通じて特定の特性の頻度が増加することを指します。ランナウェイ選択は、偶然に雌によって好まれる特性が、次世代において雄によって過剰に表現され、さらに雌の好みが強まるメカニズムとして理解されます(これは一部の人間の心理的特性にも当てはまる可能性があります;Nesse 2007)。もう一つは「良い遺伝子」仮説または「ハンディキャップ原理」と呼ばれます。たとえば、孔雀においては、大きな尾は良い遺伝子を示す可能性があります。なぜなら、その尾を「持つ余裕」がある個体は病原体に対して抵抗力がある可能性が高いからです(Alcock 2001; Rossano 2003)。さらに、雄の雌に対する魅力は対称性の問題でもあります。「良い遺伝子」は、変動対称性が減少していることと関連していると提案されています。なぜなら、病気のプロセスは特に初期の発生過程において大きな非対称性を引き起こす可能性があり、これは魅力が低いと見なされるからです(これは統合失調症に関して提唱された仮説です;Yeo et al. 1999; 第8章を参照)。
性的選択は、孔雀の尾のように生存において明らかに適応上の不利をもたらす特性が存在するため、チャールズ・ダーウィンにとって長い間謎でした。性的選択によって選ばれる特性は、「遺伝的浮動」によって生じることもあれば、優れた遺伝的質を示すこともあります。
ほとんどの種において、雄は雌へのアクセスを求めて互いに競争します(同種内競争;Buss 1988a)。したがって、選択は、特に一頭の雄が複数の雌を支配する種において、雄の体の大きさや力を優先します。これは性的二形性として知られています。領土に対する同種内競争は、体の大きさや物理的な力だけでなく、角や大きな犬歯などの武器によっても解決され、雄は支配的な階層を形成します。雄が雌に対してほぼ平等にアクセスできる場合、特別なタイプの同種内競争が発生します。このような場合、精子の量と質が、どの雄が受精した雌の子孫を父親にするかを決定することがあります。したがって、これは同種内競争が行われる別のレベルを表しています。一部の個体は「神風精子」と呼ばれる、非常に異形に見えるが他の精子に対して毒性を持つ精子細胞を生成することさえあります(Baker and Bellis 1995; 詳細は1.2.2.2節を参照)。
- 同種内競争は、肥沃な雌へのアクセスを求める雄の間で、体の性的二形性と精子競争の進化をもたらしました。
- 対照的に、ほとんどの種(人間を含む)では、雌が雄の交配相手の質を評価して選択します。
性別間の競争も存在し、これは異種間競争と呼ばれます。雌の選択の原則はすでに言及されています。雌が雄よりも交配相手の選択においてより選択的である理由は、ほとんどの種において雌が潜在的な子孫に対して雄よりも多くの投資をするからです(1.2.2.2節を参照)。言い換えれば、雌は最良の交配相手を徹底的に選ばなかった場合、繁殖適応度に関して雄よりもはるかに多くのものを失うことになります。雌の選択の背後にある論理は、繁殖適応度は単に子孫の数によって決まるのではなく、次世代の遺伝的質によっても決まるということです。したがって、雌にとっては遺伝的に高品質な雄を選ぶことが利益となります(Buss 1998)。多くの鳥類のような一夫一妻制の種では、雌はしばしばペア外交尾を行います。逆に、雄は通常、交配相手の不貞を防ぐために非常に警戒しています(Buss and Shackelford 1997)。私たちの種において、男性は配偶者に対する身体的暴力や、さらには強姦といった戦略を進化させた可能性があります(Wilson and Daly 1996; MacDonald 2000; Thornhill and Palmer 2000)。これらのメカニズムは、種の社会構造や他者との遺伝的関連性に応じて、雄と雌の行動に深い影響を与えます。
1.2.2 包括的適応度理論
進化理論の現代的な統合は、個体の繁殖成功が差異的進化発展の中心テーマであることを発見しました。包括的適応度理論は、繁殖成功は個体の子孫の数(古典的適応度)だけでなく、遺伝的に関連する個体の繁殖成功にも依存することを提案します(Hamilton 1964)。したがって、近親者の繁殖努力を支援することは、適応度の観点から有利である可能性があります。言い換えれば、包括的適応度は、個体の繁殖成功と、親族の成功をその関連度に応じて加算したものです(例えば、兄弟姉妹はいとこよりも密接に関連しています)。選択は、繁殖成功を最大化する行動を好むべきです。なぜなら、遺伝的な観点から見ると、個体が自らの遺伝子を次世代に伝えることが利益だからです。したがって、遺伝的に動機づけられた利他的行動は親族に限定されるべきであり、実際に利他的行為は社会性昆虫や裸のモグラネズミのような類似の社会構造を持つ他の種を含む動物界に広く見られます(Trivers and Hare 1976)。対照的に、人間社会では、遺伝的に遠いか無関係な個体間でも相互援助がしばしば見られます(Hill et al. 2011)。これは説明を必要とします。なぜなら、これは包括的適応度理論に矛盾しているように見えるからです。
自己中心的な行動と利他的行動に関する意思決定は、資源配分に関する対立を伴います。これは、遺伝的に無関係な個体だけでなく、異性の個体間でも発生する可能性があります。なぜなら、繁殖成功を最大化するための戦略が根本的に異なる可能性があるからです。対立は親と子の世代間でも発生します。なぜなら、親は平均して個々の子孫と50%の遺伝物質を共有するからです。これらの対立の領域は、ロバート・トリバースによって三つの重要な論文で特定されています。
- 包括的適応度理論は、個体の適応度は個体自身の繁殖成功と遺伝的に関連する個体の繁殖成功の合計であることを示唆しています。
1.2.2.1 互恵的利他主義
互恵的利他主義は、遺伝的に無関係な個体間の利他主義の一形態として説明できます。これは、一方の生物(受益者)に利益をもたらし、他方の生物(利他的行為を行う生物)に対してコストを伴うものであり、将来の利他的行為の返礼が不確実である場合を指します(Trivers 1971)。食物の共有は、多くの種で見られるこのような行動の一例です。互恵的利他主義は、利他的に行動する個体がシナリオとその恩恵を受けた個体を記憶する心理的メカニズムを持っている場合にのみ適切に機能します。さらに、互恵性の重要な特徴は、受益者が返礼を拒否する場合にそれを認識できることです。詐欺を検出することは、欺瞞的行動に対抗するために人間の系統で選択された必要な認知メカニズムの一つです。さもなければ、欺瞞的傾向を好む遺伝子が集団に広がり、相互援助のシステムが崩壊してしまうでしょう(Trivers 1971)。
- 互恵的利他主義の理論は、遺伝的に無関係な個体間の利他主義は、利他的行動が返礼される限り持続可能であり、非協力的行動を検出し、返礼を強化するためのメカニズムが存在する場合に限ると提案します。
互恵的利他主義に関連する問題は、祖先の(そして現代の)人間社会において非常に広範囲に存在しました。なぜなら、交換の機会が最も大きいのは、低い分散率と高い相互依存度を持つ長寿命の種において、親の世話が長期間にわたる場合だからです(Bell 2001)。したがって、危険な時の相互援助、食物の共有、病気の人や非常に高齢または非常に若い個体の助け、道具や知識の共有は、すべての人間文化に見られる利他的行動です(Trivers 1971)。トリバースは、社会集団や部族に属する個体間で互恵的利他主義を維持するために進化したいくつかの認知的および感情的特性を特定しています。友情、同情、感謝などのポジティブな感情は、利他的パートナーシップにおける感情的報酬として進化した可能性があります。対照的に、道徳的攻撃を含むネガティブな感情は、欺瞞的傾向を減少させ、返礼がない場合の利他的行動の継続を防ぐかもしれません。欺瞞の試みが検出された場合、罪悪感や恥といった感情(欺瞞者側)は、互恵的パートナーシップを回復するのに役立つかもしれません。認知レベルでは、道徳的攻撃、同情、または罪悪感を装って他者に利他的行動を誘発することも可能です。このような微妙な欺瞞メカニズムは、誠実な行動と偽善的な行動を区別する能力の選択につながった可能性があります。信頼と疑念の感情は、互恵的利他主義の問題に直接関連して進化した可能性があります。また、社会的規範の違反が発生した場合に協力を再構築するメカニズムとして、許しが進化したという証拠もあります。同様に、共通の利益に寄付することは、集団の結束を強化する目的を果たすかもしれません(McCullough et al. 2013)。さらに、互恵性を維持することは、個人的に重要な社会的相互作用を数年または数十年後に思い出すことを可能にする自伝的記憶の進化にも寄与した可能性があります(Suddendorf 2013)。
1.2.2.2 親の投資と性的選択
人間において、同情、恥、罪悪感などの社会的感情は、相互性に関連する問題に応じて進化したと考えられています。さらに、許しは社会的規範の違反が発生した際に協力を再構築するメカニズムとして進化しました。
集団選択は、欺瞞者に対する集団的な罰を好むことにも寄与した可能性があります(Boyd et al. 2003)。初期のヒト科の集団における個体間の非常に広範な相互依存を考慮すると、集団の結束や社会的ルールや規範への従順が選択圧の対象であった理由が明らかになります。人間は、グループメンバーの非協力的行動を罰するために追加のコストを負担することさえ厭わないという豊富な証拠があります(Fehr and Rockenbach 2004)。
精神病理学の観点から、相互利他主義の問題に関連する二つの側面が表面上の価値を持っています。一つは、疑念の極端な変異、すなわち偏執的思考であり、最終的には迫害妄想や参照妄想につながる可能性があります。興味深いことに、迫害妄想の内容は、影響を受けた個体が他者に悪意のある意図があると確信していることがほとんどです(つまり、彼を欺こうとしている)。迫害妄想の内容として善意の意図が含まれることは例外です(詳細は第8章を参照)。もう一つは、欺瞞の極端な変異、すなわち精神病質です。精神病質は、集団内で低い有病率で存在する欺瞞者の形態として解釈されています。現在の概念化では、精神病質はそれ自体が病理を表すものではないとされており、反社会的人格障害との重複があるものの(Mealey 1995; Troisi 2005; 詳細は第19章を参照)、精神病質は病理そのものを示すものではありません。
迫害妄想のような精神病理学的症候群は、欺瞞的行動の検出に関与するメカニズムの極端な変異として見ることができます。対照的に、精神病質は、自分の利益のために他者を欺くことを過度に利用する行動傾向を反映しています。
さらに、協力、相互性、信頼、公平性の規範を理解するための神経経済学的アプローチは、これらの複雑な社会的行動に影響を与える環境的条件や、精神的疾患に関連する社会的意思決定の変化に関する新たな洞察を提供します(Wischniewski et al. 2009; Kishida et al. 2010)。
性的繁殖には、無性繁殖に比べていくつかの欠点があります。
- 性的繁殖は、適切な交配相手を見つけるために時間とエネルギーを要し、遺伝物質を無駄にします。なぜなら、個体の遺伝子の50%しか子孫に受け継がれないからです。
- 性的繁殖が進化した理由は、遺伝子の再結合によって有害な突然変異の蓄積の影響を相殺するためです。
- 同様に、性的繁殖は、急速に進化する病原体や寄生虫に対する防御システムを改善する最も重要なメカニズムです(Ridley 2004)。
- 性的繁殖は、遺伝物質の回転が遅い長寿命の種にとって特に重要です。
進化は二種類の生殖細胞を生み出しました:
- 大きな生殖細胞(卵):かなりの代謝資源を必要とします。
- (エネルギー的に)「安価な」生殖細胞(精子):非常に大量に生成されます。
これらの異なる種類の生殖細胞は、慣例的に「雌」(卵)と「雄」(精子)と呼ばれます。雄と雌の生殖細胞の違いは、子孫への親の投資の量や繁殖戦略に重要な影響を与えます。
- 性的繁殖は、無性繁殖に対して有害な突然変異の蓄積を相殺し、急速に進化する病原体からの脅威に対してより良く対処するという利点があります。
トリバースは親の投資を「親が個々の子孫に行う投資であり、それが子孫の生存の可能性(したがって繁殖の可能性)を高める一方で、親が他の子孫に投資する能力を犠牲にするもの」と定義しました(Trivers 1972)。哺乳類においては、妊娠、授乳、育成、子孫の保護は雌の生物の支出であり、ほとんどの種では雄は受精以上の投資を行いません。この予測から導かれるのは、潜在的な子孫に多く投資する性(通常は雌)が交配相手の選択においてより選択的であり、投資が少ない性(通常は雄)がより多くの投資を行う性に対して性内競争を行うが、交配相手の繁殖適応度に関してはあまり選択的でないということです(Buss and Schmitt 1993)。これは、繁殖がエネルギー、資源、時間の面で高コストであり、これらの制約が雌にとってより重要であるためです。なぜなら、雌の繁殖成功は成熟した卵の数によって制限されるからです。対照的に、雄の繁殖成功は(理論的には)アクセス可能な成熟した雌の数によってのみ制限されます。言い換えれば、子孫を生産するために最も多く投資する性が、他の性が競争する制限資源となります(Buss 1999)。
霊長類、特に人間においては、繁殖成功はさらに多くの要因によって制約されています。これには、非常に長い幼少期、繁殖の高年齢、単胎出産、長い出産間隔が含まれます。その結果、選択は子孫に多く投資する意欲のある人間の雄を好むようになりました。したがって、ほとんどの人間社会(おそらく祖先の社会も含む; Walker et al. 2011)では、男性は通常一人または少数の妻(多妻制)を持ち、これは人間の雄が他の多くの霊長類に比べてより大きな父親の投資を行うことに直接関連しています(Møller and Thornhill 1998)。非常に稀に一妻多夫制が観察されることがあります。それでも、平均して人間の母親は父親よりも子供に多く投資しており、女性にとっては、子供を育てるためにパートナーからの支援を得るために、長期的な関係を追求する意欲と資源を持つ交配相手を選ぶことが利益であったに違いありません(Hrdy 2000)。逆に、親の投資理論からは、人間の男性が複数の女性と交配することが利益であると考えられます。
ほとんどの哺乳類において、雌は雄よりも潜在的な子孫に多く投資します。その結果、雌は交配相手の選択においてより選択的です(雌の選択原則)。
霊長類、特に人間においては、繁殖成功は長い幼少期、遅い繁殖、長い出産間隔によって制約されており、これが多くの他の哺乳類に対する父親の投資の増加を促進しました。
これらの予測に沿って、選択は、交配相手の監視、嫉妬の感情(Wiederman and Kendall 1999; Buss 2000a, 2000b)、隠れた排卵、そしておそらく性的強制(「雌の選択」原則を回避するため; Voland 1998; Buss 1999)を含む、いくつかの感情的、認知的、生理的適応を好むようになりました。これらの適応は行動レベルでは霊長類や人間に特有のものではありませんが、主観的な感情の次元はおそらく特有のものです。嫉妬は男性と女性の両方にとって重要な感情ですが、異なる理由から生じる可能性があります(Daly et al. 1982)。
女性は、交配相手が他の女性に奪われることを恐れるため、より感情的に嫉妬するかもしれません。対照的に、男性は不確実な父親性の問題に直面するため、より性的に嫉妬することが期待されます(Wiederman and Allgeier 1993; Sheets et al. 1997)。つまり、女性は自分の子供の母親であることを100%確信できますが、父親は同じように確信することができません。実際、浮気は多くの種、特に一夫一妻制の鳥類において広く見られ、この現象は集中的に研究されています。現代の人間社会では、最大で10%の子供が生物学的に父親と血縁関係がないとされています。実験的な研究は、男性がパートナーから離れた後に不確実な父親性のリスクを補うために、より多くの精子を生成するように選択されてきたことを示しています(Baker and Bellis 1995)。さらに、陰茎の亀頭の解剖学的形状は、他の男性の精子を膣から取り除きながら、自分の精子を挿入するための適応として解釈されています。これらのメカニズムは、いわゆる「精子競争」に関連している可能性があり、これは交尾後の雄間の競争の遅延を指します(Baker 1993)。
人間における男性と女性の親の投資の相対的に大きな量は、父親性と感情的なコミットメントを確保するための嫉妬のような感情的適応を選択する要因となっています。
ただし、一夫一妻制は人間に進化した唯一の交配戦略ではないかもしれません(Rutberg 1983; Brewer et al. 2000)。実験条件下では、排卵に近い若い女性は対称的な男性の体臭や男性的な顔の特徴を(無意識に)好む傾向がありますが、実際の交配相手の選択は男性が長期的な関係を追求する意欲によって影響を受けることがあります。私たちの最も近い親戚を詳しく見ることで、選択が交配システムにどのように異なる影響を与えてきたかを示すことができるかもしれません(McGrew et al. 1996)。例えば、ゴリラはオスとメスの間に大きな体サイズの違い(性的二形)を特徴としています。しかし、ゴリラの精巣のサイズは体サイズに対して小さく(約30g)、これはゴリラの社会構造が一雄多雌の多妻制として最もよく説明されるためです。対照的に、チンパンジーでは性的二形は比較的小さいです。それでも、オスのチンパンジーは成体のシルバーバックゴリラのサイズの三分の一しかないにもかかわらず、約120gという非常に大きな精巣を持っています。この不一致の進化的説明は、チンパンジーが多雄多雌の群れで生活し、メスが性的に受容可能なときに複数のオスと交配する「乱交的」な行動をとるためです。したがって、オスのチンパンジー間の性内競争は、オスがメスの膣内に置くことができる精子の量の変動によって交尾後に移行します(精子競争)。ただし、メスのチンパンジーは、複数のオスからの精子を受け入れることで、卵子に到達する精子が競争力を持つことを確保するために、精子競争の過程で積極的な役割を果たす可能性があります(したがって、その生産者の「良い遺伝子」を示すことになります; Baker and Bellis 1995)。
人間の身体的特徴は異なる物語を語ります。性的二形は比較的小さく(男性は約10-15%大きい)、精巣のサイズは小さく(約40g)、チンパンジーとは対照的に、女性には排卵を示す明らかな兆候がありません。隠れた排卵は、男性から資源を確保する(「食べ物のための性」仮説)、男性による幼児殺害から子孫を保護する、そしてペアボンディングを強化するなど、さまざまな適応的問題を解決するために進化した可能性があります(Moller and Thornhill 1998)。男性による幼児殺害は、多くの哺乳類種、特に霊長類において一般的であり、新しい成体のオスが社会的ヒエラルキーのアルファの地位を獲得した場合に発生します。アルファオスはほとんどの子孫をもうけるため、男性の視点から見ると、彼が父親でないすべての子孫を殺すことは理にかなっています。なぜなら、女性は再び受容可能になるまでの時間が短く、男性がアルファの地位を楽しむ時間は短い可能性が高いからです。この問題が初期の人間において重要であったかどうかは議論の余地がありますが、現代社会や狩猟採集文化の研究は、幼児が生物学的な父親よりも継父によって虐待されたり、殺されたりするリスクがはるかに高いことを示しています(Daly and Wilson 1988; Overpeck et al. 1998; 異なる見解についてはTemrin et al. 2000を参照)。
軽度の性的二形、小さな精巣のサイズ、そして隠れた排卵は、先祖の条件下で人間が一夫一妻制または軽度の多妻制が支配する社会に住んでいたという仮説を支持しています。
男性と女性の親の投資の違いは、性別間の生存比率にも反映されています。霊長類では、全生涯を通じて主に女性に有利ですが、オスが子孫を運ぶ種では、生存の利点の違いが消失するか、逆にオスに有利になることがあります。類人猿では、オスは子孫の世話に比較的少なく貢献するため、メスには大きな生存上の利点があります。チンパンジーでは、女性から男性への生存比率は約1.4、ゴリラでは約1.2ですが、人間では1.05-1.08に減少しており、これは他の類人猿と比較して子供に対する父親の世話の相対的に大きな量を示しています(Allman 1999)。
すでに示したように、これらの多くの生物学的要因は、人間の行動、特に交配相手の選択に影響を与えてきました。広範な研究により、平均して女性は社会的に高い地位の少し年上の男性を交配相手として好むのに対し、男性はウエストとヒップの比率や胸のサイズによって魅力が示される若い女性を好むことが示されています(Buss et al. 1990)。男性と女性の両方が、低い変動非対称性(両側の特徴間の非対称性)を持つパートナーを好むのは、高い変動非対称性がより大きな突然変異負荷、寄生感染、または他の環境ストレス要因と関連していることが示されているためです。したがって、潜在的な交配相手の繁殖「質」は主に無意識的に評価されます。さらに、交配相手の選択は、パートナーの類似性(身体的および心理的)の程度によっても影響を受け、完全にランダムではないことが証拠として示されています(「選択的交配」と呼ばれる; Buston and Emlen 2003)。
妄想的嫉妬やエロトマニアなどの精神病理学的症候群は、人間の交配行動に関与する適応メカニズムの歪みです。
性的選択された人間の特性として現れたメカニズムの極端な変異として、精神病理学的相関は豊富です。潜在的なパートナーとしての自分の特性をアピールすることは、躁病エピソードやヒステリックパーソナリティ障害の重要な側面かもしれません。嫉妬の妄想においては、交配相手の監視が病的に誇張されることが典型的に見られます。おそらく、エロトマニアは他の人に愛されているという妄想的な確信を反映しており、これは最も多く女性に観察されます(詳細については第8章および第19章を参照)。
1.2.2.3 親子間の対立
親と子の間には、個々の子孫に対する親の投資の量と期間に関して対立する利益が存在します。親と子の遺伝的利益には重複がありますが、利益は同一ではありません。子孫の発達のある時点で、親は他の子孫に投資することが利益となります。なぜなら、遺伝子中心の観点から見ると、親は自分の子孫と50%の遺伝子を共有しているため、繁殖成功を最大化するためにより多くの子孫を生産することが理にかなうからです。逆に、子孫にとっては、親が与えようとする以上の投資を確保することが利益となりますが、それは親が特定の子孫を育てるためにかかるコストが2倍以上になるまでのことです。なぜなら、全兄弟も50%の遺伝子を共有しているからです。この時点で、したがって、選択は親(ここでは母親)と子孫の両方に働きかけて投資を終了させるべきです(Trivers 1974)。この単純なルールは、すべての有性生殖を行う種に適用されます。しかし、哺乳類では、親子間の対立は妊娠中から問題になることがあります。例えば、胎児の遺伝子は、母親の利益よりも多くの資源を母親の体から引き出すように選択されることがあります。例えば、母親のインスリンに対する感受性は、胎盤ホルモンによってダウンレギュレーションされ、母親の血糖値が上昇し、胎児へのエネルギー供給が増加します。EPH(浮腫、蛋白尿、高血圧)妊娠中毒症は、この対立の極端(かつ病的)な変異として見ることができます(Schuiling 2000)。EPH妊娠中毒症は、胎児と母親の最適な血糖値の調整が乱れるため、母親に深刻な健康問題を引き起こします(Haig 1993)。
共有される遺伝物質の量のために、親と子孫の間の対立は、親の投資を制限するためのさまざまな適応(親の側)と、親の投資を増加させるための適応(子孫の側)を生み出しました。
哺乳類に特有の別の親子間の対立は、離乳です。通常、幼児は母親が提供するよりも大きく長い投資(ここではミルク供給の観点から)を要求するように選択されます。親子理論はまた、将来の兄弟が全兄弟よりも遺伝的に遠い関係にある場合、離乳の対立が悪化することを予測します。言い換えれば、将来の兄弟が異なるオスによって父親にされる半兄弟である場合、離乳の対立は強化されます。したがって、異なる父親が存在することが例外である可能性が高い人間では、離乳の対立は他の哺乳類種と比較して実際には弱いかもしれません。しかし、母乳を与えられている幼児は、夜間の覚醒の期間を延ばすことで母親の無月経を延長する可能性があると提案されています。これは、父方の印刷遺伝子によって部分的に媒介されるかもしれません(母方の印刷遺伝子は、より統合された睡眠を好む可能性があります; Haig 2014)。
親子間の対立は、子孫の発達のさまざまな段階で発生する可能性があり、子宮内環境(エネルギー供給の量に関して)から思春期や初期の成人期(子孫の交配相手の選択に関して)まで広がります。哺乳類に特有の親子間の対立は離乳です。
同様の観点から、幼児と母親の間で乗せられることや運ばれることに関する対立が生じることがあります。人間の子供は、自分で歩けるようになっても、しばしば運ばれることを要求します(Hrdy 2000)。余談ですが、現代社会では、幼児が母親の腰に運ばれることは伝統的な文化に比べてはるかに少なく、これは股関節形成不全の問題に寄与しています。なぜなら、母親の腰に乗ることが大腿骨の亜脱臼の最良の予防策だからです。
幼児(人間と非人間の両方)は、親からの投資を引き出すためのさまざまな行動を持っています。幼児は親よりもはるかに小さく、経験が少ないため、親が利用可能な資源を支配しているため、幼児は親の投資を増やすために身体的手段ではなく心理的メカニズムを発展させるように選択されてきました(Trivers 1974)。幼児が空腹や危険を感じたときに泣くことは、その一例かもしれません。子孫が特に無力で脆弱な種では、親は幼児のニーズにより迅速に応じるように選択されてきました。これは人間の進化の過程で非常に重要であり、人間の赤ちゃんは出生時に生理的に早産であり、他の霊長類の赤ちゃんと比較して非常に未熟です(第3章を参照)。成長するにつれて、子孫が親の投資を増やすために、より早い発達段階の行動に戻ることが「戦術」として考えられます。この現象は人間の心理学でよく知られており、「退行」と呼ばれています。癇癪は、親に投資を増やさせる別の方法です。自然な状況下では、癇癪の存在は直感に反します。なぜなら、そのような露骨な行動は捕食者を引き寄せ、エネルギー的に高コストだからです。ある意味で、癇癪は親の投資を増やすためのリスクのある戦略と見なすことができ、しばしば非常に成功します。この背後にある進化的論理は、捕食者に襲われる危険を冒す個々の子孫は、遺伝的関連性とすでに投資されたすべての資源の潜在的な喪失のために、親の適応度を減少させる可能性があるということです(Trivers 1974)。
「退行」は、親の投資を増やそうとする子孫の典型的な試みです。
兄弟間の相互作用も親子理論の観点から解釈できます。親はすべての個々の子孫に対して同等に関連しているため、理論的には資源が子孫間で均等に分配されることが親の利益となります。しかし、遺伝的関連性から考えると、個々の子孫が全兄弟に対して利他的な行動をとるのは、兄弟に対する利益が寄付者にかかるコストの2倍以上である場合のみです。言い換えれば、親は兄弟間の利他主義を促進し、自己中心的な行動を抑制するように選択されます(Trivers 1974)。
親子間の対立の他の形態は、子孫の成人(繁殖)役割に関して生じる可能性があります。Triversは、人間の親は子供の交配相手の選択に特に関心を持つべきだと主張しました。なぜなら、親が近親との結婚を促進することが子孫の親族に対する利他主義を高める利益になる可能性があるからです(この傾向は近親相姦の回避によって相殺されます)。一方で、子孫が親の望むよりも早く親の投資を終了させたいと考える状況もあるかもしれません。例えば、親が特定の子孫を繁殖から排除し、その子孫を「巣の手伝い」として保持したいと考える場合です。このような対立は、成長した子供にとって特に解決が難しいかもしれません(Trivers 1974)。
親子間の対立の特別な変種は、子孫の性別に関連しています。通常、自然選択は息子と娘に均等に投資する親を好み、約50/50の性比をもたらします。しかし、良好な条件下では、親がより多くの息子を生産することが利益になる場合があります。なぜなら、息子の繁殖成功は娘のそれよりも大きい可能性があるからです。しかし、悪条件下では、環境条件が女性に対して男性よりも不均等に小さい影響を与えるため、親は娘により多く投資することが好まれるかもしれません(Trivers-Willard仮説; Trivers and Willard 1973)。これらの生物学的に決定されたバイアスは、環境条件の(認識された?)影響の関数として、ヒトや他の哺乳類における子宮内の男性死亡率に深刻な影響を及ぼすことが示されています。一部の哺乳類種は、妊娠中に環境条件に応じて子孫の性別を「決定」し、「間違った」性別の胎児を中絶することさえできます(Kumm et al. 1994)。人間や他の霊長類のように、親の投資が長期にわたる場合、男性または女性の子孫に対する親の行動のバイアスが増加します。ただし、父親の投資の役割は、さまざまな方法でこれらのバイアスを修正する可能性があります。
親子間の対立の特別なケースは、子孫の性別に応じた親の投資の差異に関連しています。良好な環境条件下では、親は男性の子孫を好む傾向があり、これは男性の繁殖潜在能力が女性よりも大きいためです。悪環境条件では、環境条件が女性に与える影響が小さいため、女性の子孫への投資が好まれるかもしれません。
人間における息子と娘への投資の差異の存在は、特に道徳的問題が生物学的原則と混同される場合、大きな論争の対象です。しかし、幼児殺害が霊長類や人間が出生後に親の投資の調整を延期する方法であるという証拠があります。これは、先祖の狩猟採集社会や現代社会においても明らかに起こっています(Hrdy et al. 1994; Brookman and Nolan 2006)。このような場合、ほとんど常に新生児が母親との絆が形成される前に、出生後24時間以内に行われる新生児殺害が行われます(第3章および第21章も参照)。ただし、子供が母親に受け入れられるかどうかは、母親の年齢や生活状況にも依存します。これは、現代社会における母親の中絶の決定にも深刻な影響を与えます(Lycett and Dunbar 1999)。例えば、シングルマザーは、既婚女性よりも妊娠を中絶する可能性が高いです。これは、女性が結婚し、他の子供を持つ将来の見通しに関連しており、若い女性ではかなり高く、年齢とともに低下します。したがって、繁殖期の終わりに近い高齢女性は、たとえ独身であっても妊娠を中絶する可能性が低くなります(Braza 2004)。対照的に、すでに1人または2人の子供を持つ既婚女性では、中絶率が増加します(Lycett and Dunbar 1999)。先天的な出生欠陥への恐れは、意思決定において主要な役割を果たしていないようです。なぜなら、中絶率の急激な増加は、独身の高齢女性には見られないからです。
霊長類や人間は、親の投資に関する決定を出生後まで延期してきました。新生児殺害は、狩猟採集者における出生制御の方法であり、現代社会においても、シングルの十代の母親が新生児を放棄することがあります。
親の投資理論と親子間の対立に関連する問題の組み合わせは、ゲノム印刻に反映されています。父方および母方から受け継がれた遺伝子は、母体が提供する資源を巡って競争する可能性があります(Haig 1993)。この問題は、1.4節の遺伝学で扱われています。
進化生物学の専門用語で表現されているものの、個体間の対立の可能性は、その遺伝的構成の違いに起因しており、人間の心理学や精神病理学の理解に深い影響を与えます。妄想的信念や人格障害の例はすでに言及されています。生物学的に駆動される対立が現れるもう一つの重要な分野は心理療法です。生物学的な対立の領域は、個人にとってほとんど無意識であることを強調する必要があります。原則として意識的な認識にはアクセス可能ですが、おそらく制御するのは難しいでしょう。心理療法的関係は、おそらく主に親子間の対立の投影によって特徴づけられ、特に境界性人格障害(BPD)などの重度の人格障害は、治療中に親の投資を引き起こす行動や拒絶の全範囲を示すことがあります(詳細については第19章および第20章を参照)。同様に、抑圧メカニズムによって特徴づけられる精神病理学的状態(うつ病や不安障害など)は、しばしば「退行」に似た行動を利用し、これが「脅威なし」を強く示し、ケアを最大化することを目的としています(第10章および第11章を参照)。
親子間の対立に関連する行動は、しばしば(無意識に)クライアントとセラピストの関係に転送されます。
1.3 進化心理学と精神病理学の原則
進化心理学の中心的な前提は、人間の心がすべての生物の形態的特徴と同じ生物学的法則に従って進化してきたということです。したがって、自然選択と性的選択は、私たちの思考、感情、行動の仕方を形成してきました(Buss 1999; Rossano 2003)。
進化心理学や関連する学問分野(行動学、社会生物学、行動生態学など)は、人間の心の働きに関するいくつかの誤った前提を反証するのに寄与してきました。広く普及している仮定の一つは、新生児の人間の心は「タブラ・ラサ」(「白紙」)であるという見方でした。私たちは、注意深い観察から、これは事実ではないことを知っています。例えば、新生児は、機能的にはもはや役に立たないが、私たちの霊長類の遺産に深く根ざしたいくつかの原始的な反射を保持しています。モロー反射や把握反射などがその例です。さらに、新生児は通常、母親である主要なケア提供者と非常に洗練された方法で相互作用することができ、これが愛着と絆を強化するのに役立ちます。出生後すぐに、新生児は簡単な顔の動きを模倣することも可能です(Meltzoff and Moore 1977)。赤ちゃんは、幾何学的な形よりも顔に似た形を好み、孤独、 高所、大きな音などに対する生まれつきの恐怖を持っています。これらの能力は、私たちの種特有の環境条件への適応として出現した高度に適応した心理的メカニズムです。
人間の赤ちゃんは、愛着を形成し、生存するのを助けるために進化した一連の心理的メカニズムを持って生まれます。
進化心理学の中心的な前提は、人間の認知、感情、行動が解剖学と本質的に同じ方法で自然選択と性的選択によって形成されてきたということです。
しかし、ほとんどの心理的メカニズムは、経験的な修正に対して非常にオープンです。つまり、これらは「オープンプログラム」(本能に基づく「クローズドプログラム」とは対照的)を表し、人間の生涯を通じて学習経験に応じて反応します。特定の期間は「印刻のような」プロセスにとって重要です(Mayr 2001)。言い換えれば、心理的メカニズムが生理的に機能するためには、先天的な素因(「自然」)が刺激的な環境からの適切な入力(「育成」)と出会うことが不可欠です。そうでなければ、機能不全の方法で発展する可能性があります。例えば、軽度の斜視が幼児で見逃されると、「非優位」の目は決して見ることを学ばず、弱視のままになります。弱視では、影響を受けた目に有機的な障害はありません。むしろ、この障害は、影響を受けた目が適切に刺激されていないという発達上の問題から生じます(Eggers and Blakemore 1978)。類似のことが、言語や共感などの他の多くの進化した心理的メカニズムにも起こる可能性があります。適切な刺激、すなわち環境からの入力が欠如している場合、同様のことが起こる可能性があります。
ほとんどの人間の心理的メカニズムは「オープンプログラム」であり、適切な環境刺激に大きく依存して適切に発展します。したがって、環境からの入力が不足すると機能不全を引き起こす可能性があります。
1.3.1 進化した心理的メカニズムと生物社会的目標
人間の生物社会的・心理的機能は、私たちの進化の過去における選択圧に応じて進化した心理的メカニズムで満ちています(心理的メカニズムという用語は、堅苦しく、学習による修正に対して無防備であることを示唆するため、注意が必要です。しかし、これは進化心理学者によって作られた表現が実際に意図するものではありません)。より具体的には、心理的メカニズムは、適応的な重要性のある問題を解決するために進化によって設計されています(Cosmides et al. 1992)。そのためには、経済的で、信頼性が高く、効率的で、正確である必要があります(Williams 1966)。これらの特性は、特定の領域での機能によって最も維持され、複数の情報源からの情報を統合するためにいくつかの領域一般的なメカニズムが存在する可能性があります。領域特異性は、適応的な問題が進化の過程でかなりの時間にわたって繰り返し存在していた場合に仮定され、個々の繁殖成功に影響を与えます。多くの心理的メカニズムは、特定の情報のクラスに対してのみ機能し、他のモジュールから大きく独立して動作するメンタルモジュールとして概念化できます(Fodor 1983)。モジュールは、迅速な反応のために注意、感情評価、行動反応を集中させるのを助ける意思決定ルールやトリガーアルゴリズムによって活性化されます。
多くの進化した心理的メカニズムは領域特異的であり、つまり、特定の情報のクラスに対してのみ経済的、信頼性が高く、効率的、正確に機能します。進化した心理的メカニズムのモジュールは階層的に組織され、トリガーアルゴリズムによって活性化されます。
モジュールは、その効率を高めるために階層的に組織されていると考えられています(これは医療の意思決定にも重要です; Gigerenzer 1996)。例えば、脅威検出の基本モジュールは、暗闇の中で一人でいるなどの警戒が高まった状況で活性化され、動く形の検出に焦点を当てたアルゴリズムに従うかもしれません。動く形が自己推進型(空気の循環によって生じるランダムな動きとは対照的)として識別されると、動きの方向を調べるアルゴリズムによって高次のモジュールが活性化されるかもしれません。動きの方向が個人に向かっている場合、動いている物体が動物か人間かを評価することに関心を持つアルゴリズムによって別のモジュールがトリガーされるかもしれません。
進化した心理的メカニズムは、異なるタスクを非常に効率的に実行するために設計された一連の別々のデバイスを含むスイスアーミーナイフの多様な機能に例えられています(Cosmides et al. 1992)。ほとんどの比喩的な類似のように、人間の脳をスイスアーミーナイフと比較することは理想的ではありません(例えば、スイスアーミーナイフを意図的に扱う人が必要ですが、複数のタスクの調整された実行は人間の脳自体の別の機能です。したがって、進化の観点からは、人間の心の二元的な概念化の余地はありません)。しかし、この類似は、意思決定や推論のプロセスが抽象的でも論理的でもなく、人間の進化の過程で繰り返される問題に対する選択された回答として最もよく概念化されることを示しています。進化心理学の最も重要な発見の一つは、多く、あるいはほとんどの人間の脳機能が抽象的な問題よりも社会的な問題の解決に偏っているということです(Cosmides 1989)。進化した心理的メカニズムの中には、特に社会的な問題(社会的ルールや道徳的価値など)に関するものは、少なくとも厳密な経済的観点から見ると、非常に非論理的であるものもあります(社会的脳仮説に関する詳細は第2章を参照)。
心理的メカニズムは、適応的な重要性のある問題の解決に偏っており、必ずしも論理を必要としません。
人類にとって社会生活の重要性が際立っていること、そして人間の心に作用してきた多様な選択圧を考慮すると、バイオソーシャル目標の追求が進化した心理的メカニズムの実際のターゲットであることがわかります(Gilbert 1998)。人間は他の生物と同様に、個々の繁殖適応度の最大化に向けて行動が意識的に導かれる「適応度最大化者」として見ることはできないことを強調する必要があります。包括的適応度は振り返って測定されるものであり、将来の繁殖成功は計算不可能です。したがって、進化した心理的メカニズムは、時には進化した「戦略」と呼ばれることもありますが、近接的なレベルで機能します。なぜなら、バイオソーシャル目標を達成することが成功した繁殖の可能性を高めるからです(これは適応度の指標である場合もあれば、そうでない場合もあります)。注目すべきは、「戦略」という言葉は、特定のバイオソーシャル目標を達成する方法についての意識的な認識を含まないということです。
最も一般的な人間のバイオソーシャル目標には、他者からのケアを引き出す動機や行動、主に親族や親しい仲間に対するケアの提供、適切な配偶者の発見、協力的な同盟の形成、可能な限り高い社会的地位の獲得が含まれます(Gilbert 1998)。バイオソーシャル目標は、他者との社会的地位を劣位または優位の観点から比較し、高位および低位の個体との関係を交渉することも目指します。厳密に言えば、バイオソーシャル目標は近接的なレベルの一部であり、個人の生涯の中で柔軟に変化します。例えば、個人は社会的階層の梯子を上ったり下ったりしたり、他者への社会的支援の方向付けにおける努力を変えたりすることができます。したがって、バイオソーシャル目標は、認知、感情、行動の基本的な近接的原因と、遺伝子の作用や個々の育成条件、個体間の遺伝的類似性と非類似性によって引き起こされる問題との間のやや中間的な位置を取ります。男性と女性の個体は、目標の優先順位において顕著に異なり、これは先祖の条件下で男性と女性が担った異なる役割(1.3.2節を参照)や、繁殖に影響を与えた異なる繁殖制約に関連しています(Voland and Dunbar 1995)。したがって、男性にとっては、地位や資源へのアクセスが進化の歴史を通じてより重要であったのに対し、女性は強い親密な絆や感情的な結びつきにより重点を置いていた可能性があります。
人間のバイオソーシャル目標(ケアを引き出すこと、ケアを提供すること、交配、同盟形成など)は、個人の実際の行動を導きます。
進化した心理的メカニズムは、最適に設計されていることはほとんどなく、もしあっても非常に稀です(Gilbert 1998)。これらはしばしば設計の妥協を表し、基本的に失敗や機能不全のリスクを伴います。なぜなら、設計の最適性は進化するには非常にコストがかかる(代謝的に)からです。同様に、相互依存と社会的関係を調整する必要は、心理的機能不全を引き起こす問題を生む可能性があります。これらの問題や精神病理学の他の可能な原因については、第4章およびその後の臨床章でさらに探求されます。
進化した心理的メカニズムは、設計上最適ではないため、機能不全に対して脆弱です。
1.3.2 進化的適応環境
私たち人間を形成するほとんどの心理的メカニズムは、遠い過去の環境条件の中で進化したという考えは、ジョン・ボウルビー(1969年)によって提唱されました。ボウルビーは、人間の精神の多くの側面が、私たちの先祖が狩猟採集者として生活していた地質時代、すなわち更新世(約200万年前から1万年前まで)に起源を持つと主張しました。狩猟採集者の生活に関する研究は、先祖の人間が小さな親族ベースの部族グループに住んでいたことを示唆しています。これらのグループは、社会構造が比較的平等であり、物質的な富の蓄積はほとんどありませんでした。最近の狩猟採集者の社会構造に関する研究では、グループのメンバーのかなりの割合が親族でない可能性があり、成人の兄妹が共に住むことが、両性の帰巣性と分散によって維持されていることが示されています(Hill et al. 2011)。このような社会構造のモデルは、私たちの種の歴史のほとんどの期間、少なくとも解剖学的に現代の人間が出現してから約15万年前から最近の時代まで支配的であったと考えられますが、進化的適応環境(EEA)の単一のシナリオはあまりにも単純すぎると批判されています。したがって、いくつかの環境が人間の身体的および心理的特性の選択に影響を与えたと考えることが妥当です(Foley 1996)。
人間の精神の多くの側面は、狩猟採集者の生活という先祖の環境条件の下で進化しました。人間は、約150人の個人的な知り合いを持つ小さな親密な親族ベースのコミュニティで生活するように適応しています。
最も重要な普遍的な要素の一つは、同じグループのメンバー間の協力が、狩猟や採集から食料を確保するために重要であったことです。これには、女性同士が協力して子供を育てるための相互支援(出産間隔を5-6年から3-4年に短縮することができた; Mace and Sear 1997; Hrdy 2000)や、男性同士が大きな捕食者や競争する人間グループからグループを守るための協力が含まれます(Axelrod and Hamilton 1981)。先祖の人間のコミュニティは、30-40人を超えない規模で構成されており、隣接するグループや拡大家族と合わせて、約150人が互いに個人的に知り合っていました。交易や隣接グループとの戦争は、人類の歴史を通じて一般的であったと考えられます。最近の農業社会のデータによれば、資源を巡る激しい競争の中で、男性の約4人に1人が暴力的な死を遂げることが示されています(Diamond 2012)。その結果、そして現在まで続く人間の悲劇の主な原因として、人間は、グループ内の協力に対する高い傾向があるにもかかわらず、本質的に外部の人々に対して偏見を持ち、内集団に偏った行動をとる傾向があります。つまり、彼らは見知らぬ人を mistrust(不信)し、見知らぬ人が危険にさらされているときや助けを必要としているときに、同じグループのメンバーに比べて共感的でない行動をとる強い傾向があります。この行動は部分的にオキシトシンによって制御されています(Cikara et al. 2011; de Dreu et al. 2012; Shamay-Tsoory et al. 2013; Balliet et al. 2014)。さらに、幼児は生後1年以内に「本能的に」外部の人々に対する偏見を発達させ、特に見知らぬ男性に対してその傾向が強いことが示されています(Eibl-Eibesfeldt 1995)。
グループ内の協力とグループ間の競争(戦争を含む)は、人間の進化の長い期間にわたって支配的でした。
狩猟採集者としての生活の中で最近の歴史において起こった適応を超えて、更新世以前に進化した適応の重要性も考慮する必要があります。例えば、人間とチンパンジーの系統が約500万年前から700万年前に分岐して以来、冷却する気候に対する人間の適応には、二足歩行、脳の拡大と再編成、そして最終的には言語の発展が含まれます。この時期よりもずっと前に、霊長類は主に夜行性で孤独に生活する昆虫食から、昼行性で群れを成して採食する生物へと進化しました。この移行は、立体視、色覚、目と手の協調の進化を伴い、さらに社会的適応の出現を促進しました(Allman 2000)。例えば、眼球が頭の側面から前方に移動することは、狩猟能力や腕の動きの向上に寄与したかもしれませんが、同時に以前はほぼパノラマのような視野を制限し、後方から接近する捕食者を検出する能力を低下させました。これにより、社会的グループの形成が選択的に促進され、したがって社会環境からの適応的問題に対処するための脳のシステムが発展した可能性があります(Allman 2000; より遠い適応は、私たちの階層的に組織された「三重脳」に反映されています。詳細は第2章を参照)。
最近の適応の前に、二眼視は霊長類の社会性の進化に寄与した可能性があります。視野の制限は社会的グループの形成を必要としました。
逆に、いくつかの適応はさらに最近のものであり、初期の人間がアフリカを離れて地球全体に広がった後に出現しました(南極大陸を除く)。おそらく、すべての人間の近い遺伝的関連性(「進化的ボトルネック」仮説と呼ばれる)を示すように、少数の人間のグループが出発しただけでしたが、地元の集団が適応した多くのEEAが存在しました(例えば、ランダムに選ばれた2人の人間は、同じ社会グループの2匹のチンパンジーよりも遺伝的にずっと近い関係にあります)。したがって、EEAは、進化する人間に対して選択圧を生み出すのに十分な期間持続した環境の偶然の統計的な合成として最もよく概念化されるでしょう(Hrdy 2000)。
すべての人間は遺伝的に密接に関連しています。言語や書き言葉の進化に関連するいくつかの適応の修正が進行中であることを示す手がかりがあります。
1.3.3 環境の変化と人間の適応
人間は、物理的、生命的、心理的な性質を問わず、変化する環境に適応し続けています。例えば、数千年前に出現した人間のASPM遺伝子やFoxP2遺伝子ファミリーのアレル変異は、脳の拡大や言語の進化に関連しているかもしれません。ASPM遺伝子のある変異は、5800年前のものであることがわかっており、最初の書かれた文書の発見と偶然の一致ではないと推測されています(Mekel-Brobov et al. 2005)。同様に、脳のサイズを調整する役割を持つ「マイクロセファリン」という遺伝子は、人間の進化の過程で正の選択を受けています(Evans et al. 2004)。一方で、過去5万年の間に人間の脳のサイズと体のサイズは減少しており、これらの変化は「家畜化」の影響に関連している可能性があると考えられています。これは、攻撃性の低下に向けた選択の結果です(Leach et al. 2003)。
さらに最近では、テイ・サックス病の出現が挙げられます。この病気は中世の時代にさかのぼることができ、人々が過密な都市に集まるようになったときに発生しました。この病気は、結核に対する抵抗力を高める遺伝子の選択と関連している可能性があります(Diamond 1988)。これらの最近の例は、最近の人間の適応が神経系に影響を与えなかったわけではないことを示しているかもしれません(第2章も参照)。また、ある研究では、ヨーロッパ系の女性の大規模なコホートにおいて、早期の初潮や遅い閉経といった生殖期間の延長が示唆されています(Byars et al. 2010; Stearns et al. 2010a)。しかし、現代の環境における選択が人間の認知、感情、行動にどのように作用しているかは十分に理解されていません(多くの科学的に根拠のない推測が生じています)。急速に変化する環境が、異なる繁殖成功に影響を与えることはほぼ確実です。例えば、中世のペストの流行は遺伝的ボトルネックを生み出し、免疫機能の変化を選択した可能性があります。また、心理的適応にも影響を与えたかもしれません。逆に、人口の増加や現代の医療の進展による選択圧の変化、そして父親や母親の年齢の増加に関連する新たな突然変異の数が増えることで、遺伝的多様性が大きくなる可能性があります(Greenspan 2013)。
- 急速に変化する環境は、異なる繁殖成功に影響を与えます。
- 中世のペストの流行は遺伝的ボトルネックを生み出した可能性があります。
- 逆に、現代の医療の進展による選択圧の変化は、遺伝的多様性を大きくする要因となるかもしれません。
1.3.4 人間の認知、感情、行動の進化的仮説の検証
進化的な枠組みを人間の心理学に適用することの最も重要な利点の一つは、行動に関するテスト可能な予測を生成できることです(Buss 1999)。進化心理学の仮説をテストするには、トップダウンアプローチを取ることができます。つまり、親の投資理論のような生物の遺伝的関連性に関する中心的な理論からテスト可能な仮説を導き出すことができます。例えば、女性は男性と比較して、社会的に高く評価されるパートナーを求めるように選択されていると仮定することができます。これは、女性が潜在的な子孫に対してより大きな親の投資を行うためです。
対照的に、ボトムアップアプローチでは、実際の行動の観察から始めます。例えば、女性はパートナーの(認識された)社会的地位が低下した場合に離婚することが多い、または男性は女性パートナーの生殖能力が年齢の影響で低下した場合に、より若いパートナーに乗り換えることが多いといった観察です(Betzig 1989)。このような観察は、仮説を確認または反証するためのテスト可能な仮説に変えることができます。図1.1を参照してください。
進化心理学理論から導かれた仮説の実証的検証
進化心理学理論から導かれた仮説の実証的検証は、2つの方法で行うことができます。
- トップダウンアプローチ: 進化理論から直接仮説を導き出す方法。
- ボトムアップアプローチ: 観察に基づいて仮説を立てる方法。
進化のメカニズム
- 自然選択と性的選択(包括的適応度)
- 究極的レベル:
- 互恵的利他主義
- 親の投資と性的選択
- 親子間の対立
- バイオソーシャルゴール(生物社会的目標)を仲介者として
- 協力
- 社会的地位
- パートナー選び
- ケアの提供
- ケアの引き出し
- 近接的レベル:
- 認知、感情、行動
- 神経生理学
- 個々の特異性(アタッチメントを含む)
図1.1
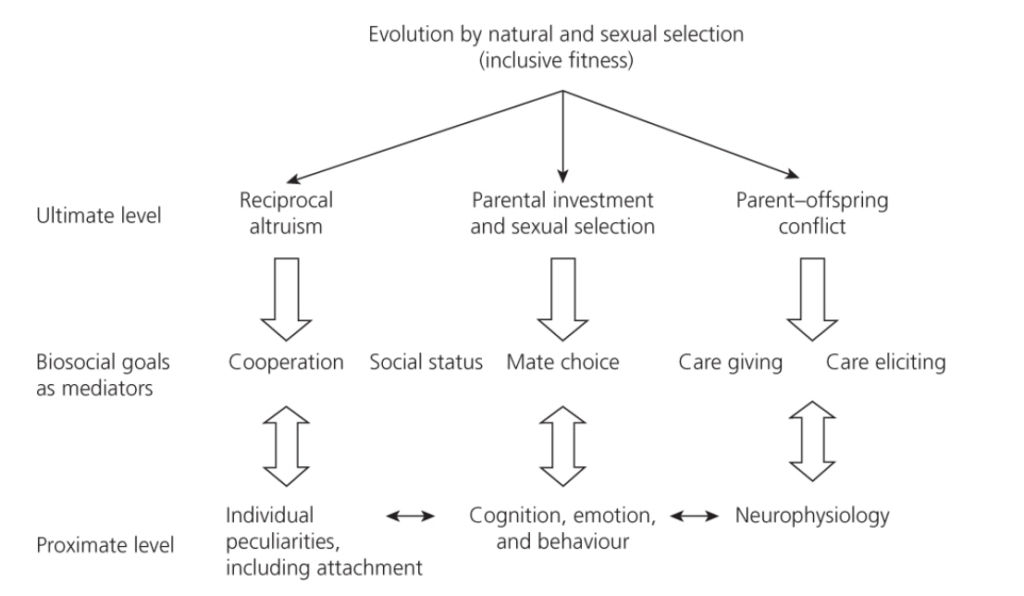
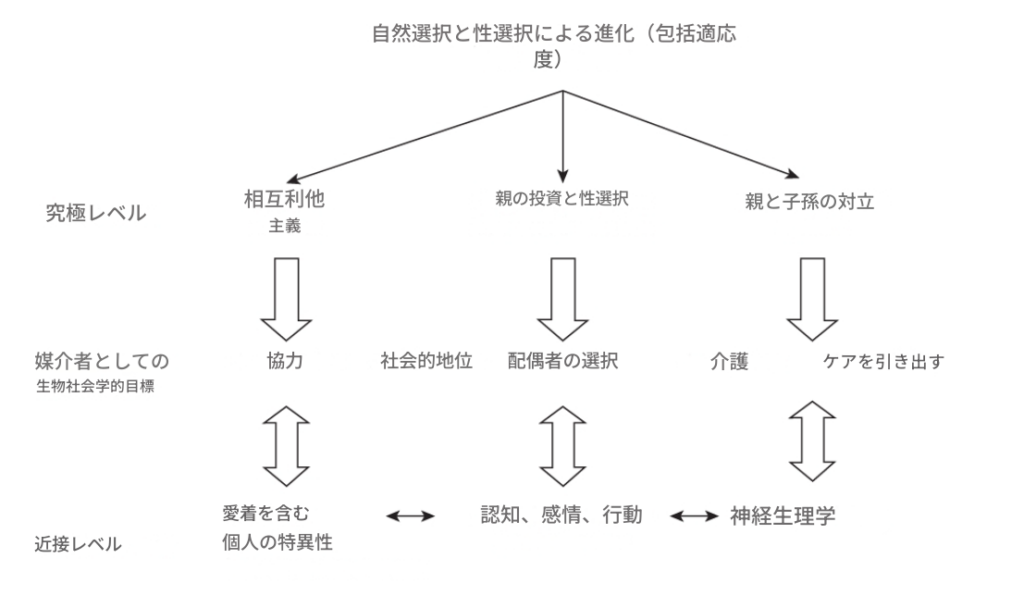
進化心理学と精神病理学に基づく認知、感情、行動の分析のための階層的構造のフレームワーク。
- トップダウン分析: 理論に基づいており、テスト可能な仮説を作成し、そこからテスト可能な予測を導き出すことに焦点を当てています。
- ボトムアップ分析: 観察に基づいています。観察された特性の適応的機能に関する仮説を実験的にテスト可能なデザインに変換し、実証結果に基づいて予測を確認します。
精神病理学においては、ボトムアップアプローチは、精神病理学的な兆候や症状が(仮説的な)ガウス曲線の両端にある変動の極端な例であるという理解のもとで有用です。
(出典: Martin Brüne, “Toward an Integration of Interpersonal and Biological Processes: Evolutionary Psychiatry as an Empirically Testable Framework for Psychiatric Research,” Psychiatry: Interpersonal and Biological Processes, 65 (1), pp. 48-57 c 2002, Taylor & Francis, 許可を得て転載)
進化心理学理論から導かれた仮説の実証的検証
1. 仮説の検証方法
| アプローチ | 説明 |
|---|---|
| トップダウンアプローチ | 進化理論から直接仮説を導き出す方法。テスト可能な仮説を作成し、予測を導く。 |
| ボトムアップアプローチ | 観察に基づいて仮説を立てる方法。観察された特性の適応的機能に関する仮説を実験的にテスト。 |
2. 進化のメカニズム
| レベル | 内容 |
|---|---|
| 究極的レベル | – 自然選択と性的選択(包括的適応度) – 互恵的利他主義 – 親の投資と性的選択 – 親子間の対立 – バイオソーシャルゴール(生物社会的目標) – 協力 – 社会的地位 – パートナー選び – ケアの提供 – ケアの引き出し |
| 近接的レベル | – 認知、感情、行動 – 神経生理学 – 個々の特異性(アタッチメントを含む) |
3. 図1.1: 階層的構造のフレームワーク
進化心理学と精神病理学に基づく認知、感情、行動の分析のための階層的構造のフレームワーク
┌───────────────────────────────────────────────┐
│ トップダウン分析 │
│ 理論に基づいており、テスト可能な仮説を作成し、 │
│ そこからテスト可能な予測を導き出す。 │
└───────────────────────────────────────────────┘
┌───────────────────────────────────────────────┐
│ ボトムアップ分析 │
│ 観察に基づいており、観察された特性の適応的機能に関する │
│ 仮説を実験的にテスト可能なデザインに変換する。 │
└───────────────────────────────────────────────┘4. 精神病理学におけるボトムアップアプローチ
- 精神病理学的な兆候や症状は、(仮説的な)ガウス曲線の両端にある変動の極端な例であるという理解のもとで有用。
1. 仮説の検証方法
ボトムアップアプローチとトップダウンアプローチの両方は、さまざまな証拠源を利用できます。これには以下が含まれます:
- 実験テスト: 認知、感情、行動における性差を調べるための実験。
- アンケート: 社会的期待や望ましさに基づく反応バイアスの問題があるが、情報を収集する手段として使用される(Daly and Wilson 1999)。
- アーカイブデータ: 進化理論なしに収集されたデータ。
- 異文化比較: 人間の普遍性を探るため、また進化したメカニズムの文化特有の修正を評価するために使用される。
- 種間比較: 密接に関連する種の間での比較。
- 精神病理学的兆候や症状: 正常な心理を理解するための有効な情報源として使用できる。
精神病理学の視点
- コンラート・ローレンツは、行動の病理(例:バキューム活動、置換活動)が、行動の生理的機能の分析よりも正常な機能を説明する力が優れていると主張しました(Lorenz 1973)。
- 精神病理学を人間の心理理解の証拠源として考えることは特に有益です。
- しかし、進化心理学ではこの視点はほとんど無視されています(Brüne 2002)。
1.4 遺伝学
生物を成長させるために必要な情報は、個々のゲノムに保存されています。すべての遺伝情報は、デオキシリボ核酸(DNA)というマクロ分子にコードされています。DNAは、以下の4種類の塩基を持つデオキシヌクレオチドから構成されています:
- アデニン(A)
- グアニン(G)
- シトシン(C)
- チミン(T)
ここで、Aは常にTと対になり、GはCと対になります。この「アルファベット」または遺伝コードは、すべての生物で同じです。したがって、動物の基本的な設計(バウプラン)は、非常に長い期間にわたって保存されてきた調節遺伝子によって維持されています(Mayr 2001)。これらの遺伝子は、構造遺伝子とは独立して機能します。
DNAの構成
- 人間のDNAは約35億の塩基対から成り立っています。
- DNAは、コーディングDNAとノンコーディングDNAの混合物です(McGuffin et al. 2005)。
- 人間のゲノムには約30,000の機能的遺伝子が含まれており、これは驚くほど少ない数ですが、動物界では特に例外的ではありません。
- 特に、コーディングDNAの約55%が人間の脳で発現しており、脳が突然変異の主要なターゲットであることを示唆しています。
- 人間のDNAの約97%はノンコーディングであり、その機能は完全には知られていません。しかし、進化は無駄なDNAを排除してきたと考えられています。なぜなら、DNAの生成は目的なしに維持するにはコストが高すぎるからです。
- ノンコーディングDNAの一部は、遺伝子の開始コドン(エクソン)を分離するのに役立つイントロンで構成されています(McGuffin et al. 2005)。言い換えれば、ノンコーディングDNAは遺伝子発現の調節に関与しています(Babbitt et al. 2010; Barrett et al. 2012)。
遺伝子の構造
- 人間のゲノムは約30,000の機能的遺伝子を含むと考えられています。
- 遺伝子は、メッセンジャーRNA(mRNA)に転写されるテンプレートとして機能するDNAの単位です。
- 遺伝子は通常、開始(プロモーター)と停止の領域を持ちます。これらの領域は、DNAからRNAへの転写プロセスの開始と終了に重要です。
- 遺伝子にはイントロンとエクソンが含まれており、イントロンはRNAスプライシングというプロセスによってプライマリ転写物から除去され、完全に成熟したmRNAが生成されます。
- RNAはDNAと似た構造を持っていますが、Tはウラシル(U)に置き換えられています。
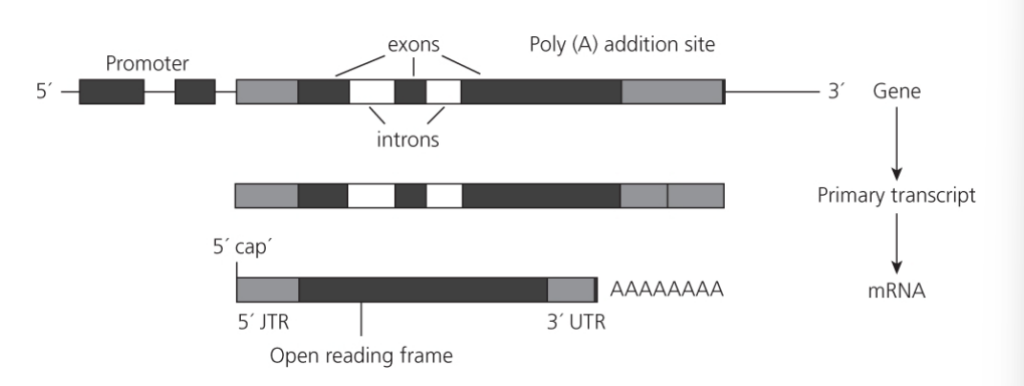
図1.2: メッセンジャーRNA(mRNA)をコードする遺伝子の構造
- 遺伝子の構造:
- 遺伝子は5’末端にプロモーターを含んでいます。このプロモーターはRNAポリメラーゼと結合し、転写を開始します。
- 遺伝子全体の配列がプライマリ転写物に転写されますが、mRNAに表現されるのは遺伝子の一部だけです。
- mRNAに存在する配列(灰色と黒)はエクソンと呼ばれます。
- エクソンの間にある配列はイントロン(白)と呼ばれます。
- 成熟mRNAの生成:
- プライマリ転写物はRNAスプライシングを経てイントロン配列が除去され、3’末端で切断され、ポリA配列が追加されます(ポリアデニル化)。
- 5’末端にはキャップが付けられます。
- 成熟したRNA配列には、ストップコドンで中断されないタンパク質をコードする配列(黒)が含まれています。これをオープンリーディングフレーム(ORF)と呼びます。
- 残りの配列と5’および3’末端は翻訳されない領域(UTR)と呼ばれ、タンパク質をコードしません。
コドンとアミノ酸
- コドンの構造:
- 3つの隣接した塩基がトリプレットまたはコドンを形成し、これがアミノ酸をコードしてタンパク質を構築します。
- 可能なトリプレットの組み合わせは64通り(4^3 = 64)あり、20種類の異なるアミノ酸をコードします。
- したがって、DNA(RNA)コードは冗長性があり、1つのアミノ酸に対して複数のトリプレットの組み合わせが存在します。
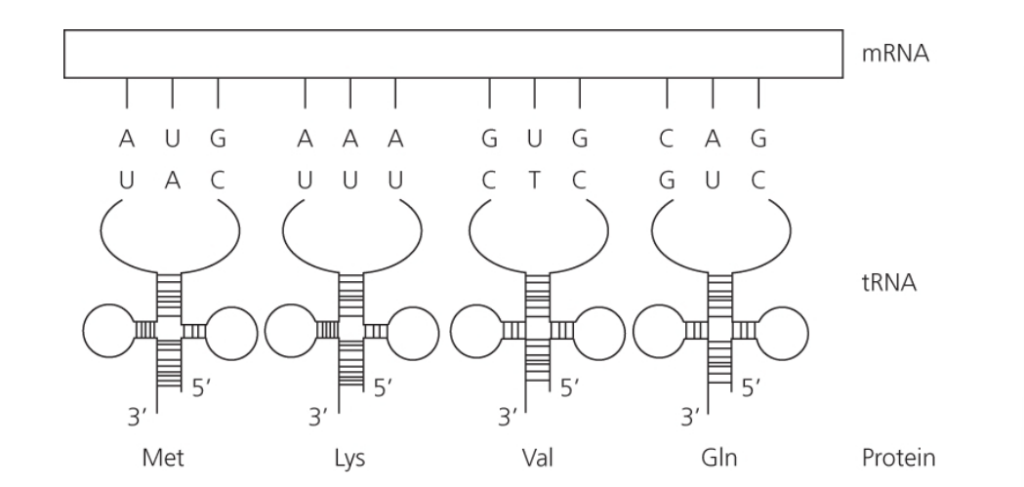
図1.3: 翻訳のプロセス
- tRNAとmRNAの関係:
- 特定のアンチコドンを持つ転移RNA(tRNA)が、mRNAのコドンを塩基対形成によって認識します。
- 各tRNAは、mRNAとリボソームの複合体からなる翻訳機構に特定のアミノ酸を運びます。
- このようにして、コドンの配列がアミノ酸の配列に翻訳されます。
DNAの構造と染色体
- DNAの形状:
- 通常、DNAは二重らせんの形を取り、人間では23対の染色体の基礎を形成しています。そのうち2つは性染色体で、XとYと呼ばれます。
- 染色体はサイズに応じて番号が付けられ、染色体1が最も大きいです。
- 大猿と比較すると、人間の46本の染色体は数が減っていますが、遺伝情報の量は変わりません。
- 染色体の進化:
- 人間とチンパンジーの系統が分かれた約400万〜500万年前に、ゴリラやチンパンジーに存在する共通の祖先の染色体12と13が合体して、人間の染色体2が形成されました(Marks 1992)。
- 性染色体:
- 哺乳類では、雌は2つのX染色体を持ち、雄はXとYの染色体を持ちます。
- Y染色体はX染色体よりもかなり小さいですが、性を決定します。Y染色体を持つと雄が生まれ、持たないと雌が生まれます(対照的に、鳥類ではXXが雄、XYが雌です)。
- 核型:
- 生物の核型は、常染色体と性染色体の数を別々に示します。
- 健康な人間は44 XXまたは44 XYを持っています。
- 異常染色体数:
- 正常な染色体数の変化は「異数性(aneuploidies)」と呼ばれます。
- 最も一般的な例は、ダウン症を引き起こす21トリソミーです。性染色体の異数性は、クラインフェルター症候群(47 XXY)、ターナー症候群(45 X0)などを引き起こすことがあります。
- 異数性は、知的障害や精神病、その他の行動異常のリスクを高めることがよくあります。
- 染色体の再配置:
- 異数性の可能性に加えて、染色体の一部が欠失(削除)、逆方向に再挿入(反転)、複製、または他の染色体に転座されることがあります。
- これらの再配置は、発達に深刻な影響を及ぼす可能性があり、精神的健康にも影響を与えることがあります。
DNAの保存と変異
- DNAの保存性:
- 人間は23対の染色体を持ち、これがDNAを運びます。これらの染色体のうち2つが性染色体です。
- 異数性は染色体の数の変化であり、しばしば行動異常を引き起こします。
- 染色体の構造:
- 染色体は中心部にあるセントロメアで「圧縮」され、長い腕(q)と短い腕(p)を形成します。
- 腕には顕微鏡で見えるバンディングパターンがあり、セントロメアに近いところから番号が付けられます。
- 同じ染色体のペアは「相同染色体」と呼ばれます。この用語は「ホモ接合体」と混同しないように注意が必要です。ホモ接合体は、相同染色体の特定の遺伝子座での情報が同一であることを意味します(そうでなければ、その個体はその遺伝子座でヘテロ接合体です;McGuffin et al. 2005)。
- DNAの類似性:
- 無作為に選ばれた2人の人間のDNAは99%以上同一です。したがって、DNAは非常に保存的な生物学的構造です。
- しかし、理論的には、任意の染色体の任意の遺伝子座で変異が起こる可能性があり、このような変異は「アレル変異」と呼ばれます。
- アレル変異のメカニズム:
- アレル変異が最も一般的に生じるメカニズムは、単一の塩基またはヌクレオチドの置換による突然変異です。
- 特定の遺伝子座で少なくとも2つのアレルの頻度が人口の1%を超える場合、これらの変異体は「多型(polymorphisms)」と呼ばれます。
- 単一ヌクレオチド多型(SNP、便利に「スニップ」と発音されます)は一般的で、平均して1,500塩基ごとに1回発生すると考えられています。
- ほとんどのSNPはノンコーディング領域に存在するため、表現型には影響を与えません。しかし、SNPがコーディング、プロモーター、エンハンサー、またはサイレンサー領域に位置する場合、遺伝子発現に劇的な影響を与える可能性があります。
DNAの変異とその影響
- DNAの同一性:
- 無作為に選ばれた2人の人間のDNAは99%以上同じです。
- 単一ヌクレオチド多型(SNP):
- 人口の1%以上に関連する単一ヌクレオチドの変異を「SNP」と呼びます。
- コーディング領域に位置する場合、1つ以上の塩基の挿入、欠失、複製は、顕著な表現型の影響を及ぼすことがあります。
- コピー数変異(CNV):
- 挿入、欠失、複製、複雑な多部位変異は、最近「コピー数変異(CNV)」または「コピー数多型(CNP)」という用語でまとめられています。
- CNVは以前考えられていたよりも人間のゲノム内で非常に頻繁に発生します。現在までに、約1,400以上のCNV領域が特定されており、これは人間のゲノムの約12%を占めています。
- CNVの機能的意義:
- CNVの機能的意義や進化的起源、特に「孤児遺伝子」と呼ばれる他の系統に機能的な同等物がない遺伝子の出現については、ほとんど理解されていません。
進化的観点からの重要性
- 適応度の向上:
- 進化的観点から、1%の適応度の向上をもたらす変異は、約4,000世代で人口の頻度が0.1%から99.9%に増加します。これは人間の場合、約10万〜12万年で達成されることになります。
- 最近の研究では、人間のゲノムの多くの遺伝子座で加速進化が起こった証拠が増えており、いくつかは正の選択によって急速に固定されたとされています。
- 脳のサイズや機能:
- 人間の系統特異的なゲノムの変化の中には、脳のサイズや機能、免疫機能に関連するものが多く含まれています。これらは進化的なトレードオフであり、人間を障害や病気に対して脆弱にする可能性があります。
COMT遺伝子と精神的健康
- COMT遺伝子の多型:
- カテコール-O-メチルトランスフェラーゼ(COMT)というタンパク質のアミノ酸鎖の位置158におけるホモ接合体バリン/バリン(val/val)多型は、メチオニン/メチオニン(met/met)変異体よりも速く働きます。
- met/met変異体は、シナプス間隙にドーパミンを長く残すため、作業記憶に測定可能な影響を与えます。
- 精神的健康への影響:
- val/valを持つ人は、作業記憶タスクでmet/metを持つ人よりもパフォーマンスが低下し、特に若い頃に大麻を摂取すると統合失調症のリスクが高まると考えられています。
- val/valアレルは病的ではありませんが、他の多くの要因と組み合わさることで、統合失調症のリスクに寄与する可能性があります。
- 情動反応とリスク:
- met/met変異体は、嫌な感情刺激に対して脳の辺縁系や前頭前野、側頭葉での反応が高まることが示されており、ネガティブな気分状態に対する耐性が低く、情動の調整が難しいことを示唆しています。
遺伝子の進化と発現
- 新しい遺伝子の出現:
- 新しい遺伝子は、既存の遺伝子の複製と、祖先の遺伝子の近くへの挿入によって生じることがあります。新しい遺伝子は新しい機能を持つことができ、元の遺伝子はそのまま残ります。
- 遺伝子流動と遺伝的浮動:
- 遺伝子流動は、遺伝子が集団の遺伝子プールに出入りするプロセスです。これは進化において非常に保守的な要因と考えられ、集団が新しい種に進化するのを防ぎます。
- 一方、遺伝的浮動は、小さな集団でアレルがサンプリングエラーによって失われる場合に、そのような影響を及ぼす可能性があります。
エピジェネティクスと遺伝子発現
- エピジェネティクス:
- 遺伝子はエピジェネティックなメカニズムによって異なる発現を示したり、抑制されたりすることがあります。一般的な抑制メカニズムはメチル化で、特にシトシン(C)とグアニン(G)のヌクレオチドが豊富な領域で発生します。
- CpGアイランドは、約50%の遺伝子の片方の端に見られ、その60〜70%がメチル化されています。
- メチル化の役割:
- DNAメチル化は、正常な細胞分化において重要なメカニズムですが、腫瘍抑制遺伝子を抑制することによって癌の発生にも関与していることが示されています。
- 遺伝子のサイレンシング:
- RNAがホモログDNA配列と相互作用することによって遺伝子サイレンシングを誘導することが発見され、これは従来の遺伝学の原則に反します。
- 動物研究では、非遺伝的な継承が生殖細胞、ホルモン、サイトカイン、エピジェネティックな修飾を介して、複数世代にわたって表現型に影響を与える可能性が示されています。
エピジェネティクスと遺伝子サイレンシング
- メチル化と遺伝子サイレンシング:
- メチル化は、X染色体の1本を不活性化することや、ゲノムインプリンティングと呼ばれるプロセスに関与しています。
- ゲノムインプリンティングは、男性または女性のDNAのメチル化の違いから生じます。
- 親の遺伝子の「利害関係」:
- 胎児における母親と父親の遺伝子の「利害関係」が異なるため、進化的観点からは、母親から伝わる遺伝子と父親から伝わる遺伝子の間で、母親の投資の量に対する対立があると考えられます。
- 例えば、父親から受け継いだ遺伝子は、母親にとって最適な資源よりも多くの資源を引き出そうとする可能性があります。一方、母親から受け継いだ遺伝子は、母親の最適な資源に近い、またはそれ以下の資源を要求するように選択されるかもしれません。
- インプリンティング遺伝子の影響:
- インプリンティング遺伝子はゲノムのわずか1%を占めていますが、成長調整に関与しているため、重要な影響を及ぼすことがあります。
- インプリンティング遺伝子のターゲット組織は、胎盤と脳です。これらは、部分的に異なる適応度の利害関係を持つ遺伝的に関連する個体間の資源移動において重要な器官です。
ゲノムインプリンティングの例
- プラダー・ウィリー症候群とアンジェルマン症候群:
- プラダー・ウィリー症候群とアンジェルマン症候群は、ゲノムインプリンティングによる対照的な効果を示す2つの例です。
- 15番染色体の長腕にある特定のDNAの部分が親のいずれかで欠失したり、片親からのみ遺伝子が伝わった場合、母親または父親の遺伝子のみが発現します。
- プラダー・ウィリー症候群: 母親からの遺伝子のみが発現し、赤ちゃんは吸引力が弱く、泣き声も弱い。
- アンジェルマン症候群: 父親からの遺伝子が2つとも伝わると発生し、赤ちゃんは強い吸引反応を示すが、協調性が欠けています。
精神的健康とゲノムインプリンティング
- 自閉症と統合失調症:
- 最近、ゲノムインプリンティングは自閉症や統合失調症に関連して議論されています。これらは、社会情報処理に関連する機能不全の対照的な現れとされています。
- 母親からのインプリンティング遺伝子が、社会的認知に関与する特定の脳領域の成長不足を引き起こす一方で、父親からのインプリンティングは脳の過成長に関与している可能性があります。
- 脳の非対称性と機能的相関:
- これらの極端な変異は、脳の非対称性の低下または増加、接続パターンや機能的相関の逆のパターンとして反映されます。
エピジェネティクスと精神的障害
- エピジェネティックメカニズム:
- エピジェネティクスは、最近、依存症、うつ病、統合失調症などのいくつかの精神的障害に関連して議論されています。
- 特に、慢性的なストレスや早期のストレスフルな生活イベントは、遺伝子サイレンシングに寄与する重要な要因です。
- 母親の育児とメチル化:
- 動物モデルの研究では、母親の育児の強度がグルココルチコイド遺伝子のプロモーターのメチル化に影響を与えることが示されています。
- 早期の逆境(例えば、児童虐待や不適切な扱い)は、ストレス調整に関与する遺伝子のメチル化の増加と関連しています。
- ストレスと生理的反応:
- 慢性的なストレスは、個体の「アロスタティック負荷」に影響を与え、これは自律神経系、視床下部-下垂体-副腎(HPA)軸の反応、体脂肪の蓄積、免疫機能など、さまざまなレベルでのストレス反応に関係しています。
- 薬理学的逆転:
- 動物モデルの研究は、これらの通常は長期的な影響が薬理学的に逆転可能であることを示唆しています。これは、精神的および心身の状態に関する今後の研究でより注目されることが期待されます。
幼少期の逆境と遺伝子メチル化
- 幼少期の逆境:
- 幼少期の逆境は、ストレス調整に関与する遺伝子のメチル化の増加と関連しています。
- 慢性的なストレス:
- 慢性的なストレスは、個体の「アロスタティック負荷」に影響を与える可能性があります。アロスタティック負荷とは、体のストレス反応に関するもので、以下のような要素が含まれます:
- 自律神経系の反応
- 視床下部-下垂体-副腎(HPA)軸の反応
- 体脂肪の蓄積
- 免疫機能
期待(Anticipation)と遺伝性疾患
- 期待の概念:
- 非常に稀な疾患、特に常染色体優性遺伝病(例:ハンチントン病)では、疾患の重症度が世代を重ねるごとに増加し、発症年齢が若くなることが観察されています。この現象は「期待(Anticipation)」と呼ばれています。
- 期待は、親から子への遺伝の過程で、繰り返しの三塩基要素の数が増加することに関連しています。
- 精神的障害との関連:
- 期待が他の精神的障害にどの程度関与するかは議論の余地がありますが、統合失調症、双極性障害、不安障害において期待が関与する可能性があるとされています。
遺伝子の相互作用と多因子性
- エピスタシス:
- 遺伝子はさまざまな方法で相互作用し、これを「エピスタシス」と呼びます。
- 多因子遺伝:
- 特性は複数の遺伝子によって制御されることが多く、これを「多因子遺伝」と呼びます。
- 逆に、複数の特性に影響を与える遺伝子は「多効性(pleiotropic)」と呼ばれます。
精神的障害に対する遺伝的寄与の特定
- 遺伝的寄与の理解:
- SNP(単一ヌクレオチド多型)、複雑な遺伝子相互作用、加算的遺伝子効果、遺伝子サイレンシングやインプリンティング、多効性効果、期待が精神的障害にどのように寄与するかは、まだ理解が始まったばかりです。
- 遺伝的素因と環境要因:
- 個人の遺伝的構成が精神的障害に対する脆弱性と関連していることは疑いありませんが、遺伝的要因と環境的要因を区別することは非常に難しいです。
- ほとんどの主要な精神的障害は、メンデル的な単純な優性-劣性の遺伝パターンに従わないためです。
- 表現型の確率と浸透率:
- 遺伝子は変動的に発現することがあり、特定の遺伝子型に対する特定の表現型の確率を示す「浸透率」は不完全である場合があります。
- 精神的障害の症候群は、通常の状態から明確に区別されるのではなく、量的または連続的に分布しています。
遺伝の複雑さとモデル
- 複雑なモデル:
- これらの要因は、数学的に複雑で多因子のリスク閾値モデルを必要とします。これらのモデルは、ポリジェニックな遺伝と複数の環境要因によって与えられる連続的なリスク分布を仮定しています。
- 遺伝率の定義:
- 遺伝率は、表現型の変動のうち、遺伝的変動によって説明される割合を指します。厳密には、遺伝率は集団内の遺伝的変動を指し、個人レベルでは有効な概念ではありません。
- 環境要因の考慮:
- 数学的モデルは、共有環境要因と非共有環境要因の推定を考慮する必要があります。
まとめ
- 精神的障害の遺伝的背景:
- 精神的障害の遺伝的背景は多因子性であり、遺伝率は遺伝的変動によって説明される表現型の変動の割合として定義されます。
精神的障害に対する遺伝的寄与の推定方法
家族、双子、養子研究
- 家族研究:
- 精神的障害の遺伝的寄与を推定する最も簡単な方法は、家族、双子、養子の研究です。
- 家族研究では、障害を持つ親族の数を調べ、その親族全体の数で割ることで、障害の生涯有病率を計算します。
- ただし、実際の生涯リスクは、すべての親族がリスクの期間を経験しているわけではないため、実際のリスクよりも低くなります。
- また、家族研究は世代間の生涯リスクを比較するのには適していません。
- 双子研究:
- 双子研究、特に一卵性双生児(MZ)と二卵性双生児(DZ)の比較は、より情報を提供します。
- 一卵性双生児は100%の遺伝物質を共有し、二卵性双生児は50%を共有します。
- MZとDZは、妊娠中、子供時代、思春期において、環境リスク要因を同程度に共有しますが、妊娠中の違いは共通の絨毛を共有しないことから生じる場合があります。
- 養子研究:
- 養子研究は、遺伝的要因と環境的要因の違いを明らかにするのに役立ちます。
- 例えば、遺伝的に影響を受けた親が養子に出した子供を、影響を受けていない家族からの養子と比較することができます。
遺伝と環境の相互作用
- 遺伝と環境の相互作用:
- 遺伝と環境の相互作用や相関関係は、解明し、定量化するのが難しいです。
- 受動的遺伝-環境相関: 子供は親から遺伝子を受け継ぐだけでなく、特定の遺伝子を持つ親がその遺伝子に基づいた環境を作ることがあります。
- 例: 不安な親は、リスクの少ない環境を子供に提供しようとするかもしれません。
- ADHDの親を持つ子供は、衝動的な反応を促進する家庭環境で育つ可能性があります。
- 能動的遺伝-環境相関:
- 子供が自分の遺伝的特性に最も適した環境条件を好むことを指します。
- 例: ADHDの子供は、不安な子供よりもリスクのある状況を求めることが多いかもしれません。
- 遺伝-環境相互作用の定義:
- 遺伝-環境相互作用は、遺伝的効果、環境的効果、遺伝-環境相関の合計として定義されます。
リンケージ解析と関連解析
リンケージ解析と関連解析の概要
- リンケージ解析:
- リンケージ解析は、特定の遺伝子やSNP(単一ヌクレオチド多型)を検出するための方法です。
- DNAの指数関数的な複製を利用して、ポリメラーゼ連鎖反応(PCR)を使用します。
- リンケージ解析は、染色体上の2つの点の間で交差(組換え)が起こる可能性に関連しています。2つの点が近いほど、交差が起こる可能性は低くなります。
- 関連解析:
- 関連解析は、遺伝的に無関係なサンプルに適用できます。
- 最も単純な比較は、障害を持つ個人とマッチした対照群の比較です。
リンケージ不均衡(LD)
- リンケージ不均衡(LD):
- LDは、異なる遺伝子座にある2つのアレルがどれだけ密接に関連しているかを測定する指標です。
- 高いLDは、2つのアレルが一緒に伝達されることを示唆しますが、同じ距離の遺伝子座間での変動が大きいため、関連性は短い距離でのみ意味があります。
精神的障害の遺伝学
- 精神的障害の定量的遺伝学:
- 複雑な精神的障害の定量的遺伝学は、かなりの進展があるにもかかわらず、まだ初期段階にあります。
- これは、異なる遺伝子座にある複数のアレルの複雑な相互作用に起因しています。そのため、研究結果の再現がしばしば失敗し、個々の発見が全体の集団リスクを説明する力が弱いです。
- 複数の遺伝子の関与:
- 複雑な特性は単一の遺伝子によって遺伝するのではなく、複数の遺伝子によって遺伝します。各遺伝子は、実際の表現型に小さな変動を寄与します(非遺伝的要因も含む)。
- 複数の遺伝子システムにおける遺伝子は「定量的形質座(QTL)」と呼ばれますが、各QTLが特性の変動にどの程度寄与しているかはほとんど知られていません。
サンプルの比較可能性とエンドフェノタイプ
- 異なる民族背景のサンプルの比較:
- 異なる民族背景を持つサンプルの比較可能性にも問題があります。
- 精神的障害の診断基準があまりにも曖昧であるため、特定の表現型に寄与する遺伝子を特定するのが難しいです。
- エンドフェノタイプ:
- エンドフェノタイプは、障害の完全な表現型の変動に比べて遺伝的に複雑さが少ないと考えられています。
- 理想的には、エンドフェノタイプは遺伝可能であり、複数の個人が影響を受ける家族内で病気と共に分離します。
- エンドフェノタイプは、一般集団よりも影響を受けていない親族に高い頻度で見られ、病気の活動に応じて変動しない状態独立性を持っています。
エンドフェノタイプと精神的障害の遺伝的要因
エンドフェノタイプの研究
- エンドフェノタイプ:
- エンドフェノタイプは、精神的障害の完全な表現型の変動を研究するよりも有望かもしれません。
- エンドフェノタイプは、理想的には遺伝可能で、障害の活動とは独立しており、影響を受けていない親族にも見られる生物学的または行動的なマーカーによって特徴づけられます。
動物モデルの利用
- 動物モデル:
- 人間のDNAをマウスなどの動物のDNAに組み込むことで、行動や生理的な相関関係を観察できることが期待されています。
精神的障害の遺伝的要因
- 遺伝的要因の複雑さ:
- 多くの精神的障害の感受性遺伝子を探すことが難しい理由の一つは、人間の行動が非常に複雑で、数百、あるいは数千の遺伝子によって影響を受けるためです。
- 進化的視点:
- 精神的障害が一般集団において非常に一般的である理由には、進化的な説明が必要です。
- 高い有病率は、精神的障害を引き起こす遺伝子が選択によって排除されていることを示唆するかもしれません。
- 隠れた適応的利点:
- 精神的障害を引き起こす遺伝子が、進化の過程で何らかの隠れた適応的利点を持っていた可能性があります。
- 例えば、遺伝子が早期の生活で適応的な利点を持ち、後に有害な影響を及ぼすことがあるかもしれません。
バランス選択と頻度依存選択
- バランス選択:
- バランス選択は、2つのアレルが進化の過程でフィットネスにおいて等しい重要性を持つことを前提としています。
- 頻度依存選択:
- 特定のアレルが少なくなると、そのアレルによって与えられるフィットネスが増加する場合に発生します。
- これは、特定の特性が低頻度で維持される理由を説明するのに役立ちます。
精神的障害の遺伝的要因の複雑さ
- アレルの累積効果:
- 精神的障害が選択によって排除されない理由の一つは、アレルの累積的な変異が個人を精神的障害にかかりやすくすることです。
- 特定の遺伝子座での稀なアレルが、他の遺伝子座でのアレルと組み合わさることで機能不全を引き起こす可能性があります。
- 「分水嶺モデル」:
- 特定の遺伝子座での複数の不利なSNP(単一ヌクレオチド多型)が組み合わさることで機能不全を引き起こすことを示しています。
- COMT遺伝子の例:
- COMT遺伝子のval/valバリアントは、前頭前野でのドーパミンの回転を高め、これが作業記憶機能に影響を与える可能性があります。
精神的障害の理解
- 遺伝子と環境の相互作用:
- 遺伝子は環境要因と密接に相互作用することで特定の表現型を生み出します。
- 精神的障害は、各遺伝子が表現型の変動に小さな影響を与えるため、選択によって排除されにくいと考えられています。
1.4.2 遺伝子と環境の相互作用および環境に対する感受性の違い
ジアセシス-ストレスモデル
- ジアセシス-ストレスモデル:
- 個人が遺伝的な要因によって精神的な病気を発症するリスクが高まるという考え方です。
- 特に、逆境のある環境にさらされたときにこのリスクが高まります。
- このモデルによれば、「脆弱性遺伝子」を持たない人は逆境に対してあまり影響を受けず、「レジリエント(回復力がある)」と見なされます。
遺伝子と環境の相互作用
- 遺伝子の影響:
- 遺伝子は、逆境のある環境と関連して病気の発症を促進する可能性があります。
- 逆に、脆弱性遺伝子を持たない人は逆境に対して影響を受けにくいです。
- 具体例:
- MAO-A酵素の低活性バリアント:
- この遺伝子を持つ人は、逆境のある環境で育つと反社会的人格障害を発症する可能性が高いです。
- 高活性アレルを持つ人と比べて、逆境の影響を受けやすいです。
- セロトニントランスポーター遺伝子(5-HTTLPR):
- s-アレルを持つ人は、ストレスの多い生活イベントがあると抑うつ症状を発症しやすいです。
- ドーパミン受容体D4遺伝子(DRD4):
- 7回繰り返しのバリアントを持つ子供は、母親が子供のニーズに無関心な場合、外向的な問題やADHDのリスクが高まります。
- 母親のバッファリング:
- 逆境がない場合、遺伝的な脆弱性を補うことができるとされています。
- 遺伝的保護:
- 一部の遺伝的変異は、厳しい逆境があっても精神的な病気の発症を防ぐことがあります。
- 例えば、コルチコトロピン放出ホルモン受容体の変異は、子供の頃に虐待を受けた人が抑うつ症状を発症するのを防ぐ可能性があります。
遺伝子と環境の相互作用の重要性
- 知識の進展:
- 精神的障害に対する遺伝的要因の研究は、遺伝子と環境の相互作用についての理解を大いに深めました。
ジアセシス-ストレスモデルの問題点
- 最近の正の選択:
- ジアセシス-ストレスモデルが直面している問題の一つは、多くの「脆弱性遺伝子」が人間の進化の中で最近正の選択を受けているという観察です。
- これは矛盾しています。なぜなら、自然選択が逆境に対する脆弱性を高めるアレルを選好したとは考えにくいからです。
- 進化的視点:
- 進化の過程で、これらの遺伝子が生殖的なフィットネスに対して未発見または見落とされた有益な効果を持っている可能性が高いことを示唆しています。
遺伝子と環境の相互作用:差異的感受性
DRD4遺伝子の7回繰り返しバリアント
- DRD4遺伝子:
- 7回繰り返しのバリアントは、約5万年前に人間の集団で出現した比較的新しいアレルです。
- 4回繰り返しのバリアントは、祖先の形です(Ding et al. 2002)。
- この長いバリアントは、定住している集団よりも移動する集団に多く見られることが知られています(Chen et al. 1999)。
- 新奇追求:
- 7回繰り返しのアレルは、「新奇追求」という性格特性と関連しています。
- 新奇追求は、人間の歴史において特に移動する集団にとって生殖的な利点をもたらした可能性があります(Reist et al. 2007; Matthews and Butler 2011)。
HTTLPR遺伝子のs-アレル
- s-アレルの影響:
- HTTLPR遺伝子のs-アレルを持つことは、逆境のある出来事と関連して抑うつ症状を引き起こす可能性があります。
- しかし、この同じ変異は、いくつかの領域で優れた認知能力や社会的適応を高めることとも関連しています。
- これは、扁桃体の活動が増加することによって部分的に媒介される可能性があります(Canli and Lesch 2007; Homberg and Lesch 2011)。
バランス多型
- バランス多型の例:
- バランス多型とは、あるSNP(単一ヌクレオチド多型)が一つの領域で不利な影響を及ぼす一方で、別の領域で有利な影響を補うことを指します。
- これにより、特定のSNPが一般集団で頻繁に見られる理由や、選択されない理由を説明できます。
環境の質と遺伝子の影響
- 環境の質の評価:
- 環境刺激の質を評価する際に、いくつかの研究者は方法論的な問題を指摘しています(Belsky et al. 2009; Ellis et al. 2011a)。
- 逆境がないことを「良い」環境の終点と見なすのは不十分だと主張しています。
- また、障害がないことを心理的機能の「良い」終点と単純に比較することもできません。
- 環境の変動の測定:
- 環境の変動を適切に測定しないと、環境の影響に対する感受性の違いを見逃す可能性があります。
- 特に、初期の家庭環境が重要な役割を果たします(Belsky et al. 2009; Ellis et al. 2011a)。
遺伝子の役割
- 遺伝子の二面性:
- 逆境のある環境で病理に対する脆弱性を高める遺伝子が、支援的または豊かな環境では有益に働く可能性があるとされています(Belsky et al. 2009)。
- 遺伝的変異によってもたらされる感受性や可塑性について話す方が、脆弱性に一方的に焦点を当てるよりも正確であるという証拠があります(Belsky and Pluess 2009)。
差異的感受性
- 差異的感受性:
- 早期の逆境と関連して病理に対する脆弱性を高める遺伝的変異が、環境がより支援的な場合には有益な効果を持つことがあります。
- これを「差異的感受性」と呼びます。
遺伝子と環境の相互作用:差異的感受性
MAO-A遺伝子と逆境
- MAO-A遺伝子の低活性バリアント:
- この遺伝子の低活性バリアントは、子供が支援的な環境で育つと、反社会的人格障害の発症率が平均よりも低いことが示されています(Widom and Brzustowicz 2006)。
- ただし、他の研究では遺伝的変異と環境が独立したリスクをもたらすことも示唆されています(Reif et al. 2007)。
5-HTTLPR遺伝子と抑うつ
- 5-HTTLPR遺伝子のs-アレル:
- s-アレルを持つことは、良好な環境条件下では抑うつのリスクを低下させることがわかっています(Taylor et al. 2006)。
文化と遺伝子の関係
- 文化の影響:
- 個人主義的な文化と集団主義的な文化の間で、セロトニントランスポーター遺伝子の遺伝的変異が不安障害や抑うつの発症率の違いに関連しているようです。
- 集団主義を重視する文化では、s-アレルの頻度が高い一方で、不安障害や抑うつの発症率は低いです(Chiao and Blizinsky 2010)。
DRD4遺伝子とADHD
- DRD4遺伝子の7回繰り返しバリアント:
- この遺伝子の7回繰り返しバリアントを持つ子供は、母親が子供の感情的なニーズに応じる場合、ADHDや外向的な問題を発症する率が平均よりも低いです(Bakermans-Kranenburg and van Ijzendoorn 2006)。
ストレスと親の応答性
- 親の応答性:
- COMTvalおよびDRD4 7回繰り返しアレルを持つ大人は、ストレスレベルが低いときに子供のニーズに最もよく応じますが、ストレスレベルが高いときには応答性が平均よりも低くなります(van Ijzendoorn et al. 2008)。
遺伝子の役割と環境への感受性
- ドーパミンとセロトニンの役割:
- これらの研究結果は、ドーパミンとセロトニンの代謝に関与する遺伝子の変異が、環境条件に対する差異的感受性に重要な役割を果たすことを示しています。
- これは、報酬や罰への応答性、ストレス調整によって媒介される可能性があります(Bakermans-Kranenburg and van Ijzendoorn 2007; Ellis et al. 2011a)。
プラスチシティ遺伝子
- プラスチシティ遺伝子の効果:
- プラスチシティ遺伝子は加算的な効果を持つ可能性があり、プラスチシティアレルの数が増えると環境への感受性が高まることがあります(Belsky and Beaver 2011)。
新しい視点の重要性
- 新しい視点の影響:
- 遺伝子と環境の相互作用に関するこの新しい視点は、精神的な病状に関して非常に重要です。
- 差異的感受性は、他の臓器で発現する遺伝子とは異なり、中枢神経系で発現する遺伝子に特有である可能性があります。
誤解と治療的アプローチ
- 誤解のリスク:
- 高い感受性を持つ個人を逆境から守ることは誤った結論です。
- 代わりに、敏感な個人には早期から応答的な社会環境を提供することが治療上の重要な課題です(Ellis et al. 2011a)。
- これは、心理的問題の予防や早期発見に関わる医療提供者にとって大きな課題です。
遺伝的決定論と「あるべき」と「ある」の混同についての考察
遺伝的決定論の誤解
- 遺伝的決定論:
- 人間の行動や精神病を理解するための進化的視点に対する批判の中で、「遺伝的決定論」という誤解がよく見られます。
- 遺伝子が行動や性格を引き起こす、または決定するという考え方です。
- 例えば、「遺伝子Xが障害Yを引き起こす」という表現は、遺伝子の働きを過度に単純化したものです。
遺伝子の役割
- 遺伝子の機能:
- 遺伝子は、複雑な生物を成長させるために必要な基本的な要素です。
- 遺伝子はタンパク質の合成をコードし、タンパク質は成長や組織の分化、神経伝達などの複雑な相互作用を調整します。
- しかし、遺伝子は単独では機能しません。環境条件が特定の範囲内にある場合にのみ、効果的にタンパク質合成を促進できます。
- 環境条件:
- 環境条件には、温度、pH値、アミノ酸合成のための原材料の供給、親のケアや育成、環境の刺激などが含まれます。
- 生物が成長するためには、遺伝子(自然)と環境(育成)の両方が必要です。
認知、感情、行動の理解
- 自然と育成の議論:
- 人間の認知、感情、行動(精神病を含む)に関する自然と育成の議論は混乱を招くことがあります。
- どちらか一方だけでは特定の表現型を説明できません(Schaffner 2001)。
- 一部の精神障害は遺伝的要因が強く、他の障害は環境条件に依存することがあります。
- しかし、精神的に病んでいる患者の大多数において、遺伝的要因と環境的要因が実際の表現型にどの程度寄与しているかは不明です。
進化的視点の重要性
- 進化的視点:
- 進化的視点は、特定の行動傾向が修正されないことを示唆するものではありません。
- 人間の進化は、非常に高い柔軟性を与えています。
- これは、長い幼少期を必要とし、再生産を遅らせる一方で、環境からの影響に非常に依存するため、人間を機能不全に陥りやすくします。
自然主義的誤謬の警告
- 自然主義的誤謬:
- 自然主義的誤謬とは、生物学的事実が道徳的命令に変わるという誤った考え方を指します。
- 生物学的に説明できる行動傾向の存在(「ある」)を、道徳の「あるべき」と混同してはいけません。
- 例えば、男性が複数のパートナーと交尾する生物学的傾向があるからといって、婚外関係が道徳的に正当化されるわけではありません。
過去の教訓
- 過去の誤り:
- 精神科医が自然主義的誤謬に基づいて行動した歴史的な事例があります。
- 例えば、ナチス政権下では、精神科医が遺伝的退化を懸念し、負の優生学(遺伝的に劣ると見なされた人々の繁殖を防ぐ)や正の優生学(繁殖実験)を主張しました。
- これにより、多くの国で強制的な不妊手術法が導入され、精神的に病んでいる多くの人々の安楽死の科学的根拠となりました。
現代の科学と進化
- 現代の科学:
- 現在、遺伝子研究を人間の種を改善するために利用すべきだと主張する科学者もいますが、進化は方向性のあるプロセスではありません。
- 個々の性格の多様性が人間の生活を面白くしています。
精神医学の責任
- 精神医学の役割:
- 精神医学は、時代の流れに影響を受けやすい医療分野です。
- したがって、精神医学に関わるすべての人は、私たちの医療分野の背後にある科学的および非科学的な概念を批判的に見直す義務があります。
- 進化的な考え方は精神病理学を理解する上で重要ですが、他の科学的概念と同様に誤用される可能性があります。
- 1.2.2.3 親子間の対立
- 1.3 進化心理学と精神病理学の原則
- 1.3.1 進化した心理的メカニズムと生物社会的目標
- 1.3.2 進化的適応環境
- 1.3.3 環境の変化と人間の適応
- 1.3.4 人間の認知、感情、行動の進化的仮説の検証
- 進化心理学理論から導かれた仮説の実証的検証
- 進化心理学理論から導かれた仮説の実証的検証
- 1. 仮説の検証方法
- 1.4 遺伝学
- 図1.2: メッセンジャーRNA(mRNA)をコードする遺伝子の構造
- コドンとアミノ酸
- 図1.3: 翻訳のプロセス
- DNAの構造と染色体
- DNAの保存と変異
- DNAの変異とその影響
- 進化的観点からの重要性
- COMT遺伝子と精神的健康
- 遺伝子の進化と発現
- エピジェネティクスと遺伝子発現
- エピジェネティクスと遺伝子サイレンシング
- ゲノムインプリンティングの例
- 精神的健康とゲノムインプリンティング
- エピジェネティクスと精神的障害
- 幼少期の逆境と遺伝子メチル化
- 期待(Anticipation)と遺伝性疾患
- 遺伝子の相互作用と多因子性
- 精神的障害に対する遺伝的寄与の特定
- 遺伝の複雑さとモデル
- まとめ
- 精神的障害に対する遺伝的寄与の推定方法
- リンケージ解析と関連解析
- エンドフェノタイプと精神的障害の遺伝的要因
- 1.4.2 遺伝子と環境の相互作用および環境に対する感受性の違い
- 遺伝子と環境の相互作用:差異的感受性
- 遺伝子と環境の相互作用:差異的感受性
- 遺伝的決定論と「あるべき」と「ある」の混同についての考察
- 第2章
- 人間の脳:解剖学、進化、機能
- 2.1 人間の脳への序論
- 脳の進化と機能
- 脳の進化と機能
- 図:人間の脳の構造と機能
- 大きな脳の生物学的問題
- 2.2 人間の脳の大まかな形態
- 図:人間の脳の構造
- 図 2.1a 左半球の側面図
- 図 2.1b 右半球の下内側図
- 小脳の構造
- 小脳の主な機能
- 2.3 三重脳(トライユン脳)
- 図 2.2 マクリーンの三重脳概念に基づく脳の階層的な組織の模式図
- 爬虫類脳
- 自律神経系
- 髄鞘を持つ迷走神経の機能
- 環境の影響
- 古哺乳類脳
- 古哺乳類脳とストレス調整
- 新哺乳類脳の進化
- 前頭前野(PFC)の役割
- 新哺乳類脳の進化の重要性
- 2.4 脳の成長曲線と人間の脳の拡大の制約
- 霊長類の進化と脳のサイズ
- 脳の特定部分のサイズ比較
- 霊長類の脳のサイズ
- 人間の脳の成長の印象
- 人間の脳と他の霊長類の比較
- 脳と体の成長の関係
- 脳の進化的変化
- 基底核の進化
- 帯状回と小脳
- 前頭葉の特異性
- 前頭葉と頭頂葉の機能
- 頭頂葉の進化
- 脳の寿命と相関関係
- 人間の新皮質の細胞構造
- 脳の成長と折りたたみ
- 脳の地図作成と受容体の分布
- 人間の脳の進化的発生学
- 🧬 異時性(Heterochrony)の階層的分類体系(Fig. 2.4 の解説)
- 📌 まとめ図(ツリー形式)
- 🧒 Paedomorphosis(幼形成熟)
- 👴 Peramorphosis(過形成熟)
- 🔍 図解イメージ(進化的変化の軸)
- 🧠 なぜ重要なのか?
- 🧒 Paedomorphosis(幼形成熟)
- 👴 Peramorphosis(過形成熟)
- 🧠 追加の視点:家畜化と異時性
- 🧬 異時性(Heterochrony)の分類と動物例
第2章
人間の脳:解剖学、進化、機能
概要
人間の脳は、進化の中で最も複雑な器官です。他の霊長類の脳と比べて、より多くのニューロン(神経細胞)とシナプス(神経細胞間の接続)を持っています。体重に対して非常に大きく、明確にひだが多いのが特徴です。進化の過程で、いくつかの脳の部分が不均衡に大きくなりました。これらの脳の領域は主に、感情の処理、自分自身や他者の心を理解し反映すること、記憶、社会的意思決定、行動計画に関与しており、人間の脳が社会的な問題に対処するために適応していることを示唆しています。また、人間の脳は成熟が遅いことでも目立ちます。これは、成人になるまでに学ぶ必要のある膨大な社会的情報に関連しています。ニューロン間の相互作用は、神経調節物質や神経伝達物質の働きによって維持されており、これらの多くは古代から存在し、植物や動物で複数の目的に使用されてきました。これらは、防御行動や攻撃的行動、社会的な結びつき、抑制的な制御を調整するのに役立ちます。
キーワード
社会的認知、遅い成熟、社会的結びつき、防御、攻撃的行動、抑制的制御、神経伝達物質
2.1 人間の脳への序論
人間の脳は、おそらく進化の中で最も複雑な器官です。脳はすべての身体機能の「制御システム」であり、人間が環境をどのようにナビゲートするかを調整します。環境の複雑さは、高度な認知能力や感情能力の進化を促し、これまでにない驚くべき行動の柔軟性を可能にしました。人間の脳が持つ多様な機能は、長い複雑な進化の歴史の結果です。
- 初期の調節システム:
- 最初の原始的な調節システムは、受容体や生化学的メカニズムで構成されており、細菌のような単純な生物が環境の変化に対処するために機能しました。
- 例えば、毒素を避けたり栄養素に近づいたりするためのもので、これは数十億年にわたって行動を調整する主な方法でした(Allman 1999)。
- 神経系の進化:
- しかし、これらのシステムはまだ神経系ではありませんでした。神経系の基礎となるニューロンは、受容体、導体、効果器を持っています。
- ニューロンは電気的に刺激され、ニューロン間のコミュニケーションはシナプスを通じた機能的な接続に依存しています。シナプスは約10億年前により原始的なプロトシナプスから進化したと考えられています(Ryan and Grant 2009)。
- 最初の神経系:
- 最初の本格的な神経系は、5億年以上前に腔腸動物(例:クラゲ)に現れ、小さなニューロンの集まりから成り立っていました。
- その後、これらの初期の神経系が拡張し、複雑で階層的に組織された器官、すなわち「脳」へと進化しました(Allman 1999)。
原始的な神経系の特徴
- 原始的な神経系は約5億年前に出現しました。
- それらは受容体、導体、効果器を持つニューロンの集まりであり、電気的興奮性を特徴としています。
脳の進化と機能
まず第一に、脳はおそらく、生物の生存を脅かす危険を避けるための装置として進化しました。また、食料、住居、配偶者などの資源を評価するためにも役立ちました。しかし、進化のある時点で、脳は環境の変化に「反応」する準備をする戦略から、環境を積極的に探索する戦略に変わりました。つまり、脳の進化は、生物が環境(特に社会的環境)を利用し操作するための行動能力を最大化する歴史です。これは、生物の繁殖適応度のために重要です。
しかし、脳の進化は常に脳のエネルギー消費によって制約されています。ニューロン(神経細胞)はエネルギー的に非常に高価だからです。ニューロンは、自身と環境との間でイオンバランスを維持しなければならず、神経細胞間のコミュニケーションのために多くのエネルギーを必要とします(Fehm et al. 2006)。例えば、樹状突起にはエネルギー供給を維持するために多くのミトコンドリアが含まれています。
ニューロンは神経伝達物質と呼ばれる化学物質や、活動電位と呼ばれる電気的な信号を介してコミュニケーションを行います。活動電位は長距離を移動でき、ミエリン鞘の進化はニューロン間の情報伝達を大幅に加速しました(Hofman 2001)。これらの高価なメカニズムは、進化的な利点があったからこそ選択されたはずです。そうでなければ、脳は存在しなかったでしょう。進化のプロセスは「倹約的」だからです(Northcutt 2001)。
原始的な脳の役割
- 原始的な脳は、生物が危険を避け、資源を探し、交尾するのを助けました。
- しかし、進化のある時点で、生物は環境を積極的に探索する戦略に変わりました。
- 脳のサイズは、ニューロンのエネルギー消費の大きさによって制約されています。
- したがって、大きな脳は非常に大きな利点をもたらさなければ進化しなかったでしょう。
人間の脳の特性
- 人間の脳には、約100,000 kmの接続ファイバーがあり、推定ストレージ容量は1.25テラバイトです。
- 大脳皮質のニューロンの平均数は約160億から200億で、これは脳全体のニューロンの約20%を占めます。
- 人間の脳の総ニューロン数は約860億と報告されています(Azevedo et al. 2009)。
- 人間の皮質の1 mm³には約4,400万のニューロンが含まれ、150 mの樹状突起、さらに100 mの軸索接続、5000万のシナプスがあります。
- グリア細胞の数は、大脳皮質のニューロンの数の約2倍から10倍とされています(Hofman 2001)。他の報告では1:1の比率に近いものもあり、これは他の哺乳類の脳と一致します(Azevedo et al. 2009)。
人間の脳はその複雑さにおいて比類がありません。数十億のニューロンを含み、推定1.25テラバイトの情報を保存できます。人間の脳は他の器官とエネルギー供給を競い合っています。さらに、大きな脳はゆっくりと発達し、適切に成熟するためには大量の環境からの情報が必要です。
脳の進化と機能
まず第一に、脳はおそらく、生物の生存を脅かす危険を避けるための装置として進化しました。また、食料、住居、配偶者などの資源を評価するためにも役立ちました。しかし、進化のある時点で、脳は環境の変化に「反応」する準備をする戦略から、環境を積極的に探索する戦略に変わりました。つまり、脳の進化は、生物が環境(特に社会的環境)を利用し操作するための行動能力を最大化する歴史です。これは、生物の繁殖適応度のために重要です。
しかし、脳の進化は常に脳のエネルギー消費によって制約されています。ニューロン(神経細胞)はエネルギー的に非常に高価だからです。ニューロンは、自身と環境との間でイオンバランスを維持しなければならず、神経細胞間のコミュニケーションのために多くのエネルギーを必要とします(Fehm et al. 2006)。例えば、樹状突起にはエネルギー供給を維持するために多くのミトコンドリアが含まれています。
ニューロンは神経伝達物質と呼ばれる化学物質や、活動電位と呼ばれる電気的な信号を介してコミュニケーションを行います。活動電位は長距離を移動でき、ミエリン鞘の進化はニューロン間の情報伝達を大幅に加速しました(Hofman 2001)。これらの高価なメカニズムは、進化的な利点があったからこそ選択されたはずです。そうでなければ、脳は存在しなかったでしょう。進化のプロセスは「倹約的」だからです(Northcutt 2001)。
原始的な脳の役割
- 危険を避ける: 原始的な脳は、捕食者から逃げるための反応を促進しました。
- 資源を探す: 食料や住居を見つけるために、環境を評価する能力を持っていました。
- 交尾: 繁殖のためのパートナーを見つけるための行動を助けました。
しかし、進化のある時点で、生物は環境を積極的に探索する戦略に変わりました。脳のサイズは、ニューロンのエネルギー消費の大きさによって制約されています。したがって、大きな脳は非常に大きな利点をもたらさなければ進化しなかったでしょう。
人間の脳の特性
- 接続ファイバー: 人間の脳には、約100,000 kmの接続ファイバーがあります。
- ストレージ容量: 推定ストレージ容量は1.25テラバイトです。
- ニューロンの数: 大脳皮質のニューロンの平均数は約160億から200億で、これは脳全体のニューロンの約20%を占めます。
- 総ニューロン数: 人間の脳の総ニューロン数は約860億と報告されています(Azevedo et al. 2009)。
- 皮質の構造: 人間の皮質の1 mm³には約4,400万のニューロンが含まれ、150 mの樹状突起、さらに100 mの軸索接続、5000万のシナプスがあります。
- グリア細胞: グリア細胞の数は、大脳皮質のニューロンの数の約2倍から10倍とされています(Hofman 2001)。
脳のエネルギー消費
- エネルギー供給の競争: 人間の脳は他の器官とエネルギー供給を競い合っています。
- 発達の遅さ: 大きな脳はゆっくりと発達し、適切に成熟するためには大量の環境からの情報が必要です。
図:人間の脳の構造と機能
以下の図は、人間の脳の主要な部分とその機能を示しています。
+---------------------+
| 大脳皮質 |
| (思考、感情、記憶) |
+---------------------+
| 小脳 |
| (運動調整、バランス)|
+---------------------+
| 脳幹 |
| (基本的な生命機能) |
+---------------------+- 大脳皮質: 高度な思考、感情、記憶を処理します。
- 小脳: 運動の調整やバランスを保つ役割を果たします。
- 脳幹: 呼吸や心拍などの基本的な生命機能を制御します。
このように、人間の脳はその複雑さにおいて比類がなく、数十億のニューロンを含み、推定1.25テラバイトの情報を保存できます。人間の脳は他の器官とエネルギー供給を競い合っており、大きな脳はゆっくりと発達し、適切に成熟するためには大量の環境からの情報が必要です。
大きな脳の生物学的問題
大きな脳は、他の器官とエネルギー供給を競わなければならないため、いくつかの生物学的問題を引き起こします(Fehm et al. 2006)。大きな脳はゆっくりと発達します。なぜなら、神経回路の形成には長い時間がかかり、多くの環境からの情報が必要だからです。このため、脳を持つ生物の繁殖能力が制約されます。未熟な生物は、捕食や飢餓などの環境の危険から保護されなければなりません。この保護は、子孫が繁殖することが親の遺伝的利益になる大人によって行われます。言い換えれば、子孫の未熟さが増すにつれて、親の育児努力も比例して増加します(Trivers 1974; Allman 1999)。これらの生物学的問題は、人間において特に顕著です。では、なぜ人間の脳は存在するのでしょうか?この質問に対する答えは、2.2から2.10のセクションで試みます。
2.2 人間の脳の大まかな形態
人間の脳は、他の哺乳類の脳と同様に、いくつかの部分に分けることができ、これらの部分は互いに強く接続されています。脳の最も基本的な部分は「後脳」(rhombencephalon)と呼ばれ、呼吸、血圧、睡眠-覚醒リズム、そしていくつかの原始的な行動反応など、最も基本的な生理機能を維持します。脳の中間部分は「中脳」(mesencephalon)と呼ばれ、基底核を含み、神経伝達物質を生成する細胞が豊富に存在する黒質(substantia nigra)などの核があります。前脳(prosencephalon)は、内分泌機能や条件付け学習を担当する視床(thalamus)、視床下部(hypothalamus)、および下視床(subthalamus)を含む「間脳」(diencephalon)と、感情調整や認知機能に関与する「大脳半球」(telencephalon)に分けられます。
人間の脳は典型的な哺乳類の脳です。進化的に古い部分(脳幹、中脳)と、最近進化した部分(大脳皮質)を含んでいます。皮質の形態は、細胞構造(サイオアーキテクチャ)によって区別できます。アロコルテックス(allocortex)は、海馬(hippocampus)、内嗅皮質(entorhinal cortex)、帯状皮質(cingulate cortex)、および扁桃体(amygdala)の一部を含みます。新皮質(neocortex)は、約96%の皮質表面を覆う6層構造です。前頭葉と側頭葉はそれぞれ30%、頭頂葉は23%、後頭葉は約15%を占めています。
脳の外側部分は「パリウム」(pallium)とも呼ばれ、より基底的な部分の大部分を覆っています。「パリウム」という言葉は「マント」や「コート」を意味します。この脳の外側部分は、アロコルテックスとアイソコルテックス(isocortex)に分けることができます。アロコルテックスとアイソコルテックスは、細胞構造に基づいて区別されます。アイソコルテックスまたは新皮質は、大きな錐体細胞を含む6層構造で、運動皮質を除いて、明確な層IVが欠けています。また、一次視覚皮質では層IVが3つのサブレイヤーに分けられます。一次感覚皮質は視床(thalamus)や外側膝状体(geniculate bodies)からの投射を受け取り、二次および三次感覚領域に投射し、これらはさらに運動皮質に投射します。進化的に古いアロコルテックスは、嗅覚系(約1%の皮質表面を占める古皮質に相当)、海馬、内嗅皮質、帯状皮質、および扁桃体の一部を含みます(約3.5%の皮質表面を占める原皮質に相当)。アロコルテックスから新皮質への細胞構造の段階的な移行があり、2つの移行部分は「周辺アロコルテックス」(periallocortex)と「前アイソコルテックス」(proisocortex)と呼ばれ、合わせて「メソコルテックス」(mesocortex)と呼ばれます。アイソコルテックスは、皮質表面の約96%を占めています。前頭葉が約32%、側頭葉が30%、頭頂葉が23%、後頭葉が15%を占めています(Zilles 1987)。図2.1aと2.1bは、左半球の側面図と右半球の内側図をそれぞれ示しています。
図:人間の脳の構造
以下の図は、人間の脳の主要な部分を示しています。
+---------------------+
| 大脳皮質 |
| (感情、認知機能) |
+---------------------+
| 中脳 |
| (運動制御、感覚) |
+---------------------+
| 脳幹 |
| (基本的な生命機能) |
+---------------------+- 大脳皮質: 高度な感情や認知機能を処理します。
- 中脳: 運動の制御や感覚情報の処理を行います。
- 脳幹: 呼吸や心拍などの基本的な生命機能を制御します。
このように、人間の脳はその構造と機能において非常に複雑であり、進化の過程でさまざまな役割を持つ部分が発展してきました。
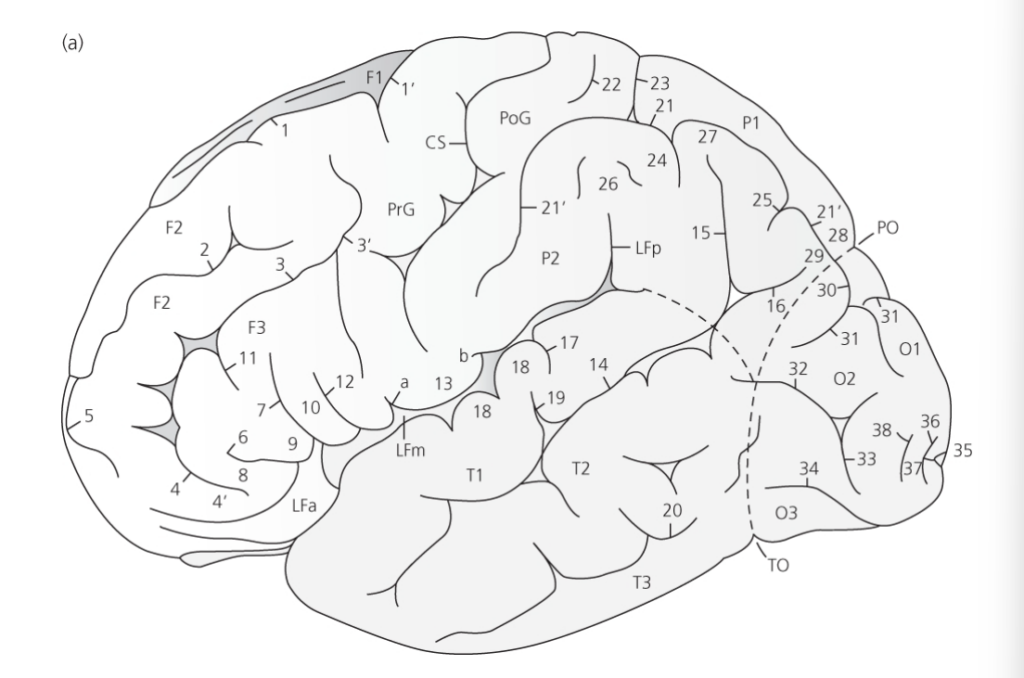
図 2.1a 左半球の側面図
(脳幹と中脳は除去されています)。点線は異なる葉の間の(理論的な)境界を示しています。
(出典: Miguel Marín-Padilla, ‘Mammalian Cerebral Cortex: Embryonic Development and Cytoarchitecture’, in The Human Brain, pp. 6-7, Figure 1, Springer Science + Business Media, 2011)
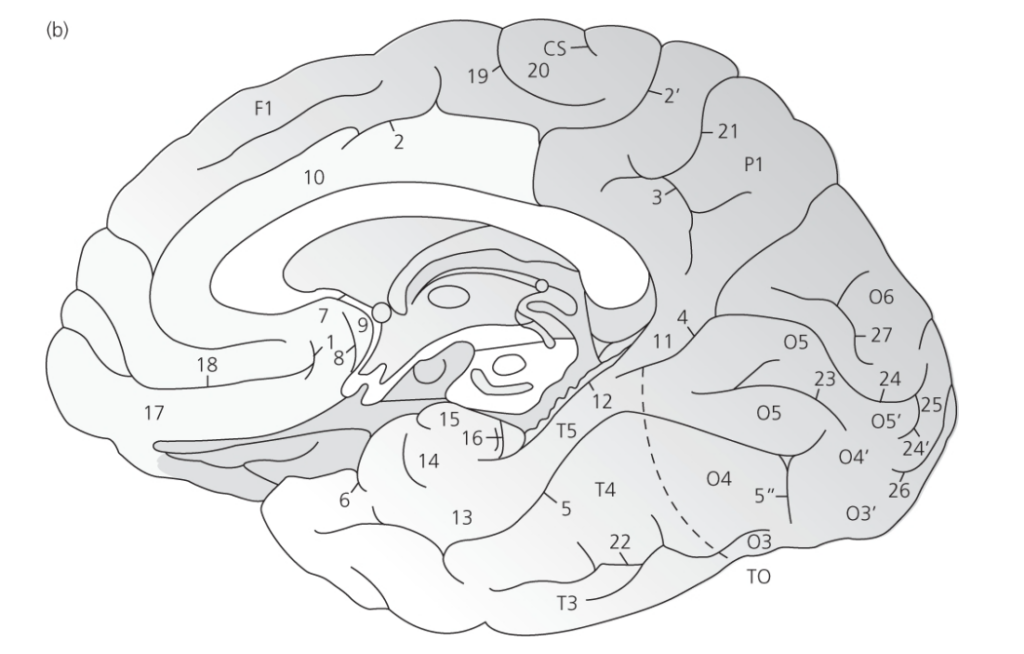
図 2.1b 右半球の下内側図
(脳幹と中脳は除去されています)。
(出典: Miguel Marín-Padilla, ‘Mammalian Cerebral Cortex: Embryonic Development and Cytoarchitecture’, in The Human Brain, pp. 28-9, Figure 13, Springer Science + Business Media, 2011)
小脳の構造
小脳は2つの半球と、中央に位置する「虫部」(vermis)という構造から成り立っています。小脳の半球は4つの葉に分かれています。大脳皮質と同様に、小脳も進化的な起源に基づいて「古小脳」(archicerebellum)、「古哺乳類小脳」(palaeocerebellum)、および「新小脳」(neocerebellum)に分けられます。小脳は多くの細胞を含み、ほぼすべての大脳皮質の部分と接続されています。
小脳は、目の動き、前庭の制御、運動の調整に関して最もよく知られていますが、注意、行動計画、視空間認知、記憶、感情調整など、さまざまな認知機能にも寄与しています(Katz and Steinmetz 2002)。
小脳の主な機能
- 目の動きの制御: 小脳は目の動きを調整します。
- 前庭の制御: 体のバランスを保つ役割を果たします。
- 運動の調整: スムーズな運動を実現します。
- 認知機能への寄与: 注意、行動計画、視空間認知、記憶、感情調整に関与します。
すべてのコミュニケーションは、下位(後脳)と上位(皮質)の大脳および小脳の脳中心間で中脳の構造を通過しなければなりません。これをスキップすることはできません。これは、哺乳類の脳が異なる層で進化的に起源を持ち、階層的に組織されていることの直接的な結果です。
2.3 三重脳(トライユン脳)
人間の脳(およびすべての哺乳類の脳)は、異なる動物のクラスにおける進化的な出現に基づいて、3つの層に大まかに分けることができます(MacLean 1990)。これらの3つの層は、「爬虫類脳」(reptilian brain)またはR複合体(R-complex)、「古哺乳類脳」(palaeomammalian brain)、および「新哺乳類脳」(neomammalian brain)と呼ばれています。この区別は、これらの3つの層がどのように協調して機能するかを単純化したものですが、「三重脳」は脳内の機能の表現を包括的に示すモデルです(図2.2参照)。
三重脳のモデル
- 爬虫類脳: 基本的な生命機能の調整を担当します。爬虫類脳の前方部分は、交尾、同性間の攻撃、服従に関する本能的な行動の座です。
- 古哺乳類脳: 感情の調整、母性のケア、分離の苦痛、環境の遊び探求に関与します。
- 新哺乳類脳: 高度な認知機能を含み、人間の場合は自己反省的な意識を持ちます。
このように、三重脳のモデルは、哺乳類の脳がどのように進化してきたかを理解するための理論的な枠組みを提供します。
f2.2
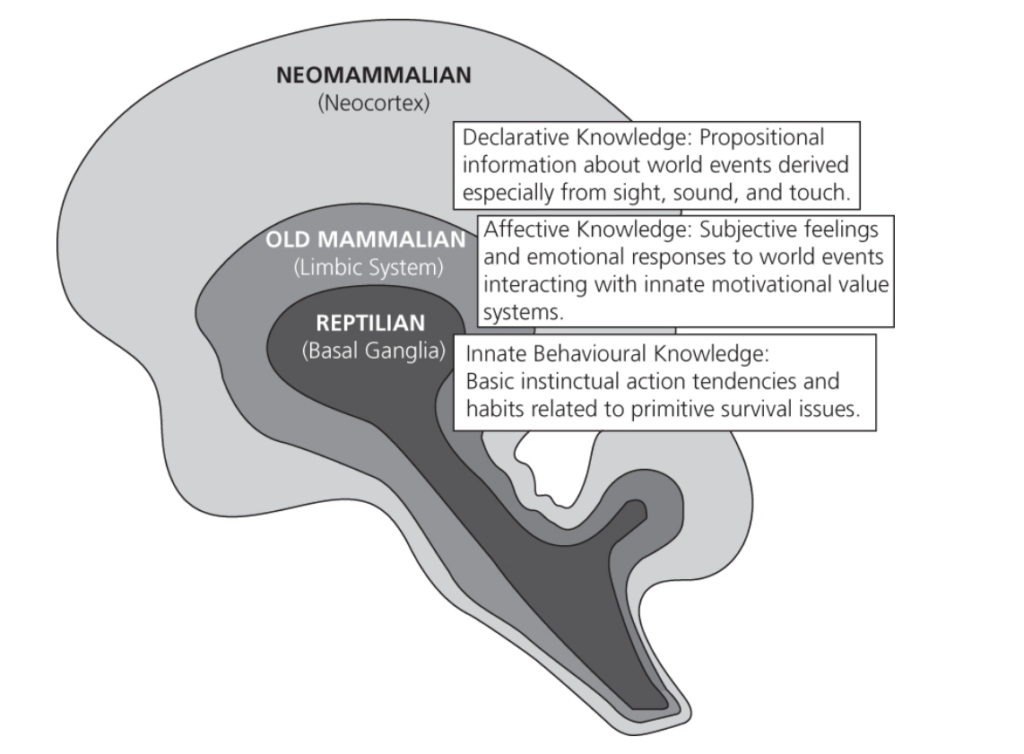
図 2.2 マクリーンの三重脳概念に基づく脳の階層的な組織の模式図
(出典: P.D. MacLean, The Triune Brain in Evolution: Role in Paleocerebral Functions, Plenum Press, New York, Copyright @ 1990, Springer-Verlag US)
爬虫類脳
- 進化的に最も古い構造: 爬虫類脳は脳幹、基底核、線条体から成り立っています。
- 基本的な機能: 体温、血圧、呼吸、睡眠-覚醒、姿勢反射などの調整を行います。
- 本能的行動: 爬虫類脳の前方部分は、交尾、同性間の攻撃、服従に関する儀式的な社会行動や、食事、狩猟などの摂食行動を制御します。
- 固定行動パターン: 本能的行動は、狭く定義された引き金によって引き起こされる固定行動パターンとして表現されます。
- 条件付け学習: 条件付け学習や摂食行動は、線条体の構造によって維持されます。
- 進化の変化: 爬虫類脳は爬虫類と同様の構造を持ち、哺乳類の進化の中で比較的変化が少ないです(MacLean 1990)。
自律神経系
- 進化の過程: 自律神経系(迷走神経系と交感神経系を含む)は、捕食者の脅威や社会的地位の競争、性的パートナーへのアクセスなど、環境の挑戦に対してより柔軟な反応を提供するために進化しました。
- ポリバガル理論: Porgesの「ポリバガル理論」(1995)によれば、最も原始的な脅威反応は、逃げられないときに見られる「固定」または「凍結」行動です。この反応は、無髄の迷走神経の機能に依存しています。
- 戦闘または逃走反応: より進化的に新しい反応パターンは、戦闘または逃走の反応を動員することです。このシステムは交感神経系の働きによって支えられています。
- 社会的コミュニケーション: 哺乳類での最も最近の進化的発展は、髄鞘を持つ迷走神経の活動に機能的に関連しています。Porgesは、髄鞘を持つ迷走神経が社会的コミュニケーションに関与していると提案しています。
髄鞘を持つ迷走神経の機能
- 心拍数の調整: 髄鞘を持つ迷走神経は、交感神経系を抑制することによって心拍数にブレーキをかけます。
- 心拍変動: 髄鞘を持つ迷走神経の活動の神経生理学的相関は、心拍数の変動性です。呼吸性洞性不整脈として測定できます。
- 機能不全の影響: 髄鞘を持つ迷走神経の機能不全は、より原始的なシステム(すなわち、動員または固定)を活性化し、心拍数の変動性を低下させます。
環境の影響
- 胎内の逆境: 胎内での逆境(例:妊娠の第2トリメスター中)は、ポリバガルシステムの成熟を妨げる可能性があります。これは、精神的健康だけでなく、ストレスの調整能力(免疫応答を含む)にも深刻な影響を与える可能性があります(Porges 1992, 2009; Grippo et al. 2007)。
古哺乳類脳
- 大脳辺縁系: 古哺乳類脳は大脳辺縁系の構造とほぼ同じです。このシステムの進化は、広範な親のケアや他の社会的感情の発展と密接に関連しています(Panksepp 1998)。
- 感情の調整: 古哺乳類脳は、恐怖や怒りなどの感情を調整し、母性行動、分離の苦痛、環境の遊び探求に関与します。
- 親のケアの選択: より大きな親のケアと育成の選択は、共感や近親者の個別認識などの社会的感情の発展における重要な相関関係と見なされています(Bernhardt and Singer 2012; Gonzalez-Liencres et al. 2013; Preston 2013)。
- ストレス調整: 古哺乳類脳は原始的な哺乳類に見られますが、霊長類の進化の過程でいくつかの重要な変化を遂げています。例えば、ストレス調整や自分の身体状態の内受容的な認識に関与しています(Craig 2003; セクション2.9参照)。
古哺乳類脳とストレス調整
- 古哺乳類脳(大脳辺縁系): ストレスの調整や内受容的な認識に関与しています。大脳辺縁系の構造は、特に幼少期の経験の影響を受けやすく、個人のストレス反応を大きく左右する可能性があります。
新哺乳類脳の進化
- 新哺乳類脳: 最も最近進化した部分で、さまざまな感覚情報を統合し、柔軟な行動を可能にします。
- 機能:
- 下位脳からの行動衝動を抑える。
- 固定的な本能行動から解放し、行動の選択肢を選ぶ。
- 機能的な領域を超えて情報を統合する。
- 将来の行動を計画し、未来の出来事を予測する(先見の明)。
- 情報を記憶する。
- 意識の重要性: 新哺乳類脳の機能を意識することは例外であり、自己反省や他者の認識を反映することは、新哺乳類脳の皮質の拡張なしには不可能です。
- 階層的な組織: 下位脳の機能(例:呼吸)は上位機能とは独立して動作することができますが、その逆はありません。新哺乳類脳の機能は、中脳や視床下部との正常な接続に依存しています(MacLean 1990)。
前頭前野(PFC)の役割
- 前頭前野(PFC): 霊長類の新哺乳類脳の中で最も最近拡大した構造です。
- 接続: 脳幹、間脳、そして大脳辺縁系と相互接続されています。
- 情報の受け取り: 内部環境、覚醒、欲求、動機、感情の内臓的相関に関する情報を提供します。
- PFCのサブディビジョン:
- 内側、外側、眼窩前頭皮質: すべてが扁桃体や海馬からの情報を受け取り、相互に接続されています。
- 内側および眼窩前頭皮質: 感情行動や基本的な欲求の制御に関連する機能を主に担います。
- 外側前頭前野: 情報の時間的統合や将来の行動目標の組織に関与します。
- 社会的情報処理: 側頭葉と前頭前野の間には強い半球内接続があり、これらの接続は社会的情報の処理に特に関連しています(Hopkins and Rilling 2000)。
- 早期経験の影響: 脳の接続性は早期の経験に大きく依存しています。例えば、極度の不安は扁桃体と前頭前野の接続性を変化させ、脳が脅威を評価する方法に機能的な影響を与える可能性があります(Birn et al. 2014)。
新哺乳類脳の進化の重要性
- 人間の進化における変化: 新哺乳類脳は人間の進化の中で最も重要な変化を遂げました。人間と最も近い親戚との脳のサイズを比較することで、どの部分でこれらの変化が起こったかを示すことができます(Semendeferi et al. 1997; Semendeferi and Damasio 2000)。
2.4 脳の成長曲線と人間の脳の拡大の制約
- 脳のサイズの変動: 種によって脳のサイズは大きく異なり、哺乳類の中でも小さな哺乳類の脳は数グラムしかありませんが、クジラの脳は最大で10kgにもなります。
- 脳の重さと体重の関係: 一般的に、哺乳類の脳の重さは体重に比例します。脳の重さは体重の75%に相当するように増加します(Allman 1999)。
- アロメトリー: 脳の重さや体積を体重に対して正規化する手法をアロメトリーと呼びます。進行指数(PI)は、この方法を用いて得られ、特定の種の脳の重さを祖先の種や最後の共通祖先に似た現存種の脳の重さで割ったものです。PIは進化的傾向を反映し、特定の脳領域の機能的重要性を示します(Rapoport 1990)。
- アロメトリ計算: アロメトリ計算は、現存種の祖先種からの推定に基づいて、器官のサイズや体積の増加を体のサイズや重さに対して決定することができます。
霊長類の進化と脳のサイズ
- 霊長類の起源: 最初の霊長類は昆虫食性の動物から進化しました。そのため、昆虫食性から初期の霊長類への進化の過程で、祖先に似た現存種(例:チュパイア)や、原始霊長類(キツネザル、ブッシュベイビー、ガラゴ)、猿(サル)、および類人猿(人間を含む)を比較することが理にかなっています。
- 脳のサイズ: 平均して、霊長類は非霊長類の哺乳類と比べて、体重に対して2〜3倍大きな脳を持っています。これは胎児の脳の体積にも当てはまります(歯のあるクジラを除く)。霊長類の中では、果物を食べる動物は葉を食べる動物よりも平均して大きな脳を持っています。これは、果物を探すことが季節的かつ空間的に分散しているため、計算リソースに対する要求が大きいからかもしれません(Allman 1999)。
- 脳と消化器官の競争: 初期の研究では、脳のサイズと腸のサイズには逆相関があることが示唆されています。つまり、脳は消化器官とエネルギーを競い合っていると言えます。なぜなら、総エネルギー使用量は体重に依存するからです。
- 人間の脳のエネルギー消費: 人間の脳は約15Wの比較的低いエネルギー消費をしていますが、体全体のエネルギーの15〜20%を消費しています。これは、成人の脳の重さが約1,400gで、体重のわずか2%に過ぎないため、非常に高い割合です(Aiello and Wheeler 1995)。
- 脳サイズの増加要因: 最近の考察では、脳のサイズが人間の進化の過程で増加した要因として、エネルギーの補助(協力的な育児)、食事の質の向上、移動にかかるエネルギーの削減(例:二足歩行)が挙げられています(Navarrete et al. 2011)。
脳の特定部分のサイズ比較
- 脳の特定部分のサイズ: 近縁種と遠縁種の間で、体重を考慮しながら脳の特定部分のサイズを比較することで、どの脳領域が進化の過程で大きくなり、どの部分が相対的に小さくなったかを知ることができます。
- 嗅覚系の重要性の低下: 進行指数(PI)に基づくと、霊長類の系統において嗅覚系の重要性は進化の過程で低下したと結論できます。例えば、昆虫食性の動物では嗅球が脳全体の18%を占めていますが、チュパイアでは約7%、ガラゴでは4%、サルでは約0.2%、人間では0.01%です(Rapoport 1990)。
- 嗅球のサイズ: 人間の嗅球のサイズは、比較可能な体重の昆虫食性動物に対して約1/40から1/50に過ぎません。一方で、人間の新皮質のPIは156であり、最も近い現存の親戚であるチンパンジーの新皮質のPIは58です(Rapoport 1990)。
- 新皮質の拡大: したがって、人間の系統とチンパンジーの系統が共通の祖先から分岐した約600万年前以降、人間の新皮質の拡大が加速したようです。人間の新皮質は、同じ体サイズの類人猿の約3倍のサイズであり、チンパンジーの脳と比べても約3倍大きいです(Jones et al. 1992)。
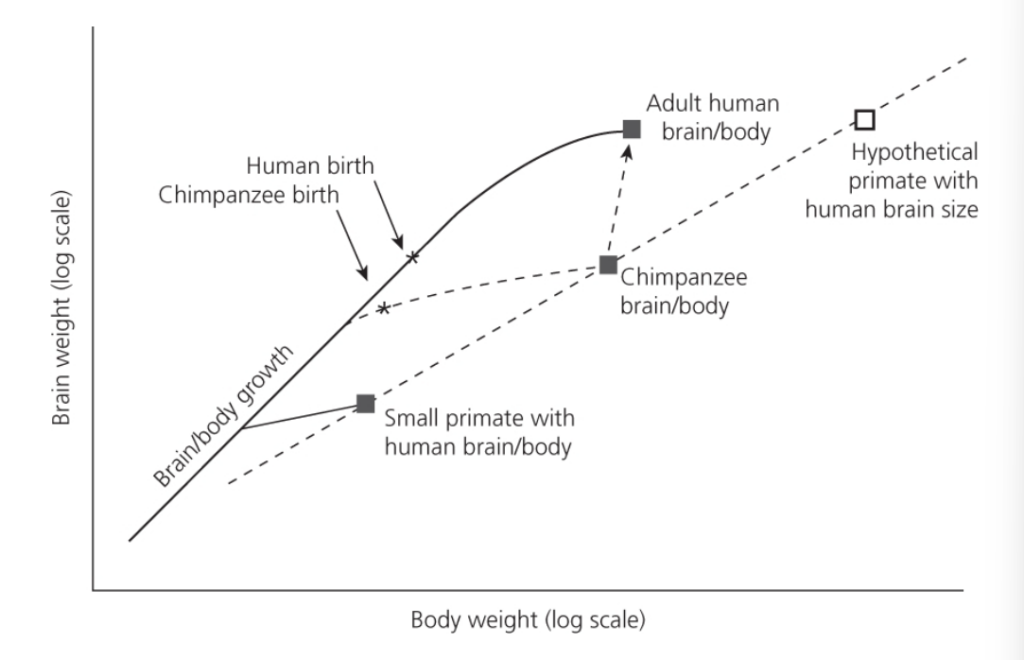
図2.3 人間と他の霊長類種における脳と身体の成長の関係。
注目すべきは、人間における胎児期(脳と身体が同じ速度で成長する期間)の長さが、早産によって出生後の時期まで大きく延長されている点である。人間の脳は、人間の体重に相当する霊長類と比べて、かなり大きい。
(出典:Stephen Jones、Robert D. Martin、および David R. Pilbeam『The Cambridge Encyclopedia of Human Evolution』、p.118、図1、ケンブリッジ大学出版、カナダ、著作権 ©1994、ケンブリッジ大学出版)
霊長類の脳のサイズ
- 霊長類の脳の大きさ: 霊長類の脳は、体重に対して平均して予想されるサイズの約3倍の大きさです。
- 人間の脳の特異性:
- 人間の新皮質は、体重に対して予想されるサイズの150倍以上の大きさです。
- 一方で、嗅球(嗅覚に関与する脳の部分)は、原始的な霊長類と比べて約1/50のサイズです。
人間の脳の成長の印象
- 脳の成長の例え: 人間の体がほかの霊長類のように脳の重さに対して成長した場合、体重は約450kg、身長は約3mになると考えられます。
- これは、時々「キングコングモデル」と呼ばれる考え方です(異なる見解についてはHerculano-Houzel 2009を参照)。
- 出生後の脳の成長: 人間の脳は出生後も高い速度で成長し続けます。これは、人間が出産時に未熟であることに関連しています(Jones et al. 1992)。
- この特徴は、人間の骨盤の解剖学的構造による設計上の妥協を反映しています(詳細は第3章を参照)。
- 図の参照: 脳の成長に関するイラストは図2.3を参照してください。
以下に、提供された内容を高校生にも理解できる程度に平易に噛み砕いて逐語的に日本語に翻訳したものを示します。
人間の脳と他の霊長類の比較
- 人間の新皮質のサイズ: 人間の新皮質は、他の類人猿と比べて約3倍の大きさです。
- 体重に対する脳の関係: もし人間が他の霊長類と同じように脳と体の関係があった場合、人間は約3メートルの身長で、体重は450kgになると予想されます。
脳と体の成長の関係
- 図2.3: 人間と他の霊長類の脳と体の成長の関係を示しています。
- 人間の胎児期(脳と体が同じ速度で成長する期間)は、早産のために出生後も長く続きます。
- 人間の脳は、体重に対して予想されるサイズよりもかなり大きいです。
脳の進化的変化
- 古い脳の部分の成長: 人間では、進化的に古い脳の部分(特に海馬、内嗅皮質、扁桃体)が不均衡に大きくなっています。これらの部分の進行指数(PI)はそれぞれ4.2、5.5、3.9です(Rapoport 1990)。
- これらの変化は、感情の調整(恐怖、攻撃、愛着、共感)や自伝的記憶の形成における辺縁系の機能的重要性の増加を反映しています。
- 海馬のCA1領域: 海馬のCA1領域は新皮質と最も広く接続されており、サル類では海馬全体の44%を占めています。人間ではそのPIは6.6です。
- 海馬の一部である歯状回も不均衡に拡大しており、昆虫食性動物に対してPIは2.6です。この領域は成人の神経新生に重要であり、行動の柔軟性や適応性に関連しています。成人の神経新生が損なわれると、うつ病や統合失調症、さまざまな形の認知症に関与していると考えられています(Kempermann 2012)。
- 扁桃体の機能: 扁桃体は複数の機能的ユニットに分かれています。扁桃体の皮質領域は採餌や性的行動を調整し、皮質下の核は防御メカニズムや攻撃行動の学習に関与しています(Allman and Brothers 1994; Amaral 2002)。
- 扁桃体は対人空間の調整にも関連しており、扁桃体のサイズの個人差は生涯にわたる社会的ネットワークのサイズや複雑さを予測します(Bickart et al. 2011)。
基底核の進化
- 基底核のサイズ: 基底核も進化の過程でサイズが増加しており、ヒトの線条体複合体のPIは16.4です(チンパンジーは9.0、原始霊長類は4.7)。これは、動機づけ行動や報酬の予測、期待の重要性が増していることを示唆しています(Knutson et al. 2001)。
帯状回と小脳
- 帯状回の進化: 帯状回は辺縁系に属し、最も前方の部分は進化的に重要な変化を遂げており、重要な刺激と非重要な刺激の区別や意思決定に寄与しています(Allman et al. 2001; Paus 2001)。
- 小脳のサイズ: 小脳も同じ体サイズの霊長類に比べて大きくなっていますが、他の類人猿と比べると小さくなっています(Rilling and Insel 1998; MacLeod et al. 2003)。小脳のサイズの増加は、新皮質の拡大に追いついていないようです。
前頭葉の特異性
- 前頭葉のサイズ: 一見すると、ヒトの前頭葉は新皮質のサイズに対して、同じ体サイズの類人猿と比べて大きくないという驚くべき結果があります(Semendeferi et al. 1997, 2002)。
- しかし、ヒトの前頭前皮質(PFC)は、同じ脳サイズの霊長類に比べてかなり複雑で、ひだが多いです。後頭側頭葉や頭頂葉も同様です。
- 前頭極の機能: 前頭極は未来の行動計画や行動の開始に関与しており、内側前頭前皮質(OFC)は社会的刺激の感情的な重要性を評価する役割を果たしています(Bechara et al. 2000a, 2000b)。
- OFCの内側部分が損傷すると共感に深刻な障害が生じ、外側部分の機能不全は動機や意欲の欠如につながります(Bechara et al. 2000a)。
- 社会的認知プロセス: 内側前頭前皮質は、自分自身や他者の思考や意図を表現する社会的認知プロセスに特に関与しています(Frith and Frith 1999, 2001)。
- 特異な溝: 特に重要なのは、パラシンギュレート溝で、これは50%の個体にしか見られず、進化的に適応的な変化が進行中であると考えられています(Paus et al. 1996; Walter et al. 2004)。
前頭葉と頭頂葉の機能
- 前頭葉の役割:
- 前頭極: 未来の行動計画や行動の開始に関与しています。
- 内側前頭前皮質(OFC): 社会的刺激の感情的な重要性を評価します。
- 内側OFC: 共感などの社会的感情を制御します。
- 外側OFC: 動機や意欲を調整します。
- 内側前頭前皮質: 自分自身や他者の思考や意図を表現する社会的認知プロセスに特に関与しています。
- 側頭葉の進化:
- 側頭葉は他の類人猿と比べて不均衡に大きくなっており、特に白質の体積が増加しています。これは、言語の理解や生成、顔の処理、意図的な動きの認識における機能的重要性を反映している可能性があります(Rilling and Seligman 2002)。
- 側頭極: 自伝的記憶の保存に寄与しています。
- 紡錘状回: 側頭葉の下部にあり、顔の不変的な特徴を認識するために重要です。これにより、顔がどの角度から見えても個人を特定できます。
頭頂葉の進化
- 頭頂葉の進化の不明瞭さ: 頭頂葉の進化の歴史は、ある程度不明瞭ですが、下頭頂小葉(上縁回と隣接する角回を含む)は最近の進化的再編成を経た可能性があります。
- この領域は前頭葉や側頭葉と豊富に接続しており、多感覚の統合や自己認識、自己と他者の区別に重要な役割を果たしています(Torrey 2007)。
- この脳領域が損傷すると、身体の無視(例: 右頭頂葉の損傷後の無知症)、洞察や自己反省の欠如、思考や行動の誤認(他者によって作られたと感じること)が生じることがあります(Torrey 2007)。
脳の寿命と相関関係
- 脳の相対的な重さと寿命: 霊長類の相対的な脳の重さは、異なる種の個体の相対的な寿命とも密接に関連しています。
- 長寿の種は、短命の種よりも統計的に多くの逆境(食糧不足や予測不可能な環境変化)を経験するため、これに対処するためにより多くの計算資源が必要です。
- 人間の脳の重さから予測される最大寿命は約101.5年で、実際の最大寿命に近いです。
- 人間では、相対的な寿命が新皮質、扁桃体、視床下部、小脳のサイズと相関していることがわかっています(Allman 1999)。
人間の新皮質の細胞構造
- 前頭前皮質のニューロン数: 前頭前皮質のニューロンの総数は進化の過程で増加しており、人間は他の類人猿と比べて約4倍のニューロンを持っています。人間は約14 cm³のニューロンを持ち、チンパンジーは約2.8 cm³です。
- 一方で、ニューロンの密度は逆の関係を示しており、人間は接続組織の増加により、立方センチメートルあたりのニューロン密度が最も低いです。つまり、ニューロンの数が増えることで白質の密度が制約されています。
- 人間の灰白質は脳全体の約50%を占めており、ヒトの新皮質は40%の白質を含んでいます。したがって、脳のサイズも重要ですが、機能に関する脳の再編成のパターンも同様に重要です(Hofman 2001; Jerison 2001)。
脳の成長と折りたたみ
- 脳の成長の制約: 皮質の表面がさらに成長するには、頭蓋骨のサイズが大きくなる必要があり、出産時に通過できなくなるか、出産のタイミングがさらに早まる必要があります。
- 皮質の折りたたみ(ひだ形成): 脳の進化において、神経伝達を促進するために接続軸索の数と長さを減少させる方法として、皮質の折りたたみが生じました。
- ニューロンをモジュール回路に区分けすることで接続ファイバーの数が減少し、皮質の折りたたみによって接続組織の長さが短縮されました(Hofman 2001)。
- つまり、ひだ形成は、大きな脳が最小の体積に最大の表面積を詰め込むための解決策を表しています。
- 成人の人間の脳では、約3分の2の皮質表面が溝に埋もれており、表面に露出しているのは3分の1だけです。
- すべてのひだと溝は出生時にすでに見えていますが、皮質の表面の3分の2を溝に隠すには20年の成長が必要です(Allman 1999; Roth and Wullimann 2001)。
- モジュール化された組織: ニューロンレベルでのモジュール化された組織は、進化心理学が提案する機能に基づく組織と部分的に一致している可能性があります(第1章を参照)。
脳の地図作成と受容体の分布
- 脳の地図作成:
- 100年以上にわたり、科学者たちは脳の地図を作成しようとしてきました。これは、皮質の層の数、細胞の密度、ニューロンの形状、または髄鞘の程度に基づいています。
- 最も影響力のある新皮質の地図は、コルビニアン・ブローマンによって提案されました。彼の地図は、脳の機能的な特化と一致していることがわかりました。
- 現在でも、機能的脳イメージングの研究では、ブローマンの地図が参照されていますが、やや単純化されすぎているかもしれません。
- 新しいアプローチ:
- 最近のアプローチでは、従来の区分手法と脳イメージング技術(MRI、DTI、PET、受容体オートラジオグラフィーなど)を組み合わせています。
- これにより、アセチルコリン、ノルエピネフリン、ドーパミン、セロトニン、グルタミン酸、GABAの受容体が人間の新皮質や古皮質に不均一に分布していることが示されました。
- 異なる受容体の分布パターンを2次元座標に重ね合わせることで、脳の特定の領域の受容体の指紋を視覚化できます。このような分子構造の脳マッピングは、従来の地図よりも正確であり、健康な被験者や精神障害のある患者の認知や感情プロセスの機能的表現をより正確に定義するために使用される可能性があります(Toga et al. 2006)。
人間の脳の進化的発生学
- 進化的発生学(ヘテロクロニー):
- 進化的発生学は、発生イベントのタイミングや速度の変化が、祖先の同様のパターンに対してどのように進化的修正が発生するかを説明します(McKinney and McNamara 1991)。
- エルンスト・ヘッケルが提唱した「個体発生は系統発生を繰り返す」という考え方は、今では不正確とされていますが、異なる種の胚の初期段階は後の発生段階よりも似ていることが多いです。
- 個体発生の発展は非常に保守的ですが、個体は祖先の成体段階を経ることはなく、発生遺伝子の変化が早期および後期の発生に影響を与えます(Deacon 1990a, 1990b)。
- したがって、発生過程は加速されて早期に終了することもあれば、遅延して後期に開始されることもあります。
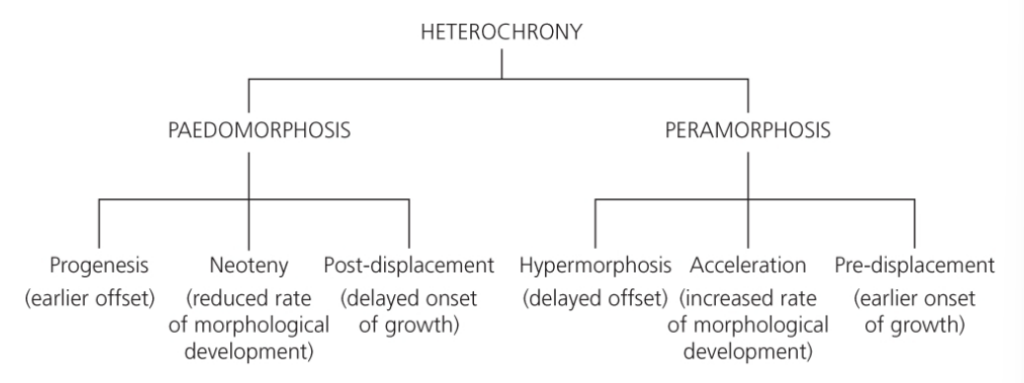
図2.4 異時性プロセスの階層的分類。
(出典:Michael L. McKinney「Classifying and Analyzing Heterochrony」所収、『Heterochrony: The Evolution of Ontogeny』、p.17、図2-3、著作権 ©1991、Springer Science + Business Media)
※「異時性(heterochrony)」とは、生物の発生過程における時間的な変化、すなわちある器官や特徴が他の種や祖先に比べて早くまたは遅く発達する現象を指します。
異時性(heterochrony)の分類体系に関する図(Fig. 2.4)は、生物の発生(ontogeny)の進化的変化を、発生のタイミングや成長の速度などの観点から階層的に分類したものです。以下にその内容を簡潔に整理・解説します。
🧬 異時性(Heterochrony)の階層的分類体系(Fig. 2.4 の解説)
🔸 第一階層:異時性の2大分類
- Paedomorphosis(幼形成熟)
→ 子どもの特徴が大人になっても残る進化的変化。 - Peramorphosis(過形成熟)
→ 通常の大人よりもさらに発達した特徴を持つ進化的変化。
🔸 第二階層:変化のメカニズムによる分類(発生過程の調整方法)
🧒 Paedomorphosis のメカニズム(3つ)
| メカニズム名 | 意味 | 例 |
|---|---|---|
| Progenesis | 発達終了が早まる(早く成体になる) | 短い発達期間で繁殖できる |
| Neoteny | 成長の速度が遅くなる(形態が子供のまま) | サンショウウオの幼体形態のまま繁殖 |
| Postdisplacement | 発生の開始が遅れる | 特定の器官の発達が遅く始まる |
👴 Peramorphosis のメカニズム(3つ)
| メカニズム名 | 意味 | 例 |
|---|---|---|
| Hypermorphosis | 発達終了が遅れる(より長く成長し続ける) | より大きな体や器官が形成される |
| Acceleration | 成長の速度が速まる | 同じ時間でより発達が進む |
| Predisplacement | 発生の開始が早まる | より早い段階で成長が始まる |
📌 まとめ図(ツリー形式)
Heterochrony
├── Paedomorphosis(幼形成熟)
│ ├── Progenesis(早く終わる)
│ ├── Neoteny(ゆっくり進む)
│ └── Postdisplacement(遅く始まる)
└── Peramorphosis(過形成熟)
├── Hypermorphosis(遅く終わる)
├── Acceleration(速く進む)
└── Predisplacement(早く始まる)
この分類は、進化発生生物学(evolutionary developmental biology, Evo-Devo)において、生物の形態変化や多様性の起源を理解するために非常に重要です。
以下に、異時性(heterochrony)の各タイプについて、具体的な動物の例を交えてさらに詳しく説明します。理解しやすいように、まず Paedomorphosis(幼形成熟) と Peramorphosis(過形成熟) に分けて、それぞれのサブタイプごとに見ていきます。
🧒 Paedomorphosis(幼形成熟)
= 子どもの特徴が成体にも残る進化的変化
| タイプ名 | 意味 | 具体例 | 解説 |
|---|---|---|---|
| Progenesis(早期成熟) | 発達の終了が早まる | 一部の線虫や無脊椎動物 | 成体になる前に性成熟し、成長を止めることで子供の形のまま繁殖可能になる。 |
| Neoteny(成長速度の減速) | 成長の速度が遅くなる | ウーパールーパー(メキシコサンショウウオ) | 幼体のまま性成熟(外鰓など幼体的特徴を維持)。ホルモン(甲状腺ホルモン)の不足が原因。 |
| Postdisplacement(発達開始の遅延) | 発達の開始が遅れる | 一部の哺乳類の顔面構造 | ヒトの顔の形が他の霊長類より平坦で幼児的なのは、顔の成長が遅く始まることによると考えられている。 |
👴 Peramorphosis(過形成熟)
= より発達した形質を持つ進化的変化
| タイプ名 | 意味 | 具体例 | 解説 |
|---|---|---|---|
| Hypermorphosis(発達終了の延長) | 発達がより長く続く | マンモスや現生ゾウ | 長期間成長し続けることで、大きな体や長い牙が形成される。 |
| Acceleration(発達速度の加速) | 発達速度が速くなる | **トリ(鳥類)**のくちばし形成 | 恐竜の子孫とされる鳥類では、顔の一部(口部)の成長が加速し、くちばしとして突出する形に。 |
| Predisplacement(発達開始の早期化) | 発達が早く始まる | 一部の魚類の鰭の発達 | 胎生魚での鰭の成長が他よりも早く始まり、大きく発達するケースがある。 |
🔍 図解イメージ(進化的変化の軸)
時間軸 →
─────────────────────────────
| | Progenesis:早く終わる(=子供の形のまま)
|---------| Neoteny:ゆっくり進む(=子供の特徴が残る)
| | Postdisplacement:遅く始まる(=成長しきらない)
| | Predisplacement:早く始まる(=より進んだ形になる)
|---------| Acceleration:速く進む(=急成長して完成度が高くなる)
| | Hypermorphosis:遅く終わる(=もっと発達する)
🧠 なぜ重要なのか?
これらの異時性のパターンは、生物の進化的多様性や形態変化を説明する鍵です。たとえば:
- 人間の顔が他の霊長類よりも幼児的な理由(Neoteny)
- 一部の動物が「子どもの姿のまま」繁殖できる現象(Progenesis)
- 恐竜から鳥類への進化における加速発達(Acceleration)
これらすべてが、発生のタイミングと速度の微妙な変化によって起こっています。
異時性(heterochrony)の各タイプに当てはまる他の動物の具体例を以下にタイプ別にまとめてみました。
🧒 Paedomorphosis(幼形成熟)
= 幼体の特徴を成体が保持する進化的変化
🔹 Progenesis(早期成熟)
- ミジンコ(Daphnia)
幼形成熟の代表例。性成熟が非常に早く、子供のまま繁殖を開始。 - ナメクジウオ(頭索動物)
原始的な形態を持ったまま性成熟する。
🔹 Neoteny(成長速度の減速)
- ウーパールーパー(Ambystoma mexicanum)
幼体のまま性成熟し、変態せずに一生を終える。 - 人間(Homo sapiens)
顔や頭の形、体毛の少なさなど、チンパンジーの幼体に似た特徴を持続。 - ブルドッグ
パグやフレンチブルドッグのような犬種では、幼児的な顔(短頭・大きな目)を維持するよう選択繁殖されている。
🔹 Postdisplacement(発達の開始が遅い)
- ヒトの顎と歯の発達
他の霊長類に比べ、歯の発達が遅く、顔がより平らで幼児的。 - 鳥類の翼の発達(一部)
初期の雛の段階で、翼の発達が後回しになる種もあり。
👴 Peramorphosis(過形成熟)
= 通常よりも発達が進む進化的変化
🔹 Hypermorphosis(発達期間の延長)
- 象(Elephas)とマンモス(Mammuthus)
長い発達期間で巨大な体を持つ。 - ナガスクジラ(Balaenoptera musculus)
巨体になるまで非常に長い時間をかけて成長。 - キリン
首が長くなるのは、発達が通常より長く続いた結果と考えられる。
🔹 Acceleration(成長速度の加速)
- ニワトリのくちばし形成
胚の段階から口先の発達が早く、くちばしが突き出す。 - プレートサウルス(恐竜)
骨格成長が加速し、巨大化が促進されたと考えられる。 - カエル
オタマジャクシ時代から脚の発達が他種より早い種類もいる。
🔹 Predisplacement(発達の開始が早い)
- マグロの筋肉発達
初期から筋肉が発達し、遊泳能力が非常に高い。 - 一部のトカゲ(例:アノール)
頭部の骨格成長が早く始まり、種特有の形を早期に獲得する。
🧠 追加の視点:家畜化と異時性
人間による**家畜化(domestication)**も、実は多くの異時性の例を生み出しています。
例:
- イヌ・ネコ:幼体のような顔つき、行動パターン(Neoteny)
- ブタやヒツジ:野生型よりも成長が早い(Acceleration)
了解です!以下に、異時性(Heterochrony)の分類とそれぞれの動物の具体例を、図(表)形式でわかりやすく整理しました。
🧬 異時性(Heterochrony)の分類と動物例
| 分類 | サブタイプ | 説明 | 動物の具体例 |
|---|---|---|---|
| Paedomorphosis(幼形成熟) | Progenesis(早期成熟) | 性成熟が早まり、子どもの形のまま成体になる | ミジンコ、ナメクジウオ |
| Neoteny(成長速度の減速) | 成長が遅く、幼体の特徴を保持 | ウーパールーパー、人間、ブルドッグ | |
| Postdisplacement(発達開始の遅延) | 発達が遅れて始まり、十分に成長しない | ヒトの顎・顔面構造、一部鳥類の翼 | |
| Peramorphosis(過形成熟) | Hypermorphosis(発達終了の延長) | 成長期間が長くなり、発達が過剰に進む | 象、マンモス、ナガスクジラ、キリン |
| Acceleration(成長速度の加速) | 成長スピードが上がる | ニワトリのくちばし、プレートサウルス、カエル | |
| Predisplacement(発達開始の早期化) | 成長の開始が早くなる | マグロの筋肉、アノールトカゲ |
📌 ビジュアル・ツリー図(簡略版)
Heterochrony
├── Paedomorphosis(幼形成熟)
│ ├── Progenesis:ミジンコ、ナメクジウオ
│ ├── Neoteny:ウーパールーパー、人間、ブルドッグ
│ └── Postdisplacement:ヒトの顔、一部の鳥の翼
└── Peramorphosis(過形成熟)
├── Hypermorphosis:象、クジラ、キリン
├── Acceleration:鳥のくちばし、恐竜、カエル
└── Predisplacement:マグロ、アノールトカゲ
ヘテロクロニーの分類
| ヘテロクロニーのタイプ | 説明 |
|---|---|
| 幼形成熟(ペドモルフォシス) | 幼児の特徴を保持したまま成長すること。 |
| 過形成熟(ペラモルフォシス) | 成長の開始が遅れること。 |
| 前進(プロゲネシス) | 成長の開始が早まること。 |
| 後方移動(ポストディスプレースメント) | 成長の速度が遅くなること。 |
| 過剰成長(ハイパーモルフォシス) | 成長の終了が遅れること。 |
人間の特徴と幼形成熟
- 人間の頭蓋骨: 人間の成人の頭蓋骨は、幼い類人猿の頭蓋骨に形が似ているという考えが、数十年前に人類学者によって提唱されました。
- 幼形成熟の特徴:
- 人間の体のいくつかの特徴(無毛や膣の前方位置など)は、祖先の幼児的特徴が成人に保持されている結果として解釈されています(McKinney and McNamara 1991)。
- 成人期における遊び心や好奇心の持続は、幼い類人猿(および他の動物)に見られる行動に似ているため、行動的幼形成熟の仮説が生まれました(Montagu 1989)。
- 幼形成熟の生物は、幼児の体の特徴を保持したまま性成熟に達します。
ヘテロクロニーの影響
- 成長の速度の違い: ヘテロクロニーは、発生中に体の異なる部分(脳を含む)の成長がどのように異なるかを説明します。一部の組織は他の組織よりも早く(または長く)成長します(McKinney and McNamara 1991)。
- 人間の特徴: 解剖学的および行動的な特徴、歯の発育や髄鞘形成の研究から、人間は幼形成熟的であることが示唆されています。つまり、幼児的特徴が成人期に保持されているということです。
- 過形成熟: 成長の開始が遅れることも、追加の計算能力を生む要因となるかもしれません。
脳の成長と成熟
- 脳の成熟: 前頭前皮質(PFC)は、発生中に最も遅く成熟する脳の領域であり、髄鞘形成やシナプス生成は30代まで続くことがあります。
- 灰白質と白質の変化:
- 灰白質の体積は出生後4〜5歳まで増加し、その後30代を通じて徐々に減少します。
- 一方、白質と脳梁の厚さは出生から30代、場合によっては50代まで増加します。これらの効果は、男の子に比べて女の子でより顕著です。
- シナプスの過剰生成: 妊娠30週目から出生後の最初の2年間にかけて、シナプスの過剰生成が40%発生し、ピーク時には1秒あたり40,000シナプスが生成されます(Rakic et al. 1986)。
- 新皮質のシナプス生成: 新皮質内では、視覚皮質でシナプス生成が6ヶ月齢でピークに達し、前頭前皮質で2歳でピークに達します。このシナプス生成の延長は、他の霊長類と比べてかなり長いです。
結論
- 人間の脳の成長: 人間の脳の成長は他の霊長類と比べてかなり拡大しており、人間の幼児期の延長は、より大きな幼形成熟へのシフトによって部分的に調整されている可能性があります。
- 選択圧の影響: 社会環境からの選択圧が、この発生タイミングのシフトに寄与している可能性があります(Leach et al. 2003)。
- 複雑な成長のメカニズム: 幼形成熟だけが人間の脳の成長のタイミングに関与しているわけではありません。人間の脳は幼い類人猿の脳に似ておらず、新しい特徴の複雑な追加は、過形成熟と呼ばれる別の発生タイミングプロセスによって調整されている可能性があります。
2.7 脳の側性化と接続性
人間の脳は、多くの機能を持っており、それらは神経ネットワークによって実行されます。これらの機能の多くは、左半球または右半球のどちらかに異なって表れます。この二つの半球による役割の分担を「側性化」と呼びます(Saugstad 1998)。
言語と側性化
- 言語は、機能の側性化の代表的な例と考えられています。
- 「線形」機能(例:スピーチの生成、文法、話し言葉の聴覚処理)は、右利きの人では主に左半球に側性化されています。
- 一方で、「全体的」な理解(例:イントネーション、比喩、感情の抑揚)は、右半球に側性化されています(Mitchell and Crow 2005)。
空間認識と自己認識
- 空間の方向感覚は通常、右半球に位置しています。
- 最近の研究では、自己と他者を区別する能力もある程度側性化されていることが明らかになっています。
- 第一人称の視点は、左下頭頂皮質に表現されます。
- 第三人称の視点は、右側の同じ領域に位置しています。
具体例
- 他の人の行動を模倣することは、左下頭頂皮質を活性化します。
- 逆に、自分の行動が模倣されているのを見ると、右側の同じ領域が関与します(Decety and Chaminade 2005)。
このように、人間の脳はさまざまな機能を持ち、それぞれの機能が左右の半球に異なって分担されています。特に言語に関しては、発話や文法などの「線形」な機能は左半球に、感情や比喩などの「全体的」な理解は右半球に関連しています。
