
第2章 進化心理学の新しい科学
学習目標
この章を学習した後、読者は以下のことができるようになります。
・進化の3つの主要な産物を説明する。
・「進化した心理的メカニズム」の概念を定義する。
・進化仮説を検証するための9つの方法を挙げる。
・進化仮説を検証するための7つのデータ源を特定する。
・適応問題を特定するための6つの手順を挙げる。
進化心理学者のカール・グラマーは、半自然的な文脈、つまりシングルスバーで起こる性的シグナルを研究するために研究チームを結成しました(Grammer, 1996)。彼はバーの中に一組の観察者を配置し、特別に設計された評価フォームを使用して、バーで男性に女性がどれくらいの頻度で触れられたかを記録しました。研究チームの別のメンバーは、各女性がバーを出るときに声をかけ、研究に参加することに同意するかどうかを尋ねました。女性の参加者は写真を撮られ、避妊具の使用や現在の月経周期の時点(例えば、最後の月経の開始からの時間)に関する情報を求める簡単なアンケートに回答しました。その後、グラマーは写真画像をデジタル化し、コンピュータプログラムを使用して、各女性が露出した皮膚の割合を計算しました。
ホルモン避妊薬を服用していない女性のグループでは、シングルスバーの男性は、排卵期頃の最も妊娠しやすい時期にある女性に触れる可能性がはるかに高かった。対照的に、排卵していない女性はあまり触れられなかった。したがって、従来の通説とは異なり、男性は女性がいつ排卵しているかの微妙な手がかりを検出できるかもしれない。しかし、別の解釈もある。排卵している女性も、服装を介してより多くの性的シグナルを示していた。彼女たちはよりタイトで、より露出度の高いブラウスと短いスカートを着用し、より多くの肌を見せていた。したがって、男性が女性がいつ排卵しているかを検出するのが得意であるとは限らないかもしれない。むしろ、排卵している女性は積極的に性的シグナルを送っているのかもしれない。この解釈は、排卵している女性が周期の他の段階にある女性よりも性的な出会いを始めることが多いという別の研究によって支持されている(Gangestad, Simpson, Cousins, Garver-Apgar, & Christensen, 2004)。
これらの研究は、進化心理学という科学の2つの特徴を浮き彫りにしています。一つは、人間の生殖生物学の特徴(この場合は女性の排卵)と顕在的な行動との間に、これまで予期されていなかった関連性を発見することです(Gildersleeve, Haselton, & Fales, 2014)。第二に、男性が女性の排卵時期を検出する適応を持っているのか、あるいは女性が自身の排卵に反応する適応を持っているのか(例:Bryant & Haselton, 2009; Gildersleeve et al., 2014)といった、適応機能について考えることは、新しい研究にとって重要な着想を与えます。
2人の尊敬される科学者によれば、「進化心理学は、過去20年間の行動科学における最も重要な新しい発展の一つであることは間違いない」(Boyer & Heckhausen, 2000, p. 917)。この声明は、本書で取り上げられる進化科学者によってなされた多くの魅力的な発見によって強調されるように、今日でも真実である。
本章は、現代進化生物学と現代心理学の科学的統合である進化心理学という科学の論理と方法に焦点を当てる。それは、包括的適応度理論、親の投資と性淘汰の理論、そして適応の有無を評価するためのより厳密な基準の開発といった、進化生物学における理論的進歩を利用する。進化心理学はまた、情報処理モデル、人工知能からの知識、そして普遍的な感情表現(Cowen & Keltner, 2020; Ekman, 1973)、人々が植物や動物を分類する普遍的な方法(Atran, 1990; Berlin, Breedlove, & Raven, 1973)、植物が人間の心をどのように形作るか(Wertz, 2019)、そして人間の交配戦略における普遍性(Lippa, 2009; Walter et al., 2020)といった、心理学における概念的・経験的進歩も取り入れている。本章の目標は、この新しい統合の概念的基礎を紹介することである。後の章は、この基礎の上に構築される。なぜ心理学が進化生物学と統合される必要があるのかを問うことから始めよう。
人間性の起源
複雑な適応メカニズムの起源に関する三つの理論
数週間裸足で歩いていると、足の裏にたこができます。たこを生成するメカニズム、つまり繰り返しの摩擦によって多数の新しい皮膚細胞を製造することは、足の解剖学的および生理学的構造を損傷から保護する機能を果たします。しかし、数週間車で乗り回しても、車のタイヤは厚くなりません。なぜでしょうか?
あなたの足と車のタイヤは、どちらも物理法則の対象となります。摩擦は物理的な物体を摩耗させる傾向があり、構築するものではありません。しかし、あなたの足は、タイヤとは異なり、別の法則、つまり自然選択の法則の対象となります。あなたの足には、自然選択のためにたこを生成するメカニズムがあります。選択による進化は創造的なプロセスです。たこを生成するメカニズムは、その創造的なプロセスの適応的な産物です。それらが現在存在するのは、過去において、摩擦の結果として余分な皮膚の厚さを発達させる傾向がある遺伝子を、たとえわずかでも持っていた人々が、生存を助けるこの余分な要素を持っていたためです。その結果、彼らは有益な素質を持たない人々よりも多くの子孫を残すために生きました。これらの成功した祖先の子孫として、私たちは祖先の成功につながった適応メカニズムを携えています。
過去1世紀の間に、タコを生成するメカニズムのような適応の起源を説明するために、3つの主要な理論が提唱されてきました。1つは創造論、または「インテリジェント・デザイン」であり、これは最高の神が、最大のクジラから海中の最小のプランクトンまで、単純な単細胞のアメーバから複雑な人間の脳まで、すべての植物と動物を創造したという考えです。創造論は、3つの理由で「科学的理論」とは見なされていません。第一に、その主要な前提から特定の経験的予測が導き出されないため、テストできません。存在するものはすべて、最高の存在がそれを創造したからという理由だけで存在します。第二に、創造論は研究者を新しい科学的発見に導いていません。第三に、創造論は、すでに発見されている有機的メカニズムの科学的説明として有用であることが証明されていません。したがって、創造論は宗教と信念の問題であり、科学の問題ではありません。それが偽であることを証明することはできませんが、予測的または説明的な理論として有用であることが証明されていません(Kennair, 2003)。
第二の理論は播種説です。播種説によれば、生命は地球上で始まったわけではありません。この理論の一つのバージョンでは、生命の種は隕石を介して地球に到着しました。播種説の第二のバージョンでは、地球外の知的生命体が他の惑星や銀河から降りてきて、地球に生命の種を植えました。しかし、種の起源に関係なく、自然選択による進化が引き継がれ、種は最終的に人間や今日観察される他の生命体に進化しました。
播種説は原理的には検証可能である。我々は隕石に生命の兆候がないか研究することができ、それは生命が他の場所で始まったという理論に妥当性を与えるだろう。我々は地球外生命体の着陸の兆候を地球上で探し回ることができる。我々は地球上で始まったはずがない生命体の証拠を探すことができる。しかし、播種説は3つの問題に直面する。第一に、現在、そのような「播種」が行われたという確固たる科学的証拠は地球上にはない。第二に、播種説は新しい科学的発見につながっておらず、既存の科学的謎を説明してもいない。しかし、最も重要なことは、播種説は生命体の因果的説明を単に時間的に遡らせるだけであるということだ。もし地球が本当に地球外生命体によって播種されたのであれば、これらの知的生命体の起源につながった因果的プロセスは何だったのだろうか?
私たちは第三の選択肢、つまり自然選択による進化論にたどり着きます。自然選択による進化論は理論と呼ばれていますが、その基本原則は何度も確認され、一度も反証されたことがないため、ほとんどの生物学者からは事実と見なされています(Alcock, 2013)。その作用の構成要素、つまり遺伝的設計の違いによる示差的繁殖は、実験室と野生の両方で機能することが示されています。例えば、ガラパゴス諸島の異なる島にいるフィンチのくちばしの大きさの違いは、各島で一般的な種子の大きさに対応するように進化してきました(Grant, 1991)。種子が大きければ大きなくちばしが必要であり、種子が小さければ小さなくちばしの方が適しています。自然選択の理論には、科学者が科学理論に求める多くの美点があります:(1)既知の事実を説明する、(2)新しい予測につながる、(3)科学的探究の重要な領域への指針を提供する。
創造論、播種説、自然選択説の3つの理論の中で、真の競争はありません。自然選択による進化は、今日私たちが周りで見ることができる驚くべき生命の多様性を説明できる唯一の既知の科学的理論です。そしてそれは、タコを生成するメカニズムから大きな脳まで、人間性を定義する複雑な適応メカニズムの起源と構造を説明する力を持つ唯一の既知の科学的理論です。
進化の三つの産物
進化の過程には3つの産物があります。適応、適応の副産物(または随伴物)、そしてランダムな効果(またはノイズ)です。表2.1に示されています(Buss, Haselton, Shackelford, Bleske, & Wakefield, 1998; Tooby & Cosmides, 1990)。
表2.1 進化のプロセスの3つの産物
適応
自然選択を通じて存在し、遺伝し、確実に発達する特性であり、その進化の期間中に集団に存在した代替設計よりも生存または繁殖の問題をよりよく解決するのに役立ったため。例:へその緒
副産物
適応問題を解決せず、機能的な設計を持たない特性。それらは、機能的な設計を持つ特性とたまたま結合しているために、それらの適応と共に「運ばれる」。例:へそ
ノイズ
偶然の突然変異、環境の突然かつ前例のない変化、または発達中の偶然の効果などの力によって生じるランダムな効果。例:人のへその特定の形
適応とは、その進化の期間中に生存または繁殖の問題を解決するのに役立ったため、自然選択を通じて存在し、遺伝し、確実に発達する特性と定義することができます(Tooby & Cosmides, 1992, pp. 61-62の後)。
この定義をその核心的な要素に分解してみましょう。適応は、その適応のための「遺伝子」を持っていなければなりません。これらの遺伝子は、適応が親から子へと受け継がれるために必要です。したがって、適応は遺伝的基盤を持っています。もちろん、ほとんどの適応は単一の遺伝子に遡ることはできず、むしろ多くの遺伝子の産物です。例えば、人間の目は、数百の遺伝子によって構築されています。過去の環境が今日の私たちが持つ遺伝子を選択しました。人の生涯にわたる環境は、適応の適切な発達に必要であり、現在の環境は、発達した適応を活性化させる責任があります。
適応は、すべての正常な環境において、種のメンバー間で確実に発達しなければなりません。つまり、適応として認められるためには、生物の生涯の適切な時期に、合理的に完全な形で現れ、したがって、特定の種のほとんどまたはすべてのメンバーに特徴的でなければなりません。これには重要な例外があり、例えば、一方の性にのみ存在する、または集団の特定のサブセットにのみ存在するメカニズム(Buss & Hawley, 2011)など、後で取り上げますが、今のところ、ほとんどの適応は種に典型的であることを強調することが重要です。
適応の確実に発達する特徴は、適応が出生時に現れなければならないという意味ではありません。実際、多くの適応は出生後かなり経ってから発達します。歩行は人間の確実に発達する特徴ですが、ほとんどの人間は生後1年経つまで歩き始めません。乳房は女性の確実に発達する特徴ですが、思春期になるまで発達しません。
適応は、選択のプロセスによって形成されます。選択は、各世代において篩として機能し、繁殖に貢献しない多くの特徴をふるい落とし、貢献する特徴を通します(Dawkins, 1996)。このふるい分けプロセスは世代から世代へと繰り返されるため、各新世代は親世代とは少し異なります。各世代でふるい分けプロセスを通過する特徴は、集団に存在する代替の(競合する)設計よりも、生存または繁殖の適応問題の解決により良く貢献するためです。
適応の機能とは、それが解決するために進化した適応問題、つまり、それが生存や繁殖に正確にどのように貢献するかを指します。適応の機能は、通常、「特別な設計」の証拠によって特定され、確認されます。そこでは、構成要素または「設計上の特徴」がすべて、特定の適応問題を解決するために正確な方法で貢献します。適応の仮説上の機能を評価するための基準には、通常、効率性(問題を巧みな方法で解決すること)、経済性(問題を費用対効果の高い方法で解決すること)、正確性(すべての構成要素が特定の目的を達成するために専門化されていること)、および信頼性(それが機能するように設計された文脈で信頼できるパフォーマンスを発揮すること)が含まれます(Confer et al., 2010; Tooby & Cosmides, 1992, 2005; Williams, 1966を参照)。
各適応には、それ自身の進化の期間があります。最初は、突然変異、つまりDNAの一部におけるコピーエラーが、一個体で発生します。ほとんどの突然変異は生存や繁殖を妨げますが、一部は、偶然にも、生物が生き残り、繁殖するのを助けることになります。突然変異が、生物に集団の他のメンバーよりも繁殖上の利点を与えるのに十分役立つ場合、それはより多くの数で次の世代に受け継がれます。したがって、次の世代では、最初は一個人の突然変異であった特性を持つ個体がより多くなります。多くの世代にわたって、それが成功し続けるならば、突然変異は集団全体に広がり、したがって、種のすべてのメンバーがそれを持つことになります。
進化的に適応した環境(EEA)とは、適応を生み出す原因となった、適応の進化の期間中に起こった選択圧の統計的な複合体を指します(Tooby & Cosmides, 1992)。言い換えれば、各適応のEEAは、深い進化の時間をかけてそれを形成する原因となった選択力、つまり適応問題を指します。例えば、目のEEAは、数億年にわたる視覚系の多くの構成要素のそれぞれを形成した特定の選択圧を指します。二足歩行のEEAは、約440万年前に遡る、より短い時間スケールの選択圧を含みます。重要な点は、EEAが特定の時間や場所を指すのではなく、適応を形成する責任がある選択力を指すということです。したがって、各適応には独自のEEAがあります。適応の進化の期間とは、それが種の普遍的な設計を特徴づけるようになるまで、一つ一つ構築された時間スパンを指します。
適応は進化の主要な産物ですが、それだけが産物というわけではありません。進化の過程は、適応の副産物も生み出します。副産物は、適応問題を解決せず、機能的な設計を持たない特性です。それらは、電球からの熱が光のための設計の副産物であるように、機能的な設計を持つ特性とたまたま結合しているために、それらの適応と共に「運ばれる」のです。
人間のへそを考えてみましょう。へそ自体が、人間の生存や繁殖を助けるという証拠はありません。へそは、食物を捕まえたり、捕食者を検出したり、ヘビを避けたり、良い生息地を見つけたり、良い配偶者を選んだりするのに役立ちません。それは、直接的または間接的に適応問題の解決に関与しているようには見えません。むしろ、へそは、適応である何かの副産物です。つまり、成長する胎児に食物を提供していたへその緒です。したがって、何かが適応の副産物であるという仮説は、それが副産物である適応を特定し、その存在がその適応と関連している理由を特定する必要があります。
へそは適応ではありません。獲物を捕まえたり、捕食者を阻止したりするのに役立ちません。むしろ、それは適応であったものの副産物です。つまり、かつては胎児が母親から栄養を得るために機能していたへその緒です。
シャッターストック
進化プロセスの3番目で最後の産物は、ノイズまたはランダムな効果です。ランダムな効果は、突然変異、環境の突然かつ前例のない変化、または発達中の事故などの力によって生じる可能性があります。これらのランダムな効果は、機械に砂を投げ込んだり、コンピュータのハードドライブに熱いコーヒーをこぼしたりすると機能が損なわれるように、生物の円滑な機能を損なうことがあります。統合失調症として知られる精神障害は、正常な心理的機能を妨げるランダムな効果を生み出す高い突然変異負荷に起因する可能性があると主張する人もいます(Keller & Miller, 2006)。一部のランダムな効果は中立的であり、適応機能に貢献も妨害もせず、一部は生物に有益です。へその形は中立的である可能性が高いです。別の例として、電球のガラスの筐体は、材料の不完全さや製造プロセスに起因する滑らかさからの乱れを含むことがよくありますが、電球の機能には影響しません。電球は、そのような不完全さがあってもなくても等しく機能します。
要約すると、進化の過程は3つの産物、すなわち適応、適応の副産物、そしてランダムな効果を生み出す。原則として、私たちは種の構成部分を分析し、どれが適応で、どれが副産物で、どれが単にランダムな効果によるものかを決定するための研究を行うことができる。進化科学者たちは、これら3つのカテゴリーの産物の相対的な大きさの推定において意見が異なる。一部は、言語のような人間特有の性質でさえ、私たちの大きな脳の単なる偶発的な副産物であると信じている(Gould, 1991)。他の人々は、人間の言語が適応であるという圧倒的な証拠を見ている(Pinker, 1994)。幸いなことに、私たちは科学者の信念に頼る必要はない。なぜなら、私たちは彼らの考えを直接テストできるからだ。
進化産物の3つのカテゴリーの相対的な大きさに関する科学的な些細な違いにもかかわらず、すべての進化科学者は一つの基本的な点で同意している:適応は選択による進化の主要な産物である(Alcock, 2013; Dawkins, 1982; Dennett, 1995; Gould, 1997; Trivers, 1985; Williams, 1992)。スティーブン・ジェイ・グールドのような進化心理学の批評家でさえ、
適応の存在と中心的な重要性、あるいは自然選択による適応の生産のどちらも否定しません…自然選択以外に、これほど見事に機能する設計の構造を構築する実証された力を持つ科学的なメカニズムは他に知りません。
(グールド、1997年、pp. 53-58)
そして、人間を含むすべての動物の性質の核心は、多数の適応の集合から成ります。これらの適応の一部は、私たちの環境における適応的に関連する情報への窓を提供する感覚器官、つまり目、耳、鼻、味蕾です。これらの適応の一部は、直立した骨格の姿勢、脚の筋肉、大きなつま先など、私たちが環境を移動するのを助けます。進化心理学者は、人間性を構成する適応の特別なサブクラスの一つ、つまり心理的な適応に焦点を当てる傾向があります。
進化心理学における進化分析のレベル
科学の本質的な特徴の一つは、仮説の定式化です。進化心理学の場合、仮説の性質は通常、適応問題とその解決策を中心に展開します。より具体的には、私たちの祖先が直面した適応問題と、それらの問題に対する適応的な心理的解決策に焦点を当てます。進化心理学者がこれらの仮説をどのように正確に定式化するかを見るために、図2.1に示すように、進化心理学内の分析レベルの階層を説明する必要があります。
図2.1 進化分析のレベル
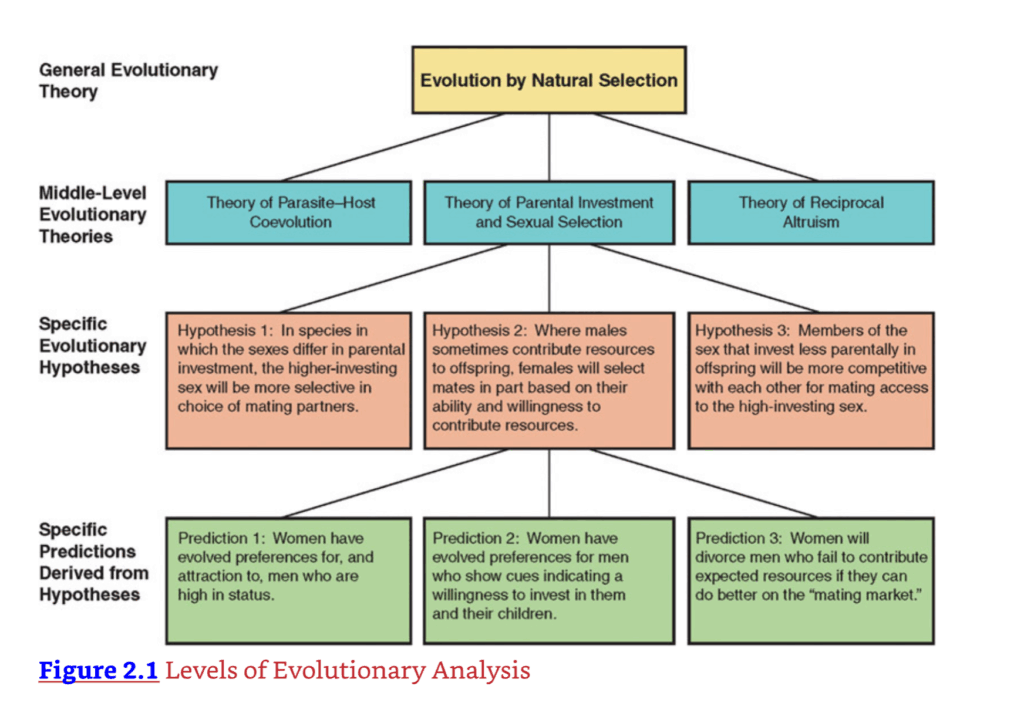
一般進化論
第一の分析レベルは、一般進化論です。その現代的な形式では、自然選択による進化は、「遺伝子の目」の視点から理解されます。つまり、示差的な遺伝子複製が、適応が形成される進化プロセスのエンジンです(Cronin, 2005; Dawkins, 1982, 1989; Hamilton, 1964; Williams, 1966)。もちろん、進化論は、第1章で説明したように、自然選択のプロセス以上のものを含みます。しかし、自然選択は、複雑な機能的設計を創造できる唯一の既知の基本的な因果過程であり、したがって、ここでは進化論的理論の階層の最も一般的なレベルとして扱われます。
この一般的なレベルでは、私たちは進化「論」について話しますが、それは生物科学者によって事実として広く受け入れられています。進化心理学の研究のほとんどは、進化論が正しいという仮定から進められていますが、その仮定を直接テストするものではありません。
原則として、一般進化論を反証できる観察がある。科学者が、自然選択が作用するには短すぎる期間(例えば、7日間)で創造された複雑な生命体を観察した場合、科学者が、ある種の適応が他の種の利益のためだけに機能することを発見した場合、科学者が、ある個体の適応が同性の競争相手の利益のためだけに機能することを発見した場合などである(Darwin, 1859; Mayr, 1982; Williams, 1966)。そのような現象はこれまで文書化されたことがない。
図2.1は、進化心理学における分析レベルの階層の一つのバージョンを示している。一般進化論は、階層の最上位を占める。各中レベルの理論は、一般進化論と矛盾してはならないが、そこから導き出されることはできない。進化した心理学的メカニズムや行動パターンに関する特定の進化仮説は、各中レベルの理論から導き出される。各特定の進化仮説は、様々な特定の検証可能な予測を生成することができる。各仮説と理論の支持は、経験的証拠の累積的な重みによって評価される。
中間レベルの進化論
一つ下のレベルに移動すると(図2.1参照)、トリヴァースの親の投資と性淘汰の理論のような中間レベルの理論が見つかります。これらの中間レベルの理論は、依然としてかなり広範で、機能の全領域をカバーしています。それらはまた、科学的検証の対象であり、偽であることが証明される可能性があります。この点を説明するために、一つの理論だけを調べてみましょう。それは、性淘汰の原動力としてのトリヴァースの親の投資理論です。ダーウィンの性淘汰理論(1871)を精緻化したこの理論は、配偶者選択と性内競争(同性間の競争)の操作を予測するための重要な要素の一つを提供しました。トリヴァースは、子孫により多くの資源を投資する性(しばしば、しかし常にではないが、雌)は、配偶者を選択する際に、より選択的または識別力を持つように進化すると主張しました。対照的に、子孫により少ない資源を投資する性は、より選択的でなく、貴重で高投資の異性への性的アクセスのために、同性のメンバーとより競争的になるように進化すると主張しました。
トリヴァースの理論の基本的な信条は、様々な種からの経験的証拠によって強く支持されています(Alcock, 2013; Dugatkin, 2020)。雌が雄よりも子孫に多く投資する多くの種では、雌は実際、配偶者選択においてより選択的である可能性が高いです。しかし、雄が雌よりも多く投資する種も少数あります。例えば、一部の種では、雌が雄に卵を産み付け、雄が生まれるまで子孫を運びます。モルモンコオロギ、ヤドクガエル、タツノオトシゴのような種では、例えば、雄はこのように雌よりも多く投資します(Josephs, Sellers, Newman, & Mehta, 2006; Trivers, 1985)。
雄のヨウジウオは、雌から卵を受け取り、カンガルーのような袋で運びます。これらの雌は、「最高の」雄をめぐって互いに激しく競争し、雄は誰と交尾するかについて選択的です。このいわゆる性役割逆転種は、トリヴァースの理論を支持しており、選択性の性差を引き起こすのは「雄らしさ」や「雌らしさ」自体ではなく、むしろ2つの性の相対的な親の投資であることを示しています。したがって、累積的な証拠の重みは、相対的な選択性と配偶者に対する競争性の決定要因としての親の投資に関するトリヴァースの中間レベルの理論を実質的に支持しています(Klug, Heuschele, Jennions, & Kokko, 2010も参照)。
図2.1をもう一度見てください。トリヴァースの中間レベルの理論は、一般進化論と両立することがわかります。彼は、進化の過程で生じ得ない何かを提案しているわけではありません。しかし、同時に、親の投資理論は、一般進化論から論理的に導き出されるものではありません。自然選択の理論には、親の投資について何も書かれていません。したがって、中間レベルの理論は、一般進化論と両立しなければなりませんが、それ自体の価値で評価されなければなりません。
特定の進化仮説
図2.1の1つ下のレベルに移動して、特定の進化仮説を調べてみましょう。例えば、人間に関して提唱されている一つの仮説は、女性が提供できる資源を持つ男性を特に好むように進化したというものです(Buss, 1989a; Symons, 1979)。その論理は次のとおりです。第一に、女性は子供に多大な投資をするため、配偶者を選ぶ際には選択的になるように進化した、というのが親の投資理論からの標準的な予測です。第二に、女性の選択の内容は、歴史的に彼女たちとその子供たちの生存と繁殖を向上させたものを反映しているはずです。したがって、女性は、自分たちとその子供たちに資源を貢献できる能力と意欲の両方を持つ男性を好む配偶者選好を進化させたと仮定されています。これは進化心理学的な仮説です。なぜなら、特定の心理的メカニズム、つまり配偶者選好の存在を提唱しており、それが特定の人間的な適応問題、すなわち彼女と彼女の子供たちに投資できると思われる配偶者を選ぶという問題を解決する機能を果たしているからです。
この特定の進化心理学的仮説は、経験的に検証することができます。科学者は、様々な文化圏の女性を研究し、彼女たちが実際に、自分たちとその子供たちに資源を貢献できる能力と意欲を持つ男性を好むかどうかを判断することができます。しかし、仮説を強力に検証するためには、それがどのような具体的な予測を生み出すかを見る必要があります。図2.1の階層の最下位レベルに移動します。女性が資源を提供する男性を好むという仮説に基づいて、次のような予測を立てることができます:(1)女性は、野心、社会的地位、知性、やや年上であることなど、資源の獲得と関連することが知られている特定の資質を男性に評価するでしょう。(2)インターネットの出会い系サイトでは、女性は資源を持っているように見える男性に、持っていないように見える男性よりも多く反応するでしょう。(3)ロマンチックなパートナーが経済的な資源を提供しない女性は、パートナーが経済的な資源を貢献する女性よりも、彼らと別れる可能性が高いでしょう。
これらの予測はすべて、女性が資源を持つ男性に対する特定の進化した嗜好を持つという仮説から導き出されます。仮説の価値は、それから導き出された予測の科学的検証にかかっています。もし予測が失敗した場合、つまり、女性が資源獲得に関連することが知られている人格特性を望んでいないこと、出会い系サイトでより多くの地位と資源を持つ男性に反応しないこと、そして資源を提供しない男性と別れる可能性が高くないことが示された場合、仮説は支持されません。もし予測が成功した場合、仮説は、少なくとも現時点では支持されます。
これはもちろん、非常に単純化されており、多くの場合、さらにいくつかの分析レベルが関与しています。私たちは、男性の投資を確保するという適応問題を解決できる、原則として、情報処理メカニズムのより詳細なタスク分析を実行することができます。そして、このタスク分析を、私たちの人間の祖先が利用できたであろう関連する祖先のてがかりを特定するためのガイドとして使用することができます。例えば、人間が進化の歴史の99パーセントを狩猟採集民として過ごしたことを知っているので(Tooby & DeVore, 1987)、女性の進化した嗜好の一部には、運動能力、優れた手と目の協調、長い狩りに必要な身体的持久力など、成功した狩猟に必要な特定の資質が含まれると予測することができます。
標準的な科学のすべての条件が当てはまります。予測が経験的に支持されない場合、それらの基盤となった仮説は疑問視されます。主要な仮説が予測の失敗によって疑問視される場合、仮説を生み出した中間レベルの理論の真実性や価値は疑わしくなります。一貫して支持される理論は、特に興味深く実り多い研究の道筋を生み出す場合、主要な中間レベルの理論として称賛されます。そのような道筋を生み出せない理論や、経験的に失敗する理論は放棄されます。
この分析レベルの階層は、次のような質問に答えるのに役立ちます:どのような証拠が進化論的な定式化を反証できるでしょうか?心理的なメカニズムに関する特定の仮説は、たとえその仮説につながった一つ上のレベルの理論が完全に正しくても、間違っている可能性があります。例えば、トリヴァースの中間レベルの親の投資理論は、女性が資源を持つ男性に対する特定の配偶者選好を進化させていないことが判明したとしても、正しい可能性があります。おそらく、女性の選好に関する関連する突然変異は発生しなかったのかもしれませんし、あるいは祖先の状況では、女性が自分自身の配偶者選択をすることが制約されていたのかもしれません(例えば、親が結婚を取り決めた場合)。
同様に、特定の進化心理学的仮説が正しいとしても、この場合、女性が資源を持つ男性に対する特定の配偶者選好を進化させてきたとしても、そこから導き出されるすべての予測が正しいという保証はありません。例えば、女性が男性の資源に関連する資質を望んでいても、それを提供しない男性とは別れないということもあり得ます。もしかしたら、夫が提供してくれない女性は、一部の文化では離婚を禁止する法律があるために、彼らと一緒に行き詰まっているのかもしれません。あるいは、女性が離婚すれば配偶者市場でそれほど良い条件を得られないと認識し、我慢することに決めるのかもしれません。これらの状況のいずれも、この特定の予測を誤ったものにする可能性があります。
重要な点は、進化論的な定式化の評価は、必ずしも単一の予測ではなく、証拠の累積的な重みに依存するということです。進化仮説は、正確に定式化されれば、非常に検証可能であり、そこから導き出された予測を証拠が支持しない場合には、反証される能力を十分に持っています(反証可能性の問題については、Ketelaar & Ellis, 2000を参照)。実際、一部の進化仮説は、男性の同性愛に関する血縁利他主義理論のように、証拠によって反証されています(Confer et al., 2010; この仮説の議論については第5章を参照)。
進化仮説の生成と検証の二つの戦略
図2.1の階層は、進化仮説と予測を生成するための1つの科学的戦略を示しています。この戦略は、仮説生成のトップダウンまたは理論駆動型アプローチと呼ばれます。一般進化論のトップから始めて、仮説を導き出すことができます。例えば、私たちは包括的適応度理論にのみ基づいて、人間が遠い遺伝的親族よりも近い遺伝的親族をより助けるだろうと予測することができます。あるいは、トリヴァースの中間レベルの親の投資理論に基づいて仮説を生成することができます。どちらの方法でも、導出は図を下に流れ、一般から特定へと進みます。
トップダウン戦略は、理論がいかに非常に有用であるかを示す一つの方法を例示しています。理論は、特定の仮説が生成される元となる一連の作業前提を提供します。それらはまた、血縁者や子供への投資のような重要な探究領域へと研究者を導くための枠組みを提供します。
進化心理学的仮説を生成するための第二の戦略があります(表2.2参照)。理論から始める代わりに、観察から始めることができます。ある現象の存在についての観察がなされたら、ボトムアップ方式で進み、その機能に関する仮説を生成することができます。人間は他者を鋭敏に知覚するため、通常、注意を向けるための正式な理論がなくても物事に気づきます。例えば、ほとんどの人は、人間が話し言葉でコミュニケーションし、二足で直立歩行し、時には他の集団と戦争をすることを教えてくれる理論を必要としません。一般進化論には、言語、二足歩行、または集団間の戦争が進化したという仮説を生み出すものは何もありません。
表2.2 進化仮説の生成と検証の2つの戦略
戦略2:観察駆動型または「ボトムアップ」戦略
戦略
例:親の投資理論から、女性は男性よりも子孫への義務的な投資が大きいため、女性は配偶者の選択においてより選択的または識別力がある傾向があるという仮説を導き出すことができます。
戦略1:理論駆動型または「トップダウン」戦略
ステップ1:既存の理論から仮説を立てる
ステップ1:既知の観察に基づいて適応機能に関する仮説を立てる
ステップ2:仮説に基づく予測を検証する 例:女性が男性の質とコミットメントを評価するためにセックスに同意する前に、より長い遅延とより厳しい基準を課すという予測を検証するための実験を行う。
ステップ3:経験的結果が予測を確認するかどうかを評価する
例:女性はセックスに同意する前に男性よりも長い遅延とより厳しい基準を課す(Buss & Schmitt, 1993; Kennair, Schmitt, Fjeldavli, & Harlem, 2009)。
例:A. 観察:男性は女性よりも配偶者の選択において外見をより重視するように見える。B. 仮説:女性の外見は、祖先の男性に繁殖力の手がかりを提供した。
ステップ2:仮説に基づく予測を検証する 例:男性の魅力の基準が女性の繁殖力の手がかりに密接に基づいているかどうかを判断するための実験を行う。
ステップ3:経験的結果が予測を確認するかどうかを評価する
例:男性は、既知の繁殖力との相関関係である低いウエスト対ヒップ比を魅力的だと感じる(Dixon, Crimshaw, Linklater, & Dixon, 2011; Singh, 1993)
私たちが自分自身や他の種について観察する多くのことが、進化論によって事前に予測されていなかったという事実は、その理論を損なうものではありません。しかし、それは一つの問題を引き起こします:これらの現象をどのように説明できるのでしょうか?進化論的思考は、私たちがそれらを理解するのに役立つでしょうか?
科学的研究によって文書化された一般的な観察を考えてみましょう:女性の外見は男性にとっての魅力の重要な部分である。これは、多くの人が何の科学的理論の指導もなしに観察することです。あなたのおばあちゃんでさえ、おそらくほとんどの男性が魅力的な女性を好むと教えてくれたでしょう。しかし、進化論的な視点はより深く探求します。それはなぜかと問います。
最も広く提唱されている進化論的仮説は、女性の外見が彼女の繁殖力に関する豊富な手がかりを提供するというものです(Sugiyama, 2005)。この仮説によれば、男性が魅力的だと感じるものは、繁殖力と関連する特定の身体的または行動的な特徴であるべきです。進化の過程で、これらの繁殖力の手がかりを示す女性に惹かれた男性は、繁殖力の手がかりを欠く女性に惹かれた男性よりも多くの子孫を残したでしょう。
心理学者のデヴェンドラ・シンは、そのような特徴の一つ、すなわちウエスト・ヒップ比(WHR)を提案しました(Singh, 1993)。ウエストがヒップよりも円周が小さいことを示す低いWHRは、2つの理由で繁殖力と関連しています。第一に、低いWHRを持つ不妊治療クリニックの女性は、高いWHRを持つ女性よりも早く妊娠します。第二に、高いWHRを持つ女性は、心臓病と内分泌系の問題の発生率が高く、どちらも低い繁殖力と関連しています。シンは、男性は低いWHRの女性を好み、この女性の繁殖力への物理的な手がかり、つまり祖先の男性が観察できたであろう手がかりへの欲求が男性に進化したと提案しました。
いくつかの異なる文化にわたる研究で、シンは様々なWHRの女性の線画を男性に提示した。一部は0.70のWHR(ウエストがヒップの10分の7のサイズ)、他は0.80のWHR、さらに他は0.90のWHRを示した。男性は最も魅力的だと思う図を丸で囲むように指示された。アフリカからブラジル、米国まで、様々な年齢の男性が、各文化において、0.70のWHRの女性を最も魅力的だと感じた。男性に女性の視覚的な画像を提示したアイトラッキング研究は、この体の領域が、乳房とともに、最も多くの初期の視覚的固視を受けることを確認しており、男性の砂時計体型の評価が非常に迅速かつ自動的に行われることを示唆している(Dixon et al., 2011)。別の研究では、先天的に失明している男性でさえ、より低いWHRを好むことがわかった(Karremans, Frankenhuis, & Arons, 2010)。男性が女性の外見を重視するという考えは一般的な観察ですが、この現象がなぜ起こるのかについて、特定の進化仮説を生成し、検証することができます。この場合、観察可能な繁殖力の手がかりのためです。
この「ボトムアップ」の仮説生成と検証の戦略について、2つの結論を導き出すことができます。第一に、科学者が現象を観察し、その後その起源と機能に関する仮説を立てることは、完全に正当なことです。例えば、天文学では、膨張する宇宙の発見が最初に観察され、その後それを説明しようとする理論が続きました。
ボトムアップ戦略は、存在するかもしれないがまだ文書化されていない現象に関する「トップダウン」の理論主導の仮説をうまく補完します。
第二に、進化仮説の価値は、その精度に一部依存します。仮説がより正確であればあるほど、そこから導き出される具体的な予測を生成しやすくなります。これらの予測は、ほとんどの場合、仮説が正しい場合に仮説上の適応が持つべき「設計上の特徴」の分析に基づいています。一歩一歩、予測ごとに、経験的に検証された予測を生み出さない仮説は破棄されます。私たちは、一貫して経験的に検証された予測を生み出す仮説を保持します。したがって、科学が、進化した心理的メカニズムの存在、複雑さ、機能性の発見に近づくにつれて、企業全体が累積的な性質を示します。
人間性の核心:進化した心理学的メカニズムの基礎
このセクションでは、進化心理学の観点から人間性の核心について説明します。まず、人間を含むすべての種は、記述し説明できる性質を持っています。第二に、人間性を構成する中核的な単位である進化した心理学的メカニズムの定義を提供します。最後に、進化した心理学的メカニズムの重要な特性を検討します。
すべての種には性質がある
ライオンの性質は、四本足で歩き、大きな毛皮のたてがみを生やし、他の動物を狩って食べることの一部です。蝶の性質は、飛ばない蛹の状態に入り、繭に身を包み、食べ物や配偶者を求めて優雅に舞い上がることの一部です。ヤマアラシの性質は針で身を守ること、スカンクの性質は悪臭で身を守ること、雄鹿の性質は角で身を守ること、そしてカメの性質は甲羅で身を守ることです。すべての種には性質があり、その性質は各種で異なります。各種は、その進化の歴史の中でいくらか独特の選択圧に直面してきたため、したがって、いくらか異なる一連の適応問題に直面してきました。
人間もまた、私たちをユニークな種として定義する性質を持っており、すべての心理学理論はその存在を暗示しています。ジークムント・フロイトにとって、人間性は荒れ狂う性的および攻撃的な衝動から成っていました。ウィリアム・ジェームズにとって、人間性は数十または数百の本能から成っていました。B.F.スキナーの急進的行動主義の理論のような、最も熱心な環境主義者の理論でさえ、人間には性質があると仮定しています。この場合、それはいくつかの非常に一般的な学習メカニズムから成っています。すべての心理学理論は、その核心に、人間性に関する基本的な前提を必要とします。
選択による進化は、その人間性の基本的な適応的構成要素を生み出すことができる唯一の既知の因果過程であるため、すべての心理学理論は、暗黙的または明示的に進化的である。もし人間が性質を持ち、選択による進化がその性質を生み出した因果過程であるならば、次の質問はこうなる:私たちの進化の起源を調べることによって、人間性に関するどのような偉大な洞察が得られるだろうか?進化の過程を調べることは、人間のケースにおけるその過程の産物について何か教えてくれるだろうか?これらの重要な問いに対する答えが、本書の残りの部分の核心を形成する。
進化生物学のより広範な分野は、生物のすべての統合された部分の進化的分析に関心があるのに対し、進化心理学は、心理的な部分、つまり、進化した情報処理メカニズムの集合としての人間精神の分析、それらのメカニズムを活性化する文脈、およびそれらのメカニズムによって生成される行動に、より狭く焦点を当てる傾向があります。そして今、私たちは、人間の心を構成する適応のクラス、つまり進化した心理学的メカニズムに直接目を向けます。
進化した心理学的メカニズムの定義
進化した心理学的メカニズムとは、以下の特性を持つ生物内のプロセスの集合です。
- 進化した心理学的メカニズムは、進化の歴史を通じて再発的に生存または繁殖の特定の問題を解決したため、その形で存在します。これは、メカニズムの形態、その設計上の特徴のセットが、特定の錠に合うように作られた鍵のようなものであることを意味します。鍵の形状が錠の内部の特徴に合うように調整されなければならないように、心理的メカニズムの設計上の特徴の形状は、生存または繁殖の適応問題を解決するために必要な特徴と調整されなければなりません。適応問題との整合に失敗することは、進化の選択的なふるいを通過することに失敗することを意味しました。
- 進化した心理的メカニズムは、情報の狭い範囲のみを取り込むように設計されています。人間の目を考えてみましょう。目を開けてほとんどすべてを見ているように見えますが、目は実際には、電磁波の広範なスペクトルからの入力の狭い範囲、つまり可視スペクトル内のものにのみ敏感です。私たちは、可視スペクトルよりも短いX線を見ません。また、より長い電波も見ません。可視スペクトル内でさえ、私たちの目は、より狭いサブセットの情報を処理するように設計されています(Marr, 1982; van der Linde, Rajashekar, Bovik, & Cormack, 2009)。人間の目には、物体からの対照的な反射を拾う特定のエッジ検出器と、動きを拾う運動検出器があります。また、物体の色に関する情報を拾うように設計された特定の錐体もあります。目は、万能の見る装置ではありません。それは、潜在的な情報のより大きな領域の中から、特定の周波数範囲内の波、エッジ、運動など、情報の狭いサブセットのみを処理するように設計されています。同様に、ヘビを恐れることを学ぶ素因の心理的メカニズムは、自己推進する細長い物体からの滑りやすい動きという、情報の狭い範囲のみを取り込むように設計されています。食物、風景、配偶者に対する私たちの進化した好みはすべて、潜在的に入力となりうる無限の配列の中から、限られたサブセットの情報のみを取り込むように設計されています。各メカニズムを活性化する限られた手がかりは、EEA中に再発した手がかり、または現代の環境でこれらの祖先の手がかりを密接に模倣する手がかりです。
- 進化した心理的メカニズムの入力は、生物にそれが直面している特定の適応問題を伝えます。滑りゆく蛇を見るという入力は、あなたが特定の生存問題、すなわち、噛まれた場合の身体的損傷、そしておそらくは死に直面していることを伝えます。潜在的に食べられる物体の異なる匂い(腐敗して腐ったもの対甘くて香ばしいもの)は、あなたが食物選択という適応的な生存問題に直面していることを伝えます。要するに、入力は、生物がどの適応問題に対処しているかを知らしめます。これはほとんど常に意識の外で起こります。人間は、焼きたてのピザの匂いを嗅いで、「ああ、私は食物選択の適応問題に直面している!」とは考えません。代わりに、匂いは無意識のうちに食物選択メカニズムを誘発し、適応問題の認識は必要ありません。
- 進化した心理的メカニズムの入力は、決定規則または手順を通じて出力に変換されます。蛇を見ると、それを攻撃するか、逃げるか、凍りつくかを決めることができます。オーブンから出したばかりのピザの匂いを嗅ぐと、それをむさぼり食うか、立ち去るかを選ぶことができます(おそらくダイエット中であれば)。決定規則は、生物をある道筋から別の道筋へと導くための一連の手順、つまり「もし…ならば」という文です。例えば、怒っているライバルと公に直面した場合、人間は次のような「もし…ならば」という決定規則を持っているかもしれません。「もし怒っているライバルがより大きく強いならば、物理的な戦いを避けなさい。もし怒っているライバルがより小さく弱いならば、公の挑戦を受け入れて戦いなさい。」この例では、入力(特定の大きさの怒っているライバルによる対決)は、決定規則(「もし…ならば」という手順)を通じて出力(戦うか逃げるかの行動)に変換されます(図2.2参照)。
図2.2 進化した心理学的メカニズム:情報処理適応としての進化した心理学的メカニズムの現代的定式化
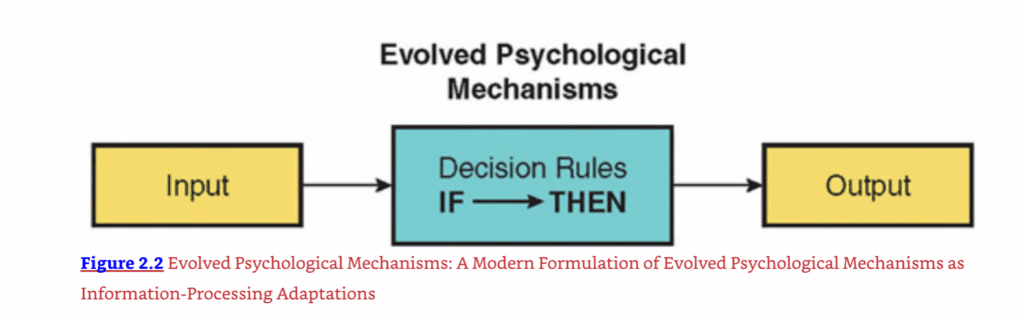
- 進化した心理的メカニズムの出力は、生理学的活動、他の心理的メカニズムへの情報、または顕在的な行動であり得る。蛇を見ると、生理学的に興奮したり恐怖を感じたりするかもしれない(生理学的出力)。この情報を使って、凍りつくか逃げるかといった行動の選択肢を評価するかもしれない(他の心理的メカニズムへの情報)。あるいは、この評価を行動、例えば逃げること(行動的出力)に使うかもしれない。
別の例を考えてみよう:性的嫉妬。あなたがロマンチックなパートナーとパーティーに行き、飲み物を取りに部屋を出たとしよう。戻ってくると、あなたのパートナーが別の人と生き生きと話しているのを目にする。彼らは互いに非常に近くに立っており、互いの目を深く見つめ、軽く触れ合っていることに気づく。これらの手がかりは、性的嫉妬と呼べる反応を引き起こすかもしれない。これらの手がかりは、メカニズムへの入力として機能し、パートナーを失う脅威という適応問題をあなたに知らせる。この入力は、一連の決定規則に従って評価される。一つの選択肢は、彼らを無視して無関心を装うことである。別の選択肢は、ライバルを脅すことである。第三の選択肢は、激怒してライバルを殴ることである。さらに別の選択肢は、あなたの関係を再評価することであろう。したがって、心理的メカニズムの出力は、生理学的(覚醒)、行動的(対決、脅迫、殴打)、または他の心理的メカニズムへの入力(あなたの関係の状態の再評価)であり得る。 - 進化した心理的メカニズムの出力は、特定の適応問題の解決に向けられています。パートナーの潜在的な不貞の手がかりが適応問題の存在を知らせるように、性的嫉妬メカニズムの出力は、その問題を解決することに向けられています。脅されたライバルは現場を離れるかもしれません。あなたのロマンチックなパートナーは、他の人と浮気することを思いとどまるかもしれません。あるいは、あなたの関係の再評価は、あなたが損失を切り捨てて前に進むことを引き起こすかもしれません。これらのいずれも、あなたの適応問題の解決に役立つ可能性があります。心理的メカニズムの出力が特定の適応問題の解決につながると述べることは、その解決が常に成功することを意味するものではありません。ライバルは、あなたの脅しによって思いとどまらないかもしれません。あなたのパートナーは、あなたの嫉妬の表示にもかかわらず、あなたのライバルと浮気するかもしれません。重要な点は、心理的メカニズムの出力が常に成功した解決につながるということではなく、むしろ、それが進化した環境において、メカニズムの出力が平均して競合する戦略よりも適応問題をより良く解決する傾向があるということです。
心に留めておくべき重要な点は、過去の進化で成功した解決策につながったメカニズムが、現在成功した解決策につながるとは限らないということです。例えば、脂肪と砂糖に対する私たちの強い味の好みは、肉、卵、ナッツからの脂肪と、熟した果物や珍しい蜂蜜からの砂糖が貴重で乏しいカロリー源であった私たちの進化の過去において、明らかに適応的でした。しかし、今では、ピザ屋がパイや砂糖を多く含んだソフトドリンクを街角の至る所で販売しており、脂肪と砂糖はもはや乏しい資源ではありません。これらの物質に対する私たちの強い味は、今や脂肪と砂糖の過剰摂取を引き起こし、それが動脈硬化、2型糖尿病、心臓発作につながり、私たちの生存を損なう可能性があります。進化心理学者は、これを祖先の環境と現代の環境の間のミスマッチと呼んでいます(Li, van Vugt, & Colarelli, 2217を参照)。本書では後で他の進化的ミスマッチを検討しますが、中心的な点は、進化したメカニズムは、それらが進化した期間中に平均して成功につながったために、それらの形で存在しているということです。それらが現在適応的であるかどうか、つまり、それらが現在、生存と繁殖の増加につながるかどうかは、ケースバイケースで決定されなければならない経験的な問題です。
要約すると、進化した心理的メカニズムとは、特定の情報のサブセットを取り込み、その情報を手順と決定規則を介して、歴史的に適応問題の解決に役立ってきた出力に変換するように設計された、生物内のプロセスの集合です。心理的メカニズムは、その生物の祖先にとって特定の適応問題の成功した解決に平均してつながったため、現在の生物に存在します。
進化した心理学的メカニズムの重要な特性
このセクションでは、進化した心理学的メカニズムのいくつかの重要な特性を検討します。それらは、「心をその自然な関節で切り分ける」ための非恣意的な基準を提供し、問題特異的で、数が多く、複雑である傾向があります。これらの特徴は組み合わさって、現代人を特徴づける行動の途方もない柔軟性を生み出します。
進化した心理学的メカニズムは、「心をその関節で切り分ける」ための非恣意的な基準を提供する
進化心理学の中心的な前提の一つは、心理的メカニズムを特定し、記述し、理解するための主要な非恣意的な方法は、その機能、つまり選択によって解決するように設計された特定の適応問題を明確にすることである。
人間の体を考えてみましょう。原則として、体のメカニズムは無限の方法で説明することができます。なぜ解剖学者は、肝臓、心臓、手、鼻、目を別々のメカニズムとして識別するのでしょうか?答えは機能です。肝臓は、心臓や手によって実行される機能とは異なる機能を実行するメカニズムとして認識されています。目と鼻は、近くに位置していますが、異なる機能を実行し、異なる入力(視覚スペクトル内の電磁波対匂い)に従って動作します。解剖学者が目と鼻を一つのカテゴリーにまとめようとすると、それは滑稽に見えるでしょう。体の構成部分を理解するには、機能の特定が必要です。機能は、これらの構成部分を理解するための非恣意的な方法を提供します。
進化心理学者は、心のメカニズムを理解するためにも同じ原則が使用されるべきだと信じています。心は無限の方法で分割することができますが、そのほとんどは恣意的なものです。人間の心の強力な非恣意的な分析は、機能に基づいています。心の2つの構成要素が異なる適応機能を実行する場合、それらは別々のメカニズムと見なすことができますが、肺と心臓が特定の方法で互いに相互作用するように、重要で興味深い方法で互いに相互作用する可能性があります。
進化した心理学的メカニズムは、問題に特化する傾向がある
ニューヨーク市からカリフォルニア州サンフランシスコの特定の番地までの道順を誰かに教えてあげると想像してみてください。「西へ向かってください」のような一般的な指示を与えた場合、その人はテキサスほど南、あるいはアラスカほど北に行き着くかもしれません。一般的な指示では、正しい州に確実に着くことはできず、ましてや正しい番地にたどり着くことはできません。
さて、その人が正しい州に着いたと仮定しましょう。「西へ向かう」という指示は、カリフォルニアの西は海なので、ほとんど役に立たないでしょう。一般的な指示は、カリフォルニア州内の正しい都市、ましてや正しい番地にたどり着くための何の指針も提供しません。その人を正しい州、都市、通り、そしてその通りの場所まで導くには、より具体的な指示を与える必要があります。特定の番地に行く方法はたくさんありますが、いくつかの道は他の道よりもはるかに効率的で時間を節約できます。
国の反対側にある特定の番地を探すことは、特定の適応的解決策に到達するために必要なことの良い例えです。番地のように、適応問題は具体的です。その蛇に噛まれないようにする、流水と隠れる場所のある生息地を選ぶ、毒物や寄生虫を含む食べ物を避ける、繁殖力のある配偶者を選ぶなどです。一般的な適応問題というものはありません(Symons, 1992)。
適応問題が具体的であるため、その解決策も具体的になる傾向があります。一般的な指示が正しい場所にたどり着かせないように、一般的な解決策は正しい適応的解決策にたどり着かせません。2つの適応問題を考えてみましょう。正しい食物を選ぶこと(生存問題)と、子供を持つための正しい配偶者を選ぶこと(繁殖問題)です。2つの問題で「成功した解決策」と見なされるものは大きく異なります。成功した食物選択には、カロリー、特定のビタミン、ミネラルを持ち、有毒物質を含まない物体を特定することが含まれます。成功した配偶者選択には、通常、とりわけ、繁殖力があり、良い親になるパートナーを特定することが含まれます。
これらの2つの選択問題に対する一般的な解決策とはどのようなもので、それらを解決するのにどれほど効果的でしょうか?一つの一般的な解決策は、「最初に来たものを選ぶ」ことです。これは、有毒植物を食べたり、不妊の人と結婚したりすることにつながる可能性があるため、悲惨なことになるでしょう。人間の進化の歴史の中で、誰かがこれらの適応問題に対してそのような一般的な解決策を実行したとしたら、彼らは祖先になることに失敗したでしょう。
これらの選択問題を解決するためには、食物と配偶者の重要な資質に関するより具体的なガイダンスが必要です。例えば、新鮮で熟したように見える果物は、腐っていて青カビに覆われた果物よりも良い栄養素を示すでしょう。若くて健康そうに見える人々は、年老いて病気に見える人々よりも平均して繁殖力が高いでしょう。これらの選択問題を成功裏に解決するためには、私たちの意思決定メカニズムの一部である特定の選択基準、つまり資質が必要です。
メカニズムの特異性は、エラーによってさらに示されます。食物の選択でエラーを犯した場合、そのエラーを修正する機能を果たす一連の適応を持っています。悪い食べ物を一口噛むと、ひどい味がするかもしれず、その場合は吐き出します。味覚を通り過ぎてしまった場合は、吐き気を催すかもしれません。そして、胃まで到達してしまった場合は、嘔吐するかもしれません。これは、有害な摂取物を排出するように設計された特定のメカニズムです。しかし、配偶者の選択でエラーを犯した場合、吐き出したり、吐き気を催したり、吐いたりしません(少なくとも通常は)。関係を離れたり、他の人に乗り換えたりすることで、他の方法でエラーを修正します。
要約すると、適応メカニズムの問題特異性は、一般性よりも好まれる傾向があります。なぜなら、(1)一般的な解決策は生物を正しい適応的解決策に導くことに失敗する、(2)たとえ機能したとしても、一般的な解決策はあまりにも多くのエラーにつながり、したがって生物にとってコストがかかる、(3)「成功した解決策」を構成するものは問題ごとに異なるからです。
人間は多くの進化した心理学的メカニズムを持っている
人間は、多くの生物と同様に、多くの適応問題に直面しています。生存の問題だけでも、数十から数百に及びます。体温調節の問題(寒すぎたり暑すぎたりすること)、捕食者や寄生虫を避けること、生命維持に必要な食物を摂取すること、高所からの落下を避けること、危険な見知らぬ人から遠ざかることなどです。そして、良い配偶者を選び、惹きつけ、維持し、悪い配偶者を追い払うという交配の問題があります。また、授乳、離乳、社会化、避難所、保護、そして異なる子供の様々なニーズに対応するという育児の問題もあります。そして、兄弟、姉妹、甥、姪のような親族に投資する問題、社会的な対立に対処する問題、攻撃的な集団から身を守る問題、そして社会的地位の階層と格闘する問題があります。
FIG BRAIN
特定の問題には特定の解決策が必要であるため、多数の特定の問題には多数の特定の解決策が必要になります。私たちの体には、血液を送り出す心臓、酸素を取り込む肺、毒素をろ過する肝臓など、何千もの特定のメカニズムが含まれているように、この分析によれば、心にも数百または数千の特定のメカニズムが含まれている必要があります。多数の異なる適応問題は、わずか数個のメカニズムでは解決できないため、人間の心は多数の進化した心理学的メカニズムで構成されている必要があります。
進化した心理学的メカニズムの特異性、複雑性、および多数性は、人間に行動の柔軟性を与える
心理学的メカニズムの定義は、入力、決定規則、出力という重要な構成要素を含み、適応がなぜ行動に必ずしも現れる硬直した「本能」ではないのかを浮き彫りにします。皮膚の下の構造を保護するために進化した、たこを生成するメカニズムの例を思い出してください。あなたは、繰り返しの摩擦を経験しないように環境を設計することができます。この場合、あなたのたこを生成するメカニズムは活性化されません。メカニズムの活性化は、環境からの入力に依存します。同様に、すべての心理学的メカニズムは、その活性化のために入力を必要とします。
心理学的メカニズムは、もう一つの重要な理由から、硬直した本能のようではありません。それは決定規則です。決定規則は、「もし蛇がシューッと音を立てたら、命からがら逃げろ」あるいは「もし私が惹かれている人が興味を示したら、微笑んで距離を縮めろ」といった「もし…ならば」という手順です。ほとんどのメカニズムでは、これらの決定規則は少なくともいくつかの可能な応答オプションを許容します。 deadly snake(致命的な蛇)に遭遇するという単純なケースでさえ、棒で攻撃するか、凍りついてそれが去ることを願うか、命からがら逃げるかという選択肢があります。
大工の道具箱を考えてみてください。大工は、切る、突く、鋸で引く、ねじ込む、ねじる、レンチで締める、平らにする、バランスをとる、金槌で叩くことができる「非常に一般的な道具」を一つ持つことによって柔軟性を得るわけではありません。代わりに、大工は道具箱の中に非常に多くの非常に特定の道具を持つことによって柔軟性を得ます。これらの非常に特定の道具は、一つの非常に「柔軟な」道具では不可能であろう多くの組み合わせで使用することができます。実際、「一般的な大工の問題」というものはないので、「一般的な」道具がどのようなものか想像することさえ困難です。同様に、人間は、多数の複雑で、特定の、機能的な心理的メカニズムを持つことから柔軟性を得ます。
心に新しいメカニズムが追加されるたびに、生物は新しいタスクを実行できるようになります。鳥には歩くことを可能にする足があります。翼を追加すると飛ぶことができます。鳥に嘴を追加すると、種子や木の実の殻を割って食用の中身にたどり着くことができます。新しい特定の適応が追加されるたびに、鳥はその適応がなければできなかった新しいタスクを達成できます。足と翼の両方を持つことで、鳥は歩くことと飛ぶことの両方の柔軟性を得ます。
これは、私たちの直感に反する結論につながります。私たちのほとんどは、多くの生得的なメカニズムを持つことは、行動を柔軟でなくさせると考えています。しかし、実際には、全く逆のことが真実です。私たちが持っているメカニズムが多ければ多いほど、私たちが実行できる行動の範囲は広くなり、したがって、私たちの行動の柔軟性は大きくなります。
領域特異的な心理学的メカニズムを超えて
前述のすべての議論は、人間が、特定の適応問題を解決することに特化した、多数の特殊な心理的メカニズムを持っていなければならないことを示唆しています。この結論は、進化心理学の分野で広く受け入れられており、実際、すべての種に対する進化的アプローチの基礎となっています(Alcock, 2013)。ある進化心理学者が述べたように、
単一の一般的な物質が、ある程度の専門化なしに、奥行きを見て、手を制御し、配偶者を引き付け、子供を育て、捕食者を避け、獲物を出し抜くなどできるという考えは、信じがたい。脳がその「可塑性」のためにこれらの問題を解決すると言うことは、魔法によって解決すると言うのと大差ない。
(ピンカー、2002年、75ページ)
しかし、一部の進化心理学者は、これらの特定のメカニズムに加えて、人間はいくつかの領域一般のメカニズムも進化させてきたと主張しています(例:Chiappe & MacDonald, 2005; Figueredo, Hammond, & McKiernan, 2006; Geary & Huffman, 2002; Livingstone, 1998; Mithen, 1996; Premack, 2010)。提案されている一般的なメカニズムの例としては、一般知能、概念形成、類推的推論、ワーキングメモリ、古典的条件付けがあります(第1章参照)。
領域一般のメカニズムの支持者たちは、適応問題の再発する特徴が特殊な適応を選択する一方で、人間は特殊な適応が進化するのに十分な規則性で再発しなかった多くの新しい問題に直面してきたと主張しています。さらに、人間は古代の適応問題を非常に斬新な方法で日常的に解決していることがわかっています。例えば、オンラインの食品配達サービスから食物を得たり、インターネットから配偶者を得たり、金物屋から道具を得たりすることができます。誰もが、人間が進化してきた環境とは非常に異なる多様な環境で繁栄してきたことを認識しています。「更新世から遠く離れた絶えず変化する世界」(Chiappe & MacDonald, 2005, p. 6)。Chiappe と MacDonald(2005)は、一般知能のような領域一般のメカニズムが、まさに「進化的目標を達成する上での非再発的問題の解決を可能にするため」(2005, p. 3)、あるいは古い問題に対する新しい解決策を開発するために進化したと提案しています。
彼らの議論の中心的な趣旨は、人類の進化の歴史において、人間は急速に変化する環境、つまり気候の予測不可能な変化、寒い氷河期と暖かい天候の間の変動、火山や地震による急速な変化などに対処することを余儀なくされたということです。同様に、Geary と Huffman (2002) は、人類の進化の歴史を通じて多くの情報パターンは非常に変動性が高かったため、経験に対して開かれた、より一般的な心理的メカニズムの進化を支持した可能性があることを示唆しています (Geary, 2009 も参照)。領域一般のメカニズムは、これらの理論家が提案するように、新規性、予測不可能性、および変動性に対処するために必要となるでしょう。興味深いことに、Kanazawa (2003b) も同様の議論を展開していますが、「一般知能」は実際には、狭いクラスの問題、つまり進化的に新しい問題を解決するように設計された領域特異的な適応であると提案しています。
一部の進化心理学者は、真に領域一般のメカニズムが進化できるかどうかについて懐疑的なままである(例:Cosmides & Tooby, 2002)。人々がインターネットをサーフィンしたり、車を運転したりするような進化的に新しいタスクを実行できるからといって、これらのタスクを実行することを可能にする適応が、それ自体が領域一般であることを必ずしも意味するわけではない。その点に関して、グリズリーベアに自転車に乗るように訓練したり、イルカに音楽に合わせて体を揺らすように訓練したりできるからといって、これらの新しい行動を可能にする適応が領域一般であることを意味するわけではない。進化心理学の科学のこの時点では、人間が特定のメカニズムに加えて、より領域一般のメカニズムを持っているかどうかについて、確固たる結論を導き出すのは時期尚早である。明らかなことは、領域特異性の仮定が、人間の心の重要なメカニズムを発見するために成功裏に使用されてきたということである。本書の以降の章では、これらの科学的成功を文書化する。領域一般のメカニズムの前提に基づく研究プログラムによって、同等の経験的発見がなされるかどうかは、未解決の問題である。
しかし、明らかなことは、人間の心が、互いに完全に壁で仕切られた、完全に孤立した別々のメカニズムだけで構成されているはずがないということです。選択は、様々な組み合わせや順列でうまく連携する、機能的に特殊化されたメカニズムを支持します。適応は、いわば「互いに話し合う」のです。例えば、一部のメカニズムから得られたデータは、他のメカニズムに情報を提供します。視覚、嗅覚、内的な空腹感からの情報がすべて、食物の可食性に関する決定規則への入力となる場合などです。この意味で、進化心理学者は、時々「モジュール性」の概念を援用する際に使用されるように、「情報カプセル化」を進化した心理学的メカニズムの定義的な特徴とはしない傾向があります(Hagen, 2005)。情報カプセル化という特性は、心理学的メカニズムが自己完結型の情報にのみアクセスでき、他の心理学的メカニズムの情報にアクセスできないことを意味します。
さらに、人間は、他のメカニズムを調節する機能を果たす、上位のメカニズムも進化させてきた可能性があります。森の中を歩いているときに、突然、空腹のライオン、熟したベリーでいっぱいの茂み、そして魅力的な潜在的な配偶者に遭遇したと想像してみてください。あなたは何をしますか?あなたは、ベリーと潜在的な配偶者をあきらめてでも、まずライオンを避けることを選ぶかもしれません。もしあなたが餓死寸前であれば、ライオンから逃げる前に、ベリーを手に入れるチャンスを狙うことを選ぶかもしれません。進化した心理学的メカニズムは、明らかに複雑な方法で互いに相互作用します。それらは、完全には理解されていない様々なシーケンスでオンになったりオフになったりします。人間が進化した上位の調節メカニズムを持っている可能性は、有望なままであり、将来の研究を待っています。
学習、文化、そして進化した心理学的メカニズム
進化した心理的メカニズムが仮定されるときに生じる一般的な質問は、次のようなバリエーションのものです:私たちが観察する人間の行動は、進化ではなく、学習と文化によって引き起こされるのではありませんか?人間の行動は、自然ではなく、育成の産物ではありませんか?これらの質問に答えるためには、心理的適応を援用する説明の正確な形式と、学習と文化を援用する説明の形式を注意深く分析する必要があります。
まず、進化心理学の枠組みは、「生まれか育ちか」、「生得的か学習的か」、そして「生物学的か文化的か」といった二分法を解消します。進化した心理的メカニズムの定義に戻ると、(1) 深い時間にわたる再発性の選択圧を特徴とする環境が各メカニズムを形成した、(2) 人の発達中の環境からの入力が各メカニズムの出現に必要である、そして (3) 環境からの入力が各メカニズムの活性化に必要であることに気づくでしょう。したがって、たこや嫉妬行動が「進化した」のか「学習した」のかを問うことは意味がありません。「進化した」は「学習した」の反対ではありません。すべての行動は、因果連鎖の各段階で環境からの入力と組み合わさった、進化した心理的メカニズムを必要とします。
次に、何かが学習されたと言うことは、正確には何を意味するのかを尋ねてみましょう。心理学で一般的に使用されるように、「学習」を説明として援用することは、単に環境からの入力の結果として生物に何らかの変化が起こったという弱い主張にすぎません。もちろん、人間は学習します。彼らは環境や文化の影響を受けます。しかし、学習には、学習を可能にする脳の構造、つまり進化した心理学的メカニズムが必要です。「結局のところ、3ポンドのカリフラワーは学習しませんが、3ポンドの脳は学習します」(Tooby & Cosmides, 2005, p. 31)。説明上の課題は、単に行動に「学習」というラベルを貼るだけでは十分ではありません。私たちは、環境からの入力の結果として人間の行動を変えることを可能にする、根底にある学習メカニズムの性質を特定する必要があります。
さて、これらの学習メカニズムの性質とはどのようなものでしょうか?3つの具体的な例を考えてみましょう:(1)人々は近親者との性交渉を避けることを学びます(学習された近親相姦回避)。(2)人々は毒素を含む可能性のある食物を避けることを学びます(学習された食物嫌悪)。(3)人々は、どの行動が社会的地位と威信を高めるかを、地域の文化から学びます(学習された威信基準)。これらの学習形態のそれぞれが、異なる進化した学習メカニズムによって最もよく説明されるという説得力のある証拠があります。近親相姦回避の適応問題を解決するには、性交渉をしてはならない個人のクラス、つまり近親者について学ぶ必要があります。人々はこれらの個人が誰であるかをどのように学ぶことができるでしょうか?進化した近親相姦回避学習メカニズムは、信頼できる親族の手がかり、つまり一緒に育った人々を使用することによって機能します。小児期における異性のメンバーとの同居期間は、性的魅力の欠如、そして実際に、彼らと性交渉をすることに対する人々の嫌悪感の量を強力に予測します(Lieberman, Tooby, & Cosmides, 2003)。
さて、学習された食物嫌悪を考えてみましょう。私たちは、特定の食物を摂取した後に吐き気を感じるメカニズムを通じて、食物嫌悪を学びます。キノコやレバー、魚が非常に嫌いな人は、通常、以前にそのような食物を摂取した後に病気になった経験があります。
最後に、地域の文化においてどの手がかりが地位や威信と結びついているかをどのように学ぶか考えてみましょう。狩猟採集社会では、優れた狩猟技術が威信につながります。学術界では、他の学者から多く引用される著名な出版物を持つ個人が高い威信を得ます。教授が教員会議に死んだヘラジカを叩きつけても、地位が上がる可能性は低いでしょう。他の地域の文化では、タトゥーの数、オートバイの大きさとメーカー、あるいはギターやビデオゲームを演奏するスキルが高い威信と関連しています。人々は、部分的には注意構造に焦点を当てることによって威信基準を学びます。つまり、地位の高い人々は、通常、最も多くの人々が最も注意を払う人々です(Chance, 1967)。他の人々が最も注意を払う人々の資質、服装のスタイル、行動に注意を払う(そしてしばしば模倣しようとする)ことによって、私たちは地域の文化の威信基準を学びます(Atkisson & O’Brien, 2012)。
これら3つの学習形態、すなわち近親相姦回避、食物嫌悪、威信基準は、機能するために明らかに異なる進化した学習メカニズムを必要とする。各学習形態は、発達中の同居、食物摂取と対になった吐き気、そして注意構造といった、異なる一連の手がかりからの入力に基づいて作動する。それぞれは、遺伝的親族への性的魅力の欠如、特定の物質の光景や匂いに対する嫌悪感、そして他者が注目している人々への注意といった、異なる機能的な出力を持つ。そして重要なことに、各学習形態は異なる適応問題を解決する。
この分析から導き出せる3つの重要な点があります。第一に、何かに「学習された」というレッテルを貼ることは説明を提供するのではなく、単に環境からの入力が生物を何らかの形で変化させるという記述にすぎません。第二に、「学習された」と「進化した」は競合する説明ではなく、むしろ、学習には進化した心理的適応が必要です。第三に、進化した学習メカニズムはしばしば本質的に特異的です(文化の進化心理学に関する詳細な議論については第13章を参照)。
進化仮説を検証する方法
進化仮説を検証する方法
- 異なる種を比較する
- 異文化間の方法
- 生理学的および脳画像法
- 遺伝学的方法
- 男性と女性を比較する
- 種内の個体を比較する
- 異なる文脈で同じ個体を比較する
- 実験的方法
- 進化的コンピュータシミュレーション
進化した心理学的メカニズムに関する明確に定式化された仮説と、関連する予測が特定されたら、次のステップはそれらを経験的に検証することです。進化心理学者は、自由に使える幅広い科学的方法を持っています(Schmitt, 2008; Simpson & Campbell, 2016)。後で見るように、進化心理学の科学的基盤は、単一の方法ではなく、様々な方法とデータ源からの収束した証拠に基づいています(表2.3参照)。
表2.3 進化仮説を検証するための方法とデータ源
進化仮説を検証するためのデータ源
- 考古学的記録
- 狩猟採集社会からのデータ
- 観察
- 自己報告
- 生活史データと公的記録
- 人間の産物
異なる種の比較
特定の次元に沿って異なる種を比較することは、機能的仮説を検証するための証拠の源の一つとなります。比較法は、「研究者が理解しようとしている動物の行動以外の種における形質の出現に関する予測を検証すること」を含みます(Alcock, 1993, p. 221)。例として、次の精子競争仮説を考えてみましょう:大量の精子量を生産する機能は、競合する男性の精子を置き換え、したがって女性の卵子を受精させる確率を高めることです。
この仮説を検証するための一つの戦略は、精子競争の有病率が異なる種を比較することです。高度に一夫一婦制の種では、精子競争はまれであるか、存在しません。特定の種の鳥(例えば、ハト)や哺乳類(例えば、テナガザル)では、雄と雌がつがいになって子孫を生み、つがいの外で性交渉をすることはめったにありません。他の種、例えばボノボチンパンジーでは、雌は多くの雄と交尾します(de Waal, 2006)。この種では、多くの精子競争があります。したがって、私たちは、乱交性の種では精子競争が激しく、一夫一婦制の種では低いことを知っています。
さて、テストです。精子競争が一般的である可能性の度合いによって種を並べることができます。例えば、霊長類の中では、ゴリラが最も乱交性が低く、次いでオランウータン、人間、そしてチンパンジーが最も乱交性が高いです。次に、体の大きさを補正した精巣の重量によって示される、これらの各種の精子量に関する比較データを取得できます。精子競争仮説からの予測は、多くの精子競争を示す種のオスは、より低いレベルの精子競争を示す種と比較して、より高い精巣重量(高容量の精子を示す)を持つべきであるということです。
比較証拠は、雄ゴリラの精巣が体重の0.02%、雄オランウータンが0.05%、雄人間が0.08%、そして非常に乱交的なチンパンジーが0.27%を占めることを明らかにしています(Short, 1979; Smith, 1984)。要するに、激しい精子競争を示す種の雄は、より大きな精巣容積を示し、最も精子競争が少ない種の雄は、最も低い精巣容積を示します。したがって、比較法は、精子競争仮説を支持します。
異なる種を比較する方法は、もちろん、精子競争と精巣容積に限定されません。特定の適応問題に直面することが知られている種と、その問題に直面しないことが知られている種を比較することもできます。崖に住むヤギと崖に住まないヤギを比較して、崖で草を食べるヤギが、より優れた空間定位能力など、落下を避けるための特殊な適応を持っているという仮説を検証できます。既知の捕食者を持つ種と、それらの捕食者を欠く種を比較して、それらの捕食者と戦うための特定の適応があるという仮説(例えば、捕食者の画像に遭遇したときに発せられる特定の警報音)を検証できます。要するに、異なる種を比較することは、適応機能に関する仮説を検証するための強力な方法です(Fraley, Brumbaugh, & Marks, 2005; Thornhill & Fincher, 2013)。
異文化的方法
異文化間の方法は、進化心理学的仮説を検証するための貴重なツールを提供します(Schmitt, 2008)。最も明白な方法は、普遍的であると仮定されている適応、例えば、基本的な感情(Cowen & Keltner, 2020; Ekman, 1973)、協力のための適応(Tooby & Cosmides, 2016)、または性的に分化した交配戦略(Lippa, 2009; Schmitt, 2005; Walter et al., 2020)に関するものです。異なる文化を比較することは、異なる生態系に対応すると仮定されている適応を調べるためにも使用できます。これは、人間行動生態学者が好む方法です。例えば、配偶者の好みは、寄生虫の有病率における生態学的変動に敏感であると仮定されており、これは37の文化における研究で確認されています(Gangestad, Haselton, & Buss, 2006)。
異文化間の方法は、競合する理論を互いに比較検討することによって、それらを検証するためにも使用できます。例えば、Lippa、Collaer、Peters(2010)は、53の文化にわたって心的回転課題における性差を調査しました。心的回転能力は、狩猟者が槍や他の狩猟道具が動物の軌道と一致するように空間を移動する際の軌道を予測する必要があるため、男性の狩猟適応の一部であると仮定されています。対照的に、社会的役割理論によれば、心理的な性差は、異なる文化によって割り当てられた役割の関数であり、したがって、男女間の平等が増すにつれて減少するはずです。Lippaの異文化間の研究では、2つの重要な発見がありました。(1)心的回転能力における性差は、文化を超えて普遍的であったこと、そして(2)社会的役割理論に反して、性差は、実際には男女平等が進んでいる文化の方がやや大きかったことです。要するに、異文化間の方法は、進化仮説の範囲を検証する上で非常に価値があり、また、競合する仮説を互いに比較検討する上でも価値があります。
生理学的および脳画像法
生理学的方法は、感情的覚醒、性的覚醒、ストレスなどの現象を評価するために使用できます。これらの方法は、心理的適応の生物学的基盤を特定するためと、それらの適応の設計上の特徴に関する仮説を検証するために使用できます。フリン、ウォード、ヌーン(2005)は、継親と暮らす子供は、2人の生物学的親と暮らす子供よりも高いレベルのストレスを経験するという仮説を検証しました。彼らは、実際、継子が非継子よりもコルチゾール(人々がストレスを経験したときに放出される主要なホルモンの一つ)のレベルが高いことを見出しました。別の研究では、配偶者競争に関与する主要なホルモンの一つであるテストステロンが、献身的な恋愛関係にある男性では減少するという仮説が確認されました(McIntyre et al., 2006)。さらに別の研究では、魅力的な女性の存在が男性のテストステロンレベルを上昇させることが見出されました(Ronay & von Hippel, 2010)。要するに、ホルモンアッセイなどの生理学的方法は、適応に関する仮説を検証する上でも、適応の根底にある基盤を特定する上でも価値があります。
機能的磁気共鳴画像法(fMRI)のような脳画像技術は、適応とその根底にある神経基盤に関する仮説を検証するためにますます使用されています。fMRI法は、血縁認識、言語、空間認知、恋愛感情、嫉妬に関する適応についての仮説を検証するために使用されてきました(Platek, Keenan, & Shackelford, 2007)。脳画像技術は、参加者が刺激にさらされている間、動かずにいなければならないため、調べることができる現象に現在制限がありますが(日常生活では、対照的に、人々が環境を移動する間に脳が機能します)、進化心理学的仮説を検証するためのその使用は、過去10年間で劇的に増加しています。
遺伝学的方法
伝統的な行動遺伝学的方法、例えば双生児研究や養子縁組研究は、いくつかの進化仮説を検証するために使用できます(Segal, 2011)。例えば、一つの進化仮説は、投資してくれる父親がいないで育った場合、女性が早期の性行動開始と初潮年齢に移行するという文脈依存的な適応を提案しています。これは、投資してくれる父親がいる場合の性行動開始の遅れと比較されます(例:Belsky, 1997; Ellis, 2011)。行動遺伝学的方法は、女性の性行動開始の個人差が、一つの進化仮説が示唆するように環境的に媒介されているのか、それとも代わりに遺伝的に媒介されているのかを判断することができます(Barbaro, Boutwell, Barnes, & Shackelford, 2017)。後者であれば、仮説は反証されることになります。
分子遺伝学的方法はより最近のものである。それらは、仮説上の適応の根底にある特定の遺伝子を特定するように設計されている。DRD4遺伝子の対立遺伝子の個人差はその一例である。DRD4遺伝子の7R対立遺伝子は、新規性追求と外向性と関連付けられており(Ebstein, 2006)、異なる地理的地域で劇的に異なる割合で発生する(例えば、アジアよりも北米で高い)。7R対立遺伝子は、新しい環境で資源を活用する上で有利であると仮説が立てられている(Chen, Burton, Greenberger, & Dmitrieva, 1999; Penke, Denissen, & Miller, 2007)。7R対立遺伝子が定住集団よりも遊牧集団で実質的に一般的であるという発見は、この進化心理学的仮説を支持する(Eisenberg, Campbell, Gray, & Soronson, 2008)。
分子遺伝学的研究は、人類の進化に関する興味深い発見も明らかにしました。第一に、それらは、第1章で見たように、OOAの現代人類の起源に関する競合する仮説を検証するために使用できます。第二に、それらは、乳製品の消化を促進する遺伝子など、過去1万年の間に現れたいくつかの単純な適応の遺伝的基盤を特定できます(Bersaglieri et al., 2004)。そして第三に、分子遺伝学的研究は、過去4万年、特に過去1万年(完新世)の間に、人類の適応進化が加速していることを示しています(Hawks, Wang, Cochran, Harpending, & Moyzis, 2007)。この驚くべき発見は、遺伝的進化が減速または停止したという以前の見解と矛盾しています。
男性と女性の比較
有性生殖種は通常、雄と雌の2つの形態で存在し、小さな配偶子(精子)または大きな配偶子(卵子)を生産する進化した機構によって定義されます。性の比較は、適応に関する仮説を検証するためのもう一つの方法を提供します。一つの戦略は、雄と雌が直面する異なる適応問題を分析することです。例えば、雌の体内で受精する種では、雄は「父性不確実性」という適応問題に直面します。彼らは、自分の配偶者の子孫の遺伝的な父親であるかどうかを完全に確信することはできません。しかし、雌はこの適応問題に直面しません。彼女たちは、卵子が自分の体の中からしか出てこないため、自分の卵子であって、ライバルの卵子ではないことを「知っています」。
男性は、父性の可能性を高める機能を果たす特定の適応を進化させてきました。これらの適応については第5章で詳しく検討しますが、ここで一点を明確にするために一つの例を挙げます。男性の性的嫉妬です。両性とも全体的に同じくらい嫉妬深いですが、研究では、女性よりもはるかに男性の嫉妬が、特に性的不貞の兆候によって活性化されることが示されています。これは父性不確実性の問題に対する一つの解決策を示唆しています(Buss, 2018a; Edlund & Sagarin, 2017)。男性の嫉妬は、ライバルを撃退したり、配偶者を不貞から思いとどまらせたりする行動を動機づけます。男性の嫉妬が特に性的不貞の手がかりによって引き起こされるという事実は、男性の心理学の一面、つまり親であることの不確実性という性に関連した適応問題に対応していることを示しています。要するに、一つの種内で性を比較することは、進化仮説を検証するための強力な方法となり得ます。
種内の個体の比較
もう一つの方法は、一つの種内の個体を比較することです。若い女性と年配の女性を考えてみましょう。10代の少女は、これから何年も繁殖できる可能性があります。30代後半の女性は、残りの繁殖可能な年数が少なくなっています。これらの違いを利用して、適応に関する仮説を立て、検証することができます。
例えば、もし投資してくれる男性がいなければ、若い女性は年配の女性よりも発育中の胎児を中絶する可能性が高いという仮説を立てたとします。その進化的根拠は次のとおりです。繁殖可能な年数が多く残っているため、若い女性は、より好都合な繁殖時期を待つために子供を持つ機会を失う余裕があります。年配の女性は、子供を持つ別の機会を得られないかもしれません。これら2つのグループの女性における中絶、流産、乳児殺しの割合を比較することは、この仮説を検証するための一つの方法を提供します。
種内個体の比較は、もちろん、年齢に限定されません。貧しい個体と裕福な個体を比較して、貧しい人々は資源を獲得するために「より危険な」戦略をとるという仮説を検証できます。裕福な人々は、富を守るために「より保守的」であるかもしれません。多くの強い兄弟が周りにいて守ってくれる女性と、一人っ子の女性を比較して、後者のグループの女性が身体的により脆弱で、したがって「ボディガード」として機能する友人や配偶者を選ぶ可能性が高いかどうかを見ることができます。配偶者としての魅力が異なる個体や、拡大家族の大きさが異なる個体を比較できます。要するに、種内比較は、適応に関する進化仮説を検証するための強力な方法を構成します。
異なる文脈における同じ個体の比較
もう一つのアプローチは、異なる状況における同じ個体を比較することです。例えば、東ボリビアのシリオーノ族では、特に狩猟が下手なある男性が、より上手な狩人である男性に何人かの妻を奪われました。彼は、狩猟が下手であることと、他の男性に妻を奪われたことの両方のために、グループ内での地位を失いました。人類学者のA. R. ホルムバーグは、この男性と一緒に狩猟を始め、彼に後で他の人々にその男性が殺したと言われる獲物を与え、散弾銃で獲物を殺す技術を教えました。最終的に、その男性は狩猟の成功の増加の結果として、社会的な地位が向上し、何人かの女性を性的なパートナーとして引きつけ、侮辱される側から他人を侮辱する側に変わりました(Holmberg, 1950)。
同じ個人を異なる状況で比較することは、進化した心理学的メカニズムを明らかにするための強力な方法です。2つの異なる状況で直面する適応問題、したがって、それぞれでどの心理的適応が活性化されるかについて仮説を立てることができます。狩猟能力の変化のおかげで低い地位から高い地位へと移行したシリオーノ族の男性の場合、高い地位は明らかに彼をより自信に満ちたものにしました。それはまた、男性を侮辱することからより敬意を払うことに変わった、他のシリオーノ族の男性の心理的メカニズムにも影響を与えたようです。
残念ながら、研究者が人が一つの文脈から別の文脈に移動するのを待つのは、時として困難です。人々はしばしばニッチを見つけてそこに留まります。さらに、たとえ人々が状況を変えたとしても、多くのことが一度に変化する傾向があり、研究者が変化の原因となった特定の因果要因を特定することを困難にします。特定の因果要因を分離する問題のため、科学者は時々、心理学実験で状況を「制御」しようとします。
実験的方法
実験では、通常、被験者の一つのグループが「操作」にさらされ、第二のグループが「対照」として機能します。外部の脅威が「内集団の結束」の強さに及ぼす影響に関する仮説を立てたとしましょう。その仮説は、人間が、敵対的な人間集団による侵略のような外部からの脅威に反応する機能を果たす、進化した心理的適応を持っていると述べています。脅威の状況下では、内集団への好意を示す傾向や、外集団に対する偏見の増加などによって示されるように、集団の結束が高まるはずです。
研究室では、実験者はランダムに一つの被験者グループを選び、別のグループが彼らがいる部屋に優先権を持っているため、より小さな部屋に行かなければならないかもしれないと伝えます。彼らが去る前に、実験者は研究に参加したことへの支払いとして100ドルを渡し、そのお金を2つのグループの間で好きなように分けるように指示します。対照グループも、自分たちのグループと別のグループの間でお金を分けるように指示されますが、別のグループが彼らの部屋を乗っ取るとは言われません。その後、対照グループと実験グループがどのようにしてお金を分けることに決めたかを比較することができます。もし実験グループと対照グループの間に差がなければ、私たちの予測は失敗したと結論づけます。もし脅されたグループが自分たちにより多くのお金を割り当て、対照グループが均等に割り当てた場合、私たちの予測は確認されます。つまり、外部の脅威が内集団への好意を高めるということです。要するに、実験的方法、つまり異なるグループを異なる条件にさらすことは、適応に関する仮説を検証するために使用できます。
進化的コンピュータシミュレーション
コンピュータシミュレーションを使用すると、特定の決定ルールをプログラムし、これらの戦略を使用して互いに競争する個人の適応度を測定することができます。これらのコンピュータシミュレーションは、どの戦略が進化するか、どの戦略が進化的に安定しているかを発見するために使用できます。これは、実験室や現場では非常に困難または不可能な、原則として特定の戦略が進化できるかどうかをテストするために非常に役立ちます。例として、一部の研究者は、コンピュータシミュレーションを使用して、どの配偶者選好を統合するためのアルゴリズムが、互いに競合させたときに他のアルゴリズムよりも優れているかを調査しました(Conroy-Beam & Buss, 2016)。彼らは、ユークリッドアルゴリズム、つまり実際の配偶者選択で複数の配偶者選好を最大化するアルゴリズムが、進化する可能性が最も高いことを発見しました。さらに、コンピュータシミュレーションで見つかったユークリッドアルゴリズムは、ペアで結ばれたカップルにおける実際の配偶者選択への満足度のような現実世界の結果を予測することができました。これらのトピックについては、第4章と第5章で詳しく説明します。
エージェントベースモデリングは、研究者が「エージェント」と呼ばれる個人の集団をシミュレートするコンピュータシミュレーション技術であり、エージェントはコンピュータ内で時間とともに行動し、相互作用し、進化します。エージェントベースモデルを使用すると、進化心理学者はリアルタイムで進化とその産物を観察し、その結果を人間の研究からのデータと比較することができます。プロトタイプを構築することがエンジニアが機械を設計するのに役立つのと同様に、エージェントベースモデルは心理学者が複雑な進化的または心理的プロセスを理解するのに役立ちます。エージェントベースモデルは、研究者の理論的な「盲点」を特定するのに特に役立ちます。リチャード・ファインマンが言ったように、「私が創造できないものは、私は理解していない」。もし心理学者が何らかの心理的現象がどのように機能し、進化したかを理解していると信じているならば、彼らはそのプロセスをシミュレーションで一つ一つ再創造できるはずです。そうしようとすると、研究者は見落としていた要素を発見したり、その後実験室でテストできる予期せぬ新しい現象を見たりすることがよくあります。このようにして、実験室での研究は研究者のエージェントベースモデルが説明する必要があることを伝え、その見返りに、これらのモデルの(しばしば)驚くべき行動は、研究者が次に実行する必要がある実験室での研究を伝えます。
進化仮説を検証するためのデータ源
研究方法に加えて、進化心理学者は仮説を検証するためのデータを取得できる豊富な他の情報源を持っています。このセクションでは、これらの情報源のいくつかを簡単に紹介します。
考古学的記録
世界中から確保された骨の断片は、興味深い遺物で満たされた古生物学的記録を明らかにします。炭素年代測定法により、私たちは頭蓋骨や骨格の年代を大まかに見積もり、数千年にわたる脳の大きさの進化をたどることができます。祖先のキャンプ地で見つかった大型動物の骨は、私たちの祖先が食料を確保するという適応問題をどのように解決したかを明らかにすることができます。化石化した糞便は、祖先の食生活の他の特徴に関する情報を提供することができます。骨の断片の分析は、損傷、病気、死の原因も明らかにすることができます。考古学的記録は、私たちがどのように生き、進化し、私たちの祖先が直面した適応問題の性質についての一連の手がかりを提供します。
狩猟採集社会からのデータ
現代の伝統的な人々、特に西洋文明から比較的隔離されている人々の研究も、進化仮説を検証するための豊富なデータ源を提供します。例えば、人類学者のキム・ヒルとヒラード・カプラン(1988)による研究は、成功した狩猟者が、肉がグループで共有されるため、その努力から直接利益を得ることはないが、他の生殖に関連する方法で利益を得ることを示しています。成功した狩猟者の子供たちは、グループからより多くの世話と注意を受け、その結果、健康状態が優れています。成功した狩猟者はまた、女性にとって性的に魅力的であり、より多くの愛人やより望ましい妻を持つ傾向があります。
現代の狩猟採集民からの発見は、もちろん、決定的ではありません。しかし、このデータ源は、他のデータ源と併せて、心理的適応に関する仮説を立て、検証することを可能にする証拠を提供します。
観察
体系的な観察は、進化仮説を検証するための第三の方法を提供します。人類学者のマーク・フリンは、トリニダードで体系的に観察を収集するための行動スキャン技術を考案しました(Flinn, 1988a; Flinn, Geary, & Ward, 2005)。彼は毎日、対象となる村を歩き、すべての世帯を訪れ、彼が行ったすべての観察を記録シートに記録しました。彼は、例えば、繁殖力のある妻を持つ男性は、繁殖力の低い妻を持つ男性(妊娠中または高齢の妻)よりも、より激しい「配偶者防衛」を行うという仮説を確認することができました。彼はこれを、男性が妻が繁殖力のあるときに他の男性とより多くの喧嘩をし、妻が繁殖力のないときには喧嘩が少なくなることを示す行動スキャンを通じて決定しました。観察データは、フリンのような訓練された観察者、対象となる被験者の夫や妻、友人や親戚、そしてさらには偶然の知人など、様々な情報源から収集することができます。
自己報告
被験者本人による報告は、非常に貴重なデータ源となります。自己報告データは、インタビューやアンケートを通じて確保できます。一部の心理学的現象は、自己報告を通じてのみ調べることができます。性的ファンタジーを考えてみましょう。これらは、化石を残さず、部外者によって観察できない私的な経験です。ある研究では、進化心理学者のブルース・エリスとドナルド・サイモンズが、性的ファンタジーにおける性差に関する仮説を検証しました(Ellis & Symons, 1990)。彼らは、男性の性的ファンタジーは、より多くの性的パートナーと、より多くのパートナーの入れ替わりを伴う傾向があり、より視覚的に指向していることを見出しました。女性の性的ファンタジーは、より多くのミステリー、ロマンス、感情表現、文脈を持つ傾向がありました。自己報告がなければ、この種の研究は実施できませんでした。
自己報告は、配偶者の好み(Buss, 1989a; Walter et al., 2020)、配偶者に対する暴力(Kaighobadi & Shackelford, 2009)、欺瞞の戦術(Tooke & Camire, 1991)、社会的階層で出世するための戦術(Buss et al., 2020; Kyl-Heku & Buss, 1996)、そして協力と援助のパターン(McGuire, 1994)に関する様々な進化心理学的仮説を検証するために使用されてきました。
すべてのデータ源と同様に、自己報告にも偏りと限界があります。人々は、婚外恋愛や異常な性的ファンタジーなど、望ましくないと判断されることを恐れて、行動や思考を明かすことをためらうかもしれません。人々は、実験者を喜ばせるため、あるいは研究を妨害するために、嘘をついたり、物事を言ったりするかもしれません。これらの理由から、進化心理学者は、自己報告にのみ頼らないようにしています。
生活史データと公的記録
人々は公文書にその人生の痕跡を残します。結婚と離婚、出生と死亡、犯罪と軽犯罪はすべて公的記録の一部です。ある一連の研究では、進化生物学者のボビー・ローは、数世紀前に記録されたスウェーデンの異なる教区からの結婚、離婚、再婚に関するデータを掘り起こすことができました。これらの教区の司祭は、これらの公的出来事について、細心の注意を払って正確かつ詳細な記録を保持していました。400年前の結婚率と離婚率を見ることによって、今日起こっているパターンが人間の歴史の中で長く続いてきたものなのか、それとも単に現代の産物なのかを見ることができます。ローは、これらの公的記録を用いて、多くの進化仮説を検証することができました。彼女は、例えば、裕福な男性は貧しい男性と比較して、より若く、したがってより繁殖力のある女性と結婚する傾向があることを確認しました(Low, 1991)。
公的記録は、要するに、特に他のデータ源と組み合わせて使用される場合、創造的な科学者が進化心理学的仮説を検証するための宝の山となり得る。
人間の製品
人間が作るものは、その進化した心の産物です。例えば、現代のファストフードレストランは、進化した味覚の好みの産物です。ハンバーガー、フライドポテト、ミルクシェイク、ピザは、脂肪、砂糖、塩、タンパク質で満たされています。それらがよく売れるのは、まさにこれらの物質に対する進化した欲求に対応し、それを悪用しているからです。したがって、食品の創造は、進化した味覚の好みを明らかにします。
他の種類の人間の産物は、私たちの進化した心の設計を明らかにします。例えば、ポルノグラフィーやロマンス小説の産業は、共通のファンタジーの創造物と見なすことができます。演劇、絵画、映画、音楽、オペラ、小説、ソープオペラ、そしてポピュラーソングに共通するテーマはすべて、私たちの進化した心理学について何かを明らかにします(Carroll, 2016)。したがって、人間の創造物は、進化仮説を検証するための追加のデータ源として役立ちます。
単一のデータソースの限界を超える
すべてのデータソースには限界があります。化石記録は断片的で、大きなギャップがあります。現代の狩猟採集民については、現在の慣行が現代の影響によってどの程度汚染されているかわかりません。自己報告では、人々は嘘をついたり、真実を知らなかったりする可能性があります。観察報告では、行動の多くの重要な領域は詮索好きな目から隠されています。そうでないものも、観察者の偏見によって歪められる可能性があります。実験室での実験は、しばしば作り物で人工的であり、現実世界への一般化可能性を損ないます。公的記録からの生活データも、客観的に見えるかもしれませんが、体系的な偏見の対象となる可能性があります。人間の産物でさえ、有効であるかもしれないし、そうでないかもしれない一連の推論を通じて解釈されなければなりません。
これらの問題に対する解決策は、進化仮説を検証する際に複数のデータソースを使用することです。同じ方法論的な限界を共有しないデータソース全体で一貫して現れる発見は、特に強力です。複数のデータソースを使用することによって、研究者は単一のデータソースの限界を超え、進化心理学のためのより確固たる経験的基盤に到達することができます。
適応問題を特定する
人間は、多くの種と同様に、人類の進化の歴史を通じて多くの適応問題に直面し、多くの複雑な適応メカニズムを生み出してきたことは明らかです。次の重要な問いは、これらの適応問題が何であるかをどのようにして知るかということです。
概念的な作業だけでは、人間が直面してきたすべての適応問題の完全なリストを決定的に得ることはできません。この不確定性は、いくつかの要因によって引き起こされます。第一に、私たちは進化の時計を巻き戻して、私たちの祖先が過去に直面したすべてのことを見ることはできません。第二に、各新しい適応は、他の適応メカニズムと協調するようになるなど、新しい適応問題を生み出します。人間の適応問題の完全なセットを特定することは、科学者が今後何十年も取り組むことになるであろう巨大なタスクです。それにもかかわらず、いくつかのガイドラインが私たちにスタートを与えてくれます。
現代進化論からの指針
現代進化論の構造そのものが、一つの指針の源となります。それは、設計上の違いをコードする遺伝子の示差的繁殖が、子孫を生み出すことを通じても、遺伝的親族が子孫を生み出すのを助けることを通じても、適応が形成される進化プロセスのエンジンであることを教えてくれます。したがって、すべての適応問題は、定義上、繁殖に必要なもの、あるいは間接的に繁殖を助けるものでなければなりません。
したがって、まず、進化論は私たちを次の広範な適応問題のクラスに導きます。
- 生存と成長の問題:生物を繁殖可能な時点まで到達させること。
- 交配の問題:配偶者を選び、惹きつけ、維持し、成功した繁殖に必要な性的行動を行うこと。
- 育児の問題:子孫が生き残り、繁殖可能な時点まで成長するのを助けること。
- 遺伝的親族を助ける問題:自分の遺伝子のコピーを持つ非子孫の親族の繁殖を助けることに伴う課題。
これらの4つの問題のクラスは、優れた出発点を提供します。
普遍的な人間構造の知識からのガイダンス
第二の指針の源は、普遍的な人間構造に関する蓄積された知識から得られます。時折の隠者を除いて、すべての人間は集団で生活しています。この事実を知ることは、人間が解決策を進化させた可能性のある、多くの潜在的な適応問題を示唆します。例えば、一つの明白な問題は、どのようにして自分が集団に含まれ、排斥されたり追い出されたりしないようにするかということです(Baumeister & Leary, 1995; Kurzban & Neuberg, 2005)。もう一つの問題は、集団生活は、同じ種のメンバーがより近くに住み、したがって、生存と繁殖に必要な資源へのアクセスをめぐって、互いにより直接的な競争関係にあることを意味するということです。
既知のすべての人間集団には社会的階層があります。地位の階層が普遍的であるという事実は、別のクラスの適応問題を示唆しています(第12章参照)。これらには、出世する問題(階層を上るにつれて資源へのアクセスが増加するため)、地位の低下を防ぐ問題、あなたの地位を狙う今後の競争相手に対処する問題、そして従属的な立場にあることの課題が含まれます。要するに、集団生活や地位の階層のような人間社会の相互作用の普遍的な特徴を特定することは、人間の適応問題を特定するための指針を提供します。
伝統的な社会からのガイダンス
第三の指針は、狩猟採集民のような伝統的な社会から得られます。これらの社会は、私たちが進化した条件を、現代社会よりも密接に反映しています。例えば、人間が人類の歴史の99パーセント、つまり1万年前の農業の出現前の過去数百万年間、狩猟採集民であったという強力な証拠があります(Kraft et al., 2023; Tooby & DeVore, 1987)。したがって、狩猟採集社会を調べることは、私たちの祖先が直面した適応問題の種類に関する手がかりを提供します。
少なくとも、銃や他の武器が発明される前に利用可能だった道具では、単独で大型の獲物を狩ることは事実上不可能です。狩猟採集社会では、大型の獲物の狩猟は、ほとんどの場合、集団または連合で行われます。成功するためには、これらの連合は、作業の分担方法や集団の努力の調整方法など、一連の適応問題を解決しなければならず、その両方には明確なコミュニケーションが必要です。伝統的に、大型の獲物の狩猟は主に男性の活動と見なされてきましたが、最近の証拠は、女性がこれまで考えられていたよりも狩猟においてより積極的な役割を果たしていた可能性を示唆しています(Anderson, Chilczuk, Nelson, Ruther, & Wall-Scheffler, 2023)。このトピックについては、第3章で考察します。
古考古学と古人類学からのガイダンス
第四の指針は、石と骨から得られます。例えば、私たちの人類の祖先の歯の分析は、祖先の食生活の性質に関する情報を明らかにします。骨格の骨折の分析は、私たちの祖先がどのように死んだかに関する情報を明らかにします。骨は、祖先の人類集団を悩ませた病気の種類に関する手がかりさえも与え、それによって別の適応問題のセットを明らかにすることができます。
現在のメカニズムからのガイダンス
第五の、そして非常に有益な情報源は、現在の人間の心理的メカニズムから得られます。文化を超えて最も一般的な人間の恐怖症が、ヘビ、クモ、高所、暗闇、見知らぬ男性であり、例えば、車、バー、酔っ払いではないという事実は、祖先の生存問題に関する豊富な情報を明らかにします。それは、私たちが祖先の危険の可能性を恐れる傾向を進化させたが、現代の危険は恐れないことを教えてくれます。性的嫉妬の普遍性は、祖先の女性と男性が常に配偶者に性的に忠実ではなかったことを教えてくれます。要するに、私たちの現在の心理的メカニズムは、私たちの祖先を悩ませた適応問題の性質を見るための窓を提供します。
タスク分析からのガイダンス
適応問題(およびサブ問題)を特定するためのより正式な手順は、タスク分析として知られています(Marr, 1982)。タスク分析は、人間の構造に関する観察(例:人間は地位の階層を持つ集団で生活する)または十分に文書化された現象(例:人間は遺伝的親族を優遇する)から始まります。タスク分析は、この質問を提起します:この構造または現象が発生するためには、どの情報処理手順と行動タスクを実行する必要がありますか?
人々が非親族よりも遺伝的親族を助ける傾向があるという観察を考えてみましょう。もしあなたが大学生なら、両親が授業料、部屋代、食事代、衣服、または交通手段のいずれかの方法であなたを助けている可能性が高いです。また、両親が隣人の子供たちを助けていない可能性も高いです。たとえ彼らが彼らをとても気に入っていてもです。人々はまた、遠い遺伝的親族よりも近い遺伝的親族を助ける傾向があります(Stewart-Williams, 2008)。
タスク分析は、祖先の環境で利用可能であった情報のみを使用して、それが発生するために実行されなければならない認知手順を特定することを含みます。例えば、人々は、自分の遺伝子のコピーを持っている人々、つまり親族認識の問題を特定する方法を必要とします。彼らは、身体的外見の特徴や成長期における近接性など、当時利用可能であった情報のみを使用してこの問題を解決しなければなりませんでした。さらに、人々は、遺伝的親族がどれだけ近い関係にあるかを測る問題、つまり親族関係の近さの問題を解決する必要があります。人々は、ほとんどの場合、これらのことについて意識的に考えていません。それらは自動的に起こります。要するに、タスク分析は、私たちが観察する現象が発生するために解決されなければならない適応問題、およびそれらを解決できる潜在的な適応の設計上の特徴を特定することを可能にします。
適応問題の構成
本書は、人間の適応問題と、それらを解決するために進化した心理的解決策を中心に構成されています。私たちは、少なくとも思春期まで生存しなければ繁殖はあり得ないため、生存問題から始めます。次に、望ましい配偶者を選び、惹きつけ、維持するという問題を含む、交配の問題に進みます。その後、交配の産物である子供に移ります。人間の子供は、助けなしには生き残り、成長することができません。したがって、このセクションでは、親が子供に投資する方法を扱います。これらすべては、より大きな血縁集団、つまり人間が遺伝的親族と共有するDNAの鎖の中で起こります。その後、本書は、私たちが生きているより大きな社会領域、つまり協力、攻撃、男女間の対立、社会的地位に移ります。最終章は、より広い焦点に戻ります。それは、進化心理学の理論的傘の下で心理学の主要な分野を再定式化することを扱い、文化の進化と、文化進化を説明する可能性のあるさまざまな理論を探求します。
まとめ
本章では、4つのトピックを取り上げました:(1)私たちの進化した心理学的メカニズムに関する仮説を生成する論理、(2)進化のプロセスの産物、(3)進化した心理学的メカニズムの性質、および(4)これらの仮説を検証するための科学的手順。
進化仮説の論理は、最も一般的なものから最も具体的なものへと進む4つの分析レベルの検討から始まります。つまり、一般進化論、中間レベルの進化論、特定の進化仮説、そしてこれらの仮説から導き出された経験的現象に関する特定の予測です。仮説生成の一つの方法は、より高いレベルから始めて下に移動することです。中間レベルの理論はいくつかの仮説を生み出すことができ、それぞれが今度はいくつかの検証可能な予測を生み出します。これは、仮説と予測形成の「トップダウン」戦略です。
第二の方法は、男性が女性の外見に重要性を置くなど、存在することが知られているか観察されている現象から始めることです。この現象から、それが設計された可能性のある機能に関する仮説を生成することができます。このボトムアップ法は、リバースエンジニアリングと呼ばれることもあり、トップダウン法を補完するのに役立ちます。
進化の過程は、適応、適応の副産物、そしてランダムな効果またはノイズという3つの産物を生み出します。進化心理学者は、適応に焦点を当てる傾向があります。より具体的には、彼らは人間性を構成する適応の特別なサブクラスの一つである心理的メカニズムに焦点を当てます。
心理学的メカニズムは、人類の進化の歴史を通じて再発的に生存または繁殖の特定の問題を解決してきたため、その形で存在する情報処理装置です。それらは、情報の狭いサブセットのみを取り込み、決定規則を通じてその情報を変換し、生理学的活動、他の心理学的メカニズムへの情報、または顕在的な行動の形で出力を生成するように設計されています。進化した心理学的メカニズムの出力は、特定の適応問題の解決に向けられています。進化した心理学的メカニズムは、「心をその関節で切り分ける」ための非恣意的な基準を提供し、問題特異的である傾向があり、数が多く、本質的に機能的です。
進化した心理学的メカニズムに関する仮説が定式化されたら、科学的な試みの次のステップはそれを検証することです。進化仮説の検証は、比較に依存しており、特定の点で異なると予測されたグループが実際にそうであるかどうかを明らかにします。この方法は、異なる種を比較したり、異なる文化の人々を比較したり、人々の生理学的反応や脳の画像を比較したり、異なる遺伝子を持つ人々を比較したり、種内の雄と雌を比較したり、各種の異なる個体を比較したり、異なる文脈で同じ個体を比較したりすることによって、仮説を検証するために使用できます。コンピュータシミュレーションはまた、どの戦略が進化できるか、どの戦略が他の戦略を打ち負かす傾向があるかを評価するためにも使用されます。
進化心理学には、考古学的記録、現代の狩猟採集社会、自己報告、観察者報告、実験室での被験者から引き出されたデータ、公的記録からの生活史データ、そして人々によって作られた製品を含む、活用できる追加の情報源が豊富にあります。
すべてのデータ源には長所がありますが、それぞれに限界もあります。それぞれは、通常、他のデータ源からは同じ形で得られない情報を提供します。そして、それぞれには、他のものと共有しない欠点と弱点があります。2つ以上のデータ源を使用して進化仮説を検証する研究は、単一の情報源に依存する研究よりも優れています。
この章の最後のセクションでは、適応問題の主要なクラスを概説します。現代進化論からは、4つのクラスの適応問題が導き出されます。生存と成長の問題、交配の問題、育児の問題、そして遺伝的親族との相互作用の問題です。適応問題を特定するための追加の洞察は、普遍的な人間構造、伝統的な部族社会、古考古学、タスク分析、そして現在の心理的メカニズムの知識から得られます。高所恐怖症、脂肪の多い食物への嗜好、サバンナのような風景への好みなどの現在のメカニズムは、過去の適応問題の性質を見るための窓を提供します。
クリティカルシンキングの質問
- 嫉妬は、その活性化のために入力(例:配偶者密猟者の存在)と、その機能を達成するための行動という形での出力(例:貴重な関係への脅威を撃退する)の両方を必要とします。この例が、「進化した心理学的メカニズム」の主要な構成要素をどのように示しているか説明してください。
- 進化仮説を検証する方法には、異文化間の方法、実験的方法、生理学的方法が含まれます。進化心理学的仮説を検証するために、なぜ単一の方法よりも複数の方法の方が優れているのか説明してください。
- 適応問題は、狩猟や採集によって食物を得たり、捕食者を撃退したり、子供を守ったりするなど、より伝統的な文化の人々が達成しなければならないタスクを観察することによって、時々特定することができます。この方法が、人間が歴史的に直面した他の適応問題を特定するためにどのように使用できるか説明してください。
推薦図書
Al-Shawaf, L. (2019, August 20). Seven key misconceptions about evolutionary psychology. Areo. Retrieved from https:// areomagazine.com/2019/08/20/seven-key-misconceptions-about-evolutionary-psychology/
Buss, D. M. (Ed.). (2016a). The handbook of evolutionary psychology (2nd ed.). Hoboken, NJ: Wiley.
Haselton, M. (2018). Hormonal: The hidden intelligence of hormones-How they drive desire, shape relationships, influence our choices, and make us wiser. New York: Little, Brown.
Lewis, D. M., Al-Shawaf, L., Conroy-Beam, D., Asao, K., & Buss, D. M. (2017). Evolutionary psychology: A how-to guide. Ameri- can Psychologist, 72 (4), 353-373.
Nettle, D., & Scott-Phillips, T. (2023). Is a non-evolutionary psychology possible ?. In Evolutionary thinking across disciplines: Problems and perspectives in generalized Darwinism (pp. 21-42). Cham: Springer International Publishing.
Pinker, S. (1997). How the mind works. New York: Norton.
Rodeheffer, C. D., Daugherty, J. R., & Brase, G. L. (2011). Resistance to evolutionary psychology as a continuation of conflicts over scientific integration. Futures, 43, 777-786.
Thornhill, R., & Fincher, C. L. (2013). The comparative method in cross-cultural and cross-species research. Evolutionary Biology, 40, 480-493.
Tooby, J., & Cosmides, L. (2016). Theoretical foundations of evolutionary psychology. In D. M. Buss (Ed.), The handbook of evolutionary psychology (2nd ed., pp. 3-87). Hoboken, NJ: Wiley.
Winegard, B. M., Winegard, B. M., & Deaner, R. O. (2014). Misrepresentations of evolutionary psychology in sex and gender textbooks. Evolutionary Psychology, 12 (3), 474-508.
