
ご提示いただいたファイル「fnhum-08-00419.pdf」(論文タイトル:Response inhibition and interference control in obsessive–compulsive spectrum disorders)の全文を、省略なく正確に日本語に翻訳いたします。元の論文の構造、見出し、表・図の説明、参考文献フォーマットをすべて維持し、学術的なニュアンスを損なわないように努めました。
反応抑制および干渉制御における強迫性スペクトラム障害
Laura S. van Velzen¹,²†, Chris Vriend¹,²,³,⁴†, Stella J. de Wit¹,²,³, Odile A. vanden Heuvel²,³,⁴*
¹GGZ InGeest, アムステルダム, オランダ
²Neuroscience Campus Amsterdam (NCA), アムステルダム, オランダ
³VU University Medical Center 精神医学部, アムステルダム, オランダ
⁴VU University Medical Center 解剖学・神経科学部, アムステルダム, オランダ
Edited by: Guido Van Wingen, Academic Medical Center Amsterdam, オランダ
Reviewed by: Robert Hester, University of Melbourne, オーストラリア; Jens Kuhn, University of Cologne, ドイツ
*Correspondence: Odile A. vanden Heuvel, Department of Psychiatry, VU University Medical Center, PO Box 7057, Amsterdam 1007 MB, Netherlands; e-mail: oa.vandenheuvel@vumc.nl
†Laura S. van VelzenおよびChris Vriendは本研究に等しく貢献しました。
過去20年にわたり、運動反応抑制および干渉制御は、行動および神経精神障害の発達における重要な役割により、相当な科学的努力と注意を払われてきました。神経イメージング研究の結果から、運動反応抑制および干渉制御は皮質-線条体-視床-皮質(CSTC)回路に依存していることが示されています。強迫性障害(OCD)や関連障害(注意欠如・多動性障害(ADHD)、トゥレット症候群(TS)、抜毛症(TTM)など)を含む多くの神経精神障害において、CSTC回路内の構造的および機能的異常が報告されています。これらの障害は運動反応抑制および干渉制御の障害を共有しており、これがそれらの行動的および認知的症状の一部の基礎となっている可能性があります。これらの障害における抑制機能に関するタスク関連神経イメージング研究の結果は、障害されたタスクパフォーマンスがCSTC回路の異常な動員と関連していることを示しています。以前の研究では、抑制パフォーマンスはドーパミン、ノルアドレナリン、セロトニンシグナルに依存しており、これらの神経伝達物質はこれらの障害の病態生理学に関与していることが示されています。
本ナラティブレビューでは、OCDおよび関連障害における抑制関連機能不全の共通および障害特異的な病態生理メカニズムについて議論します。
キーワード: 反応抑制、強迫性障害、トゥレット症候群、抜毛症、注意欠如・多動性障害、干渉制御
はじめに
反応抑制とは、適切でなくなったり必要でなくなったりした先行的行動を抑制する能力であり、日常生活における目標指向的行動にとって重要です(Chambers et al., 2009)。過去数十年にわたり、研究者は反応抑制への関心を高めてきました。反応抑制は、衝動性および強迫性の一定の側面の操作化と考えられています(Bari & Robbins, 2013)。衝動性は一般的に、衝動に従って行動する傾向、すなわち自発的なコントロールなしに即座に行われる行動と定義されます。一方、強迫性は特定の行動を繰り返す傾向およびその行動がもはや適切でない場合でもそれを抑制できない傾向と定義されます(Bari & Robbins, 2013)。日常生活における反応抑制の重要性から、多くの神経心理学的パラダイムが抑制パフォーマンスを探るために開発されてきました。これらのパラダイムでは、被験者に目標刺激に応答するよう求められますが、無関係または気を散らす刺激、または気を散らす刺激の特徴については応答を保留するよう指示されます(Nigg, 2000)。反応抑制は単一の構成要素ではなく、運動反応抑制および干渉制御から構成されます。運動反応抑制は、先行的かつ自動的な運動反応の抑制を含み、さらに行動抑制(または行動抑制)および行動取消(Schachter et al., 2007)に細分化することができます。Go/No-Goタスク(Donders, 1969)は行動抑制を探るために考慮され、Stop-signalタスク(Logan, 1994)は行動取消を測定するために考慮されます(これらのタスクの説明については図1を参照)。一方、干渉制御は、関連および無関係な刺激または刺激の特徴の競合による干渉を防ぐために必要な認知制御を指します(Nigg, 2000)。ストループタスク、フランカータスク、およびシモンタスクを含むいくつかのタスクは、干渉制御の尺度として機能します(図1を参照)。抑制負荷が最高となるのはStop-signalタスクであることが提案されています。これは、抑制する必要のある反応がすでに開始されているためです(Schachter et al., 2007)。運動反応抑制タスクとは対照的に、干渉制御タスクは応答選択プロセスにも依存する可能性があります(Nee et al., 2007)。干渉制御、行動抑制、および行動取消は、それぞれ反応抑制の早期、中間、および後期のプロセスを表していることが示唆されています(Sebastian et al., 2013a)。強迫性スペクトラム障害における症状は、手を洗う、髪を引っ張る、運動チック、または衝動的行動などの特定の行動を抑制できないことによって特徴づけられます。したがって、反応抑制はこれらの共有症状の神経底盤を調査するための適切な尺度である可能性があります。
本ナラティブレビューでは、健康な被験者およびこれらの障害を有する患者における反応抑制障害の神経解剖学的および機能的基盤を調査した研究の概要を提供します。ただし、このレビューは決して現在の文献を網羅的に説明するものではありません。我々は、これらの障害における抑制機能不全および症状の基礎となる可能性のある共有メカニズムに焦点を当てます。
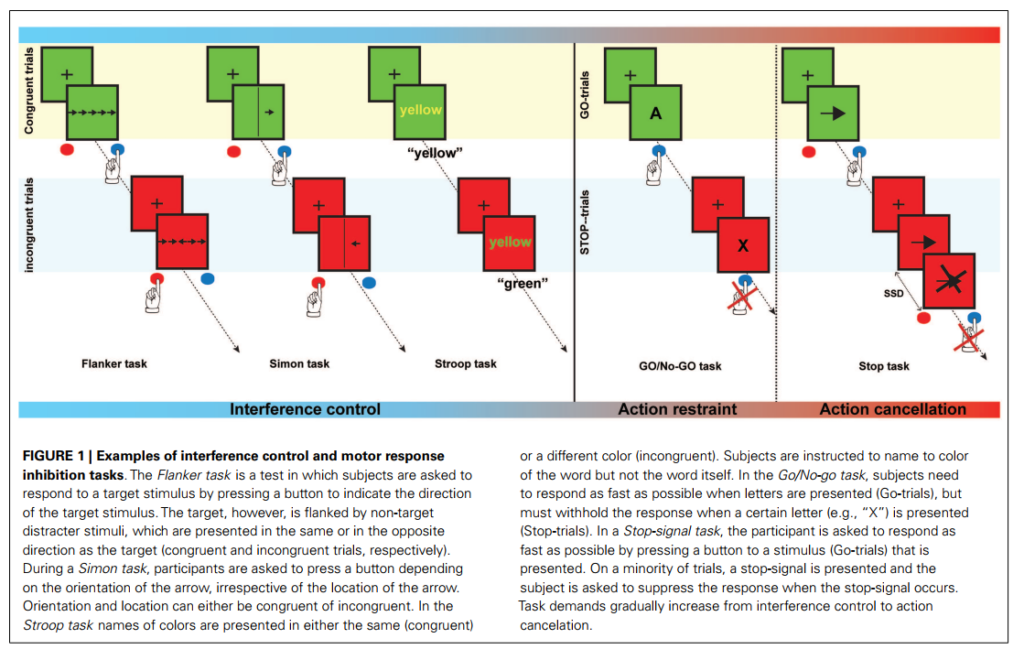
[Image blocked: 図1: 干渉制御および運動反応抑制タスクの例。フランカータスクでは、被験者は目標刺激の方向を示すボタンを押すよう求められます。ただし、目標刺激は非目標の気を散らす刺激に挟まれており、これらは目標刺激と同じ方向または反対方向に提示されます(それぞれ合同試行および非合同試行)。シモンタスクでは、参加者は矢印の向きに応じてボタンを押すよう求められますが、矢印の位置については無視します。向きと位置はどちらも合同または非合同になる可能性があります。ストループタスクでは、色の名前が同じ色(合同)または異なる色(非合同)で提示されます。被験者は、単語そのものではなく、単語の色を名前として言うよう指示されます。Go/No-Goタスクでは、文字が提示されたとき(Go試行)には可能な限り速やかに応答しなければなりませんが、特定の文字(例えば、「X」)が提示されたとき(Stop試行)には応答を保留しなければなりません。Stop-signalタスクでは、参加者は提示された刺激にボタンを押して可能な限り速やかに応答するよう求められます(Go試行)。少数の試行ではストップシグナルが提示され、参加者はストップシグナルが発生したときに応答を保留するよう求められます。タスクの要求は、干渉制御から行動取消へと段階的に増加します。]
図1: 干渉制御および運動反応抑制タスクの例。フランカータスクでは、被験者は目標刺激の方向を示すボタンを押すよう求められます。ただし、目標刺激は非目標の気を散らす刺激に挟まれており、これらは目標刺激と同じ方向または反対方向に提示されます(それぞれ合同試行および非合同試行)。シモンタスクでは、参加者は矢印の向きに応じてボタンを押すよう求められますが、矢印の位置については無視します。向きと位置はどちらも合同または非合同になる可能性があります。ストループタスクでは、色の名前が同じ色(合同)または異なる色(非合同)で提示されます。被験者は、単語そのものではなく、単語の色を名前として言うよう指示されます。Go/No-Goタスクでは、文字が提示されたとき(Go試行)には可能な限り速やかに応答しなければなりませんが、特定の文字(例えば、「X」)が提示されたとき(Stop試行)には応答を保留しなければなりません。Stop-signalタスクでは、参加者は提示された刺激にボタンを押して可能な限り速やかに応答するよう求められます(Go試行)。少数の試行ではストップシグナルが提示され、参加者はストップシグナルが発生したときに応答を保留するよう求められます。タスクの要求は、干渉制御から行動取消へと段階的に増加します。
健康な対照群における反応抑制の神経相関 神経イメージングによる反応抑制
健康な対照群を対象とした神経イメージング研究により、反応抑制の神経底盤が明らかになっています(優れたレビューについては、Robbins (2007)、Chambers et al. (2009)、および Aron (2011) を参照)。反応抑制への理解への主要な貢献は電気生理学的研究から得られていますが、本レビューでは神経イメージング研究に焦点を当てます。反応抑制の電気生理学に関心のある読者については、Huster et al. (2013) を参照してください。要約すると、反応抑制は主に右側側頭前頭領域のネットワークを活性化させます。下前頭回(IFG)および前補足運動野(pre-SMA)は重要な構成要素です(Aron et al., 2003b; Chambers et al., 2006; Floden & Stuss, 2006; Cai et al., 2012)。さらに、ニューラルストップシグナルは、これらの前頭領域から皮質-線条体-視床-皮質(CSTC)投射を介して運動皮質に送信されます(Chambers et al., 2009)。反応抑制の構成要素は、重複しているが異なる脳領域に依存していることが見出されています。干渉抑制、行動抑制、および行動取消は、すべてIFG、前島、前帯状回(ACC)、背外側前頭前皮質(DLPFC)、pre-SMA、および頭頂葉領域の活性化に関連しています(Wager et al., 2005; Nee et al., 2007; Sebastian et al., 2013b)。抑制タスクの負荷が増加すると、前頭-線条体領域の活性化が増加し、追加の抑制関連脳領域が動員されます(Blasi et al., 2006; Swick et al., 2011; Sebastian et al., 2013b)。ただし、各タスクはそれらが表す固有の認知プロセスに依存して異なる脳領域も動員します。応答選択に関与する領域(例えば、頭頂葉皮質)は、干渉制御タスクおよび行動抑制タスクにおいて、より大きな範囲で活性化されます(Rubia et al., 2001; Sebastian et al., 2013b)。
反応抑制における神経伝達物質
神経活性化の違いに加えて、干渉制御、行動抑制、および行動取消を基盤とする神経伝達物質系の違いも観察されてきました。現在の研究は、干渉制御がセロトニンおよびドーパミン神経伝達に依存していることを示唆しています。セロトニンおよびドーパミンの枯渇は、ストループタスクにおける非合同試行での干渉効果を減少させ、それによってパフォーマンスを改善することが示されています(Schmitt et al., 2000; Scholes et al., 2007)。セロトニンレベルの低下は、覚醒と注意を増加させることによってパフォーマンスを改善する可能性があります(Scholes et al., 2007)。セロトニン枯渇後のストループタスクのパフォーマンスでは、DLPFCおよびACCの活性化増加が観察されました(Horacek et al., 2005)。ストループタスクのパフォーマンスは、右DLPFCにおけるセロトニントランスポーター(SERT)結合と正の相関があることが示されています(Madsen et al., 2011)。ただし、相反する結果も報告されています。神経イメージング研究では、女性のストリオイドにおけるドーパミントランスポーター(DaT)結合の減少およびストリオイドにおける投稿シナプス性D2受容体可用性の減少が、ストループタスクでのパフォーマンスの低下と関連していることが示されています(Mozley et al., 2001; Volkow et al., 1998)。ドーパミンD2受容体作動薬の投与は干渉を減少させ、それによってストループタスクでのパフォーマンスを改善しました(Roesch-Ely et al., 2005)。健康な被験者に関する利用可能な文献に基づくと、セロトニンおよびドーパミンは両方とも干渉制御において重要であるように見えます。行動抑制(Go/No-Goタスク)は、主にセロトニンによって媒介されているようです(証拠のレビューについては、Eagle et al. (2008) を参照)。セロトニン枯渇は、Go/No-Goタスクにおける抑制時のIFG活性化の減少および誤り監視時の内側前頭前葉の活性化の減少を示しています(Rubia et al., 2005a; Evers et al., 2006)。セロトニン2C受容体作動薬(Anderson et al., 2002)またはミルタザピン(Vollmetal., 2006)(これらはノルアドレナリンおよびセロトニン系の両方に作用する)の投与は、右IFGにおける抑制関連活性化の増加をもたらしました。ただし、ドーパミンはメチルフェニデート(ドーパミン再取り込み阻害薬)がGo/No-Goタスクでのパフォーマンスを改善し、タスク関連のストリオイド活性化を減少させたことから、行動抑制にも関与している可能性があります(Vaidya et al., 1998)。ドーパミンによる行動取消への重要な役割を支持するいくつかの証拠線があります。ストリオイドにおけるドーパミンD2/3受容体可用性の増加は、ストップシグナルタスクでのパフォーマンスの向上(すなわち、ストップシグナル反応時間(SSRT)の短縮)および背側尾状核および被殻における抑制関連活性化と正の相関があることが関連しています(Ghahremani et al., 2012)。また、D2受容体作動薬の投与は行動取消を改善しました(Nandam et al., 2013)。メチルフェニデートおよびアトモキセチン(これらはドーパミンおよびノルアドレナリン系の両方を標的とする)の投与は、シナプス間隙からの再取り込みを阻害することにより、ヒトおよび動物においてSSRTを減少させました。これにより、ノルアドレナリンも運動反応抑制に関与している可能性が示唆されます(Chamberlain et al., 2006b; Eagle et al., 2007; Bari et al., 2009; Nandam et al., 2011)。セロトニンは、選択的セロトニン再取り込み阻害薬(SSRIs)およびセロトニン枯渇を使用しても、ヒトまたは動物におけるストップシグナルタスクでの行動取消に影響を与えないため、ストップシグナルタスクのパフォーマンスを媒介していないように見えます(Clark et al., 2005; Chamberlain et al., 2006b; Bari et al., 2009; Eagle et al., 2009; Drueke et al., 2010)。まとめると、干渉制御はセロトニンおよびドーパミンの両方によって媒介されているようです。行動抑制は主にセロトニンによって媒介されているようです。一方、行動取消はドーパミンおよびノルアドレナリンによって媒介されているようです。反応抑制における神経薬理学の詳細情報については、Dalley & Roiser (2012) および Bari & Robbins (2013) を参照してください。
反応抑制における遺伝子
いくつかの遺伝子関連研究では、ドーパミン作動性およびセロトニン作動性システムに関与する遺伝子と反応抑制との関係が調査されてきました。ドーパミンD4受容体遺伝子(DRD4)の遺伝子多型(機能活動の減少と関連しています;Asghari et al., 1995)は、ストップシグナルタスクでのパフォーマンスの低下と関連していることが見出されています(Congdon et al., 2008)。ただし、相反する結果も報告されています(Kramer et al., 2009)。DRD2遺伝子はドーパミン受容体D2をコードします。ストリオイドにおけるD2受容体可用性の減少と関連しているTaqIAアレルの存在(Thompson et al., 1997)は、ストップシグナルタスクでの反応抑制の低下と関連しています(White et al., 2008)。また、皮質および視床におけるD2受容体可用性の減少と関連している第二の多型(Hirvonen et al., 2009)は、行動取消の不良と関連していることが見出されています(Colzato et al., 2010)。DaT発現の増加と関連している遺伝子多型は、干渉制御タスクでのパフォーマンスの低下および動作取消時のSTNおよびpre-SMAにおける脳活性化の減少と関連していることが関連しています(Cornish et al., 2005; Congdon et al., 2009)。さらに、DaT遺伝子における2つの新規単一ヌクレオチド多型は、個々のSSRTを予測し(Cummins et al., 2012)、これらの多型のいずれかの遺伝子型は、タスクパフォーマンス時の前頭領域および尾状核核の活性化を予測します。カテコール-O-メチル移 transferase(COMT)およびモノアミン酸化酸A(MAO-A)遺伝子の遺伝子多型(神経伝達物質代謝において役割を果たす酵素をコードする)は、抑制関連活動における正常な変異と関連していることが関連しています。COMTはドーパミンおよびノルアドレナリンの分解に関与し、MAO-Aはドーパミン、ノルアドレナリン、およびセロトニンの分解に関与します。機能が低下したCOMTの多型(Chen et al., 2004)は、行動取消時のIFG活性化の増加および干渉抑制の減少と関連していることが関連しています(Congdon et al., 2009;ただし、相反する結果も報告されています;Kramer et al., 2007)。MAO-A遺伝子の多型(MAO-A活性の増加と関連している)は、右IFGおよびACCにおける活性の増加および上部頭頂葉皮質における活性の減少と関連していることが関連しています(Passamonti et al., 2006)。セロトニン伝達に関与する遺伝子にも焦点が当てられた遺伝子関連研究があります。脳内でのセロトニン合成は、トリプタン-ヒドロキシラーゼ-2(TPH-2)によって調節されます(Walther & Bader, 2003)。TPH-2遺伝子の多型のTアレルに対してホモジゴウスである個人は、ストップシグナルタスクでのSSRTの増加を示しています(Stoltenberg et al., 2006)。また、別の研究では、TPH-2遺伝子の他の2つの多型が、EEG研究における行動抑制時の脳活性化の減少と関連していることが見出されています(Baehne et al., 2009)。最後に、Osinsky et al. (2009)は、TPH-2遺伝子のプロモーター領域に位置する多型が、ストループタスクのパフォーマンスにおける反応時間に影響を与えることを発見しました。これらの発見の解釈は、これらの多型がセロトニンレベルにどのように影響するかが不確実であるため、困難を伴います。
SERT遺伝子(SLC6A4)の多型(シナプス間隙からの再取り込み速度を減少させる)は、干渉抑制の減少と関連していることが関連しています(Holmes et al., 2010)。ただし、行動取消とは関連していないことが関連しています(Clark et al., 2005)。SERT機能が低下した多型を持つ参加者は、誤りに対する反応での前帯状回ACC活性化の増加およびフランカータスクにおける干渉時の背側ACC活性化の減少を示しています(Holmes et al., 2010)。
中間まとめ
まとめると、現在の証拠は、反応抑制がCSTC回路内の脳領域に依存しており、これらの回路内の活性化が抑制負荷の増加とともに増加することを示唆しています。これらのCSTC回路の適切な機能は、ドーパミン、セロトニン、およびノルアドレナリンの複雑な相互作用に依存しており、ただしそれらの重要性の重みは反応抑制の構成要素によって異なる可能性があります。セロトニンおよびドーパミンレベルの低下は、干渉制御を改善するように見えます。一方、ドーパミンレベルの増加は、行動抑制および行動取消を改善するように見えます。遺伝子関連研究では、主にドーパミンシグナルの減少と関連している多型が、運動反応抑制パフォーマンスの低下と関連していることが報告されてきました。CSTC回路における構造的および機能的異常、およびセロトニン、ノルアドレナリン、およびドーパミン伝達の異常は、強迫性障害(OCD)患者および関連障害患者における反応抑制障害の基礎となる可能性があります。
強迫性障害と抑制
強迫性障害(OCD)は人口の2〜3%に影響を及ぼす不安障害であり、社会的および職業的機能に深刻な障害を引き起こします(Ruscio et al., 2010)。この障害は、不安を引き起こす強迫観念(反復的な侵入的思考)および強迫行為(反復的な儀式的行動)によって特徴づけられます。これらの行動は不安を軽減するために実行されます(American Psychiatric Association, 2013)。これらの症状は一般的であり、人口の25%以上が一生のうちに亜臨床的な強迫観念または強迫行為を経験しています(Ruscio et al., 2010)。OCDの薬物療法は主にSSRIsで構成されており、これはセロトニン作動性系がこの障害の病態生理学に関与していることを示唆しています。ただし、推定ではOCD患者の40〜60%がこの治療に反応せず、セロトニン作動性およびドーパミン作動性の両方のシステムに影響を与える atypical 抗精神病薬による追加治療を必要とします(Denys et al., 2004a; Fineberg et al., 2005)。神経イメージング研究は、中脳、視床、および脳幹におけるSERT可用性の減少および前頭前皮質、頭頂葉、および側頭葉脳領域におけるセロトニン2A受容体可用性の減少を証明することにより、OCDにおけるセロトニン作動性機能不全の概念を強化してきました(Hesse et al., 2005; Perani et al., 2008)。ドーパミン作動性系の異常もOCD患者で観察されており、ストリオイドにおけるDaTレベルの増加およびストリオイドにおけるD1およびD2受容体の可用性の減少が関連しています(Kim et al., 2003; Denys et al., 2004b; van der Wee et al., 2004; Olver et al., 2009)。過去数年間、研究の関心は、OCD症状のモデルとしての反応抑制に焦点を当ててきました(Chamberlain et al., 2005)。これを支持するように、干渉制御の障害(例えば、非合同試行での反応時間の増加)がOCDで記述されています(Bannon et al., 2002; Penades et al., 2007; Nabeyama et al., 2008; Nakao et al., 2009; Schlosser et al., 2010)。OCD研究において機能的神経イメージングとともに干渉制御パラダイムが使用されたいくつかの研究があります(表1を参照;Fitzgerald et al., 2005; Nakao et al., 2005a, 2009; vanden Heuvel et al., 2005; Viard et al., 2005; Nabeyama et al., 2008; Woolley et al., 2008; Page et al., 2009; Schlosser et al., 2010; Huyser et al., 2011; Rubia et al., 2011a; Marsh et al., 2013)。いくつかの研究では、OCDを有する成人および児童における誤りおよび干渉制御後のACCの過活性化が報告されています(Fitzgerald et al., 2005; Huyser et al., 2011)。一方で、他の研究ではACCの低活性化が報告されています(Nakao et al., 2005a; Rubia et al., 2011a)。pre-SMAにおける抑制関連の脳活性化の変化も観察されています(Fitzgerald et al., 2005; Rubia et al., 2011a)および島皮質における変化も観察されています(Huyser et al., 2011)。前頭-線条体領域(IFGおよび被殻を含む)における活性化の増加は、OCD患者においてシモンタスクのパフォーマンス時に観察されています(Marsh et al., 2013)。干渉制御タスクにおける脳活性化の変化は、ネットワークレベルでも観察されています。Schlosser et al. (2010)は、動的因果モデリング(DCM)を使用して、ストループタスクのパフォーマンス時の前頭帯状回ネットワークにおける機能的結合性を調査し、OCD患者において健康な対照群と比較してDLPFCとACCの間の機能的結合性が増加していることを発見しました。さらに、OCD患者においてシモンタスクのパフォーマンス時に、被殻、下頭頂葉皮質、尾状核、視床、および前頭領域の間の機能的結合性の増加が観察されました(Marsh et al., 2013)。OCDにおける行動取消および行動抑制の障害は記述されています(Chamberlain et al., 2007b; Penades et al., 2007)。患者は、健康な対照群と比較して、ストップシグナルタスクでのSSRTの増加(すなわち、抑制の遅れ)およびGo/No-Goタスクでの誤り率の増加を示しています。運動反応抑制の障害は、OCD患者の無影響の一等親戚でも観察されており(Chamberlain et al., 2007b; Menzies et al., 2007)、これは運動反応抑制が遺伝可能な特性(エンドフェノタイプ)であり、家族内で疾患と共に segregate することを示唆しています(Gottesman & Gould, 2003)。OCD患者における運動反応抑制障害の構造的神経相関は特定されています。OCD患者およびその無影響の一等親戚における行動取消の障害は、ACC、被殻、尾状核、扁桃体、頭頂葉領域、および小脳における灰白質体積の増加および眼窩前頭皮質(OFC)、IFG、ACC、前運動皮質、および側頭葉領域における灰白質体積の減少と関連しています(Menzies et al., 2007)。OCD患者における運動反応抑制障害の機能的神経相関も特定されています(表2を参照;Maltby et al., 2005; Rothe et al., 2007; Woolley et al., 2008; Page et al., 2009; Rubia et al., 2010; de Wit et al., 2012; Kang et al., 2012)。CSTC回路における抑制時のタスク関連活性化の減少は、OCD患者で観察されています(Rothe et al., 2007; Woolley et al., 2008; Page et al., 2009; Rubia et al., 2010; de Wit et al., 2012; Kang et al., 2012)。ただし、これらの領域における活性化の増加が報告された研究も1つあります(Maltby et al., 2005)。これまでで最大の研究において、de Wit et al. (2012)は、薬物を服用していないOCD患者において抑制時のIFGおよび下頭頂葉皮質の活性化の減少を発見しました。この過活性化の前頭前野(pre-SMA)は、彼らの無影響の兄弟姉妹でも観察されています。pre-SMAの過活性化は、患者および兄弟姉妹におけるSSRTと負の相関があり、これによりpre-SMAの過活性化が補償メカニズムと考えられることを示唆しています。全体として最も一貫している結果は、OCD患者において抑制時のDLPFC、IFG、被殻、および視床の活性化の減少です(Rothe et al., 2007; Woolley et al., 2008; Page et al., 2009; Rubia et al., 2010; de Wit et al., 2012)。最近の研究では、薬物を服用していない成人のOCD患者、その無影響の兄弟姉妹、および健康な対照群におけるストップシグナルタスクのパフォーマンス時の機能的結合性を調査しました(van Velzen et al., under review)。心理生理学的相互作用(PPI)分析およびDCMを実施したところ、患者およびその兄弟姉妹におけるIFGと扁桃体の間の異常な結合性が見出されました。これにより、この結合性のパターンがエンドフェノタイプである可能性が示唆されました。辺縁系の活性化は、OCDにおけるCSTC回路の活性化に干渉する可能性があります。我々は、抑制時のIFG、pre-SMA、および被殻の間の異常な結合性の証拠を見つけることはできませんでした。これらの結果は、他のサンプルでの再検討を warranted します。OCDの薬理学的治療が反応抑制に与える影響を調査した研究は2つあります。SSRIsによる治療は、干渉制御タスクのパフォーマンス時のタスク関連脳活性化の増加および症状の改善と関連していることが関連しています(Nakao et al., 2005b)。ただし、研究デザインにより、この活性化の変化が症状の改善による二次的なものであるのか、または薬理学的治療によるものであるのかは不明のままです。もう1つの研究では、SSRIsで治療を受けたOCD患者におけるGo/No-Goタスクのパフォーマンス時の複数の皮質および皮質下脳領域の活性化の増加が、SSRIsで治療を受けていないOCD患者と比較して報告されています(Rothe et al., 2007)。ただし、この研究は横断的研究であり、小さな患者群を含んでおり、疾患の重症度との関係を調査していませんでした。
[Image blocked: 表1: 強迫性障害における干渉制御タスクを使用したfMRI研究の概要。]
表1: 強迫性障害における干渉制御タスクを使用したfMRI研究の概要。
| 研究 | タスク | 年齢群 | 参加者 | 薬物および併存疾患 | コントラスト | OCD患者における結果 |
|---|---|---|---|---|---|---|
| Fitzgerald et al. (2005) | フランカータスク | 成人 | OCD患者 8人(女性 2人) 健康対照群 7人(女性 2人) | 3人の患者がSSRIで治療済み、1人がベンゾジアゼピンで治療済み、1人が抗精神病薬で治療済み 3人の患者がうつ病の診断基準を満たす、2人がジストヒミアの診断基準を満たす 重篤な医学的状態、神経学的障害、または頭部外傷なし | E>Corr IC>C” | 前帯状回ACC(症状の重症度と相関)# 前補足運動野 pre-SMA” 両側尾状核 |
| Huyser et al. (2011) | フランカータスク | 児童/思春期 | OCD患者 25人(女性 16人) 健康対照群 25人(女性 16人) | 参加前少なくとも2週間薬物フリー 48%の患者が併存不安障害、12%が併存感情障害、12%がADHD/ODD、8%がチック障害 | E>Corr IC>C” | ACC、島皮質” 両側島皮質 |
| Nakao et al. (2005a) | ストループタスク | 成人 | OCD患者 24人(女性 15人) 健康対照群 14人(女性 9人) | 参加前少なくとも2週間薬物フリー 併存軸I障害なし、重篤な医学的状態なし、神経学的障害なし、頭部外傷なし、物質乱用なし | IC>C” | 前頭葉R.# 両側ACC、側頭葉、尾状核R. |
| Nabeyama et al. (2008) | ストループタスク | 成人 | OCD患者 11人(女性 7人) 健康対照群 19人(女性 11人) | 参加前少なくとも2週間薬物フリー 併存障害未報告 | IC>C# | 右ACC、右小脳 |
| Woolley et al. (2008) | モーターストループタスク | 児童/思春期 | OCD患者 10人(女性 0人) 健康対照群 9人(女性 0人) | 8人の患者がSSRIで治療済み、5人がCBTで治療済み 併存軸I障害なし、神経学的障害なし、頭部外傷なし、重篤な医学的状態 | IC>C# | 中間側頭回、両側小脳 |
| Page et al. (2009) | モーターストループタスク | 成人 | OCD患者 10人(女性 0人) 健康対照群 11人(女性 0人) | 薬物フリー 2人の患者がジストヒミアの診断基準を満たす、3人が以前にうつ病の診断基準を満たしていた、1人が以前にアルコール依存症の診断基準を満たしていた | IC>C” | 左小脳、左後帯状回# 両側楔前部、右側頭葉、左側頭頂葉接合部 |
| Nakao et al. (2009) | ストループタスク | 成人 | OCD患者: 病歴<10年 12人(女性 12人) OCD患者: 病歴>10年 8人(女性 8人) 健康対照群 16人(女性 7人) | 参加前少なくとも2週間薬物フリー 併存軸I障害なし、重篤な医学的状態なし、神経学的障害なし、頭部外傷なし、物質乱用なし | IC>C# | 病歴<10年の患者における尾状核、小脳 病歴>10年の患者および対照群と比較して |
| (続き) | ||||||
| Schlosser et al. (2010) | ストループタスク | 成人 | OCD患者 21人(女性 16人) 健康対照群 21人(女性 16人) | 参加前少なくとも2日間薬物フリー 併存軸I障害なし、精神病性なし、神経学的障害なし | IC>CIC” | 両側DLPFC” 両側上前頭回、背側ACC、左前中央回、右上頭頂葉、右下頭頂葉 |
| vanden Heuvel et al. (2005) | ストループタスク | 成人 | OCD患者 18人(女性 12人) 対照群 19人(女性 9人) | 参加前少なくとも4週間薬物フリー 神経学的疾患なし、他の精神医学的障害なし | IC>C” | 右楔前部、左側海馬回、左脳幹 |
| Viard et al. (2005) | 対立タスク | 成人 | OCD患者 12人(女性 5人) 健康対照群 15人(女性 4人) | 11人の患者がSSRIで治療済み、1人がまたTCAで治療済み 併存障害なし、重篤な医学的状態なし、神経学的障害なし、頭部外傷なし | IC>N | 脳活性化に差なし |
| Marsh et al. (2013) | シモンタスク | 成人 | OCD患者 22人(女性 11人) 健康対照群 22人(女性 11人) | 薬物フリー 5人の患者が生涯にわたってうつ病の既往あり | IC>C” | 右IFG、島皮質、および被殻 |
| Rubia et al. (2011a) | シモンタスク | 児童/思春期 | OCD患者 10人(女性 0人) 健康対照群 20人(女性 0人) | 8人の患者がSSRIで治療済み;5人がCBTで治療済み 併存精神医学的障害なし、学習障害の既往なし、物質乱用なし | IC>oddball# | 右pre-SMA、ACC、上頭頂葉皮質 |
| ADHD、注意欠如・多動性障害;C、合同試行;CBT、認知行動療法;corr、正解試行;DLPFC、背外側前頭前皮質;E、誤試行;f、女性;IC、非合同試行;ODD、反抗性挑戦障害;OFC、眼窩前頭皮質;SMA、補足運動野;SSRI、選択的セロトニン再取り込み阻害薬;STG、上側頭葉;TCA、三環系抗うつ薬。患者および左pre-SMAの活性化の増加。このpre-SMAの過活性化は、彼らの無影響の兄弟姉妹でも見られました。pre-SMAの活性化は、患者および兄弟姉妹におけるSSRTと負の相関があり、これによりpre-SMAの過活性化が補償メカニズムと考えられることを示唆しています。全体として最も一貫している結果は、OCD患者において抑制時のDLPFC、IFG、被殻、および視床の活性化の減少です(Rothe et al., 2007; Woolley et al., 2008; Page et al., 2009; Rubia et al., 2010; de Wit et al., 2012)。最近の研究では、薬物を服用していない成人のOCD患者、その無影響の兄弟姉妹、および健康な対照群におけるストップシグナルタスクのパフォーマンス時の機能的結合性を調査しました(van Velzen et al., under review)。心理生理学的相互作用(PPI)分析およびDCMを実施したところ、患者およびその兄弟姉妹におけるIFGと扁桃体の間の異常な結合性が見出されました。これにより、この結合性のパターンがエンドフェノタイプである可能性が示唆されました。辺縁系の活性化は、OCDにおけるCSTC回路の活性化に干渉する可能性があります。我々は、抑制時のIFG、pre-SMA、および被殻の間の異常な結合性の証拠を見つけることはできませんでした。これらの結果は、他のサンプルでの再検討を warranted します。OCDの薬理学的治療が反応抑制に与える影響を調査した研究は2つあります。SSRIsによる治療は、干渉制御タスクのパフォーマンス時のタスク関連脳活性化の増加および症状の改善と関連していることが関連しています(Nakao et al., 2005b)。ただし、研究デザインにより、この活性化の変化が症状の改善による二次的なものであるのか、または薬理学的治療によるものであるのかは不明のままです。もう1つの研究では、SSRIsで治療を受けたOCD患者におけるGo/No-Goタスクのパフォーマンス時の複数の皮質および皮質下脳領域の活性化の増加が、SSRIsで治療を受けていないOCD患者と比較して報告されています(Rothe et al., 2007)。ただし、この研究は横断的研究であり、小さな患者群を含んでおり、疾患の重症度との関係を調査していませんでした。 |
[Image blocked: 表2: 強迫性障害における反応抑制パラダイムを使用したfMRI研究の概要。]
表2: 強迫性障害における反応抑制パラダイムを使用したfMRI研究の概要。
| 研究 | タスク | 年齢群 | 参加者 | 薬物および併存疾患 | コントラスト | OCD患者における結果 |
|---|---|---|---|---|---|---|
| Maltby et al. (2005) | Go/No-Goタスク | 成人 | OCD患者 11人(女性 7人) 健康対照群 11人(女性 7人) | 薬物フリー;OCDが主要診断、他の軸I障害の基準を満たす患者が6人 精神病性なし、神経学的障害なし、頭部外傷なし、物質乱用なし | FS>Go SS>Go” | 外側前頭前皮質、ACC、外側眼窩前頭皮質、尾状核、視床 失敗時および成功時の抑制中 |
| Rothe et al. (2007) | Go/No-Goタスク | 成人 | OCD患者 12人(女性 7人) 健康対照群 14人(女性 8人) | 6人の患者がSSRIで治療済み 2人の患者がうつ病の診断基準を満たす、1人が社会恐怖症の診断基準を満たす 神経学的障害なし、頭部外傷なし、重篤な医学的状態、物質乱用なし | NoGo>Go# | 右IFG、右中前頭前皮質 |
| Woolley et al. (2008) | ストップシグナルタスク | 児童/思春期 | OCD患者 10人(女性 0人) 健康対照群 9人(女性 0人) | 8人の患者がSSRIで治療済み、5人がCBTで治療済み 併存軸I障害なし、神経学的障害なし、頭部外傷なし、重篤な医学的状態 | SS>FS FS>Go# | 右眼窩前頭皮質、視床、基底核# DLPFC、側頭葉活性化 |
| Page et al. (2009) | Go/No-Goタスク | 成人 | OCD患者 10人(女性 0人) 健康対照群 11人(女性 0人) | 薬物フリー 2人の患者がジストヒミアの診断基準を満たす、3人が以前にうつ病の診断基準を満たしていた、1人が以前にアルコール依存症の診断基準を満たしていた | NoGo>Go NoGo>Go” | 後帯状回VMPFC、前運動皮質、小脳# 眼窩前頭皮質、DLPFC、ACC、被殻、海馬、視床 |
| Rubia et al. (2010) | ストップシグナルタスク | 思春期 | OCD患者 10人(女性 0人) 健康対照群 20人(女性 0人) | 患者が治療を受け、部分寛解状態 主要な精神医学的障害なし、物質乱用なし、学習障害の既往なし | SS>Go FS>Go# | 右眼窩前頭皮症(症状改善と相関)# 左中前頭前皮質de Wit et al. (2012) |
| Kang et al. (2012) | ストップシグナルタスク | 成人 | OCD患者 18人(女性 6人) 健康対照群 18人(女性 6人) | 薬物フリー 主要な精神医学的障害なし、精神病性なし、神経学的疾患なし、物質乱用なし、うつ病および知的障害あり | SS>Go SS>Go” | 両側上頭頂葉皮質、小脳、右側海馬回皮質、左前中央回、右回状結合織、被殻および側頭葉、中後頭葉皮質、左角回、小脳および帯状回 |
| ACC、前帯状回;CBT、認知行動療法;DLPFC、背外側前頭前皮質;f、女性;FS、失敗停止;IFG、下前頭回;MTL、中間側頭葉;OFC、眼窩前頭皮質;SS、成功停止;SSRI、選択的セロトニン再取り込み阻害薬;VMPFC、腹側前頭前皮質。 |
まとめると、OCD患者は干渉制御および運動反応抑制の両方において障害を示します。前頭前皮質およびCSTC回路内の他の脳領域は、干渉制御時に過活性化しているように見えますが、結果は一貫していません。行動抑制および行動取消時の抑制負荷が増加すると、CSTC領域は一般的に対照群と比較して低活性化になりますが、一部の補償が生じる可能性があります。CSTC回路におけるセロトニンの減少およびドーパミンの増加は、OCDにおける反応抑制障害の基礎となっている可能性があります。反応抑制障害の存在および機能的相関は、OCDと関連する障害(トゥレット症候群(TS)、抜毛症(TTM)、および注意欠如・多動性障害(ADHD))においても調査されてきました。これにより、これらの認知的機能不全の障害特異性が可能となり、これらの障害間での抑制障害の比較が可能となります。
他の前頭-線条体障害における抑制
トゥレット症候群
ジル・ド・ラ・トゥレット症候群(トゥレット症候群とも呼ばれる)は、運動チックおよび声帯チックによって特徴づけられる神経発達障害です(American Psychiatric Association, 2013)。TSは人口の0.4〜1%に影響を及ぼします(Swain et al., 2007; Robertson, 2008)。OCDと同様に、セロトニン作動性およびドーパミン作動性システムの機能不全がTSの病態生理学に関与していることが示唆されています(レビューについては、Steeves & Fox (2008) を参照)。いくつかの臨床試験では、ドーパミン拮抗薬(リスペリドンおよびハロペリドールなど)の投与が、ほとんどの患者におけるチックの抑制に効果的であることが示されています(Blochetal., 2011; Roessneretal., 2011)。神経イメージング研究では、皮質(眼窩前頭皮質、前帯状回、島皮質、側頭葉、および後頭葉皮質)および皮質下領域(視床および海馬)におけるD2およびD3受容体の可用性の減少が報告されています(Gilbertetal., 2006; Steevesetal., 2010)およびストリオイドにおけるDaT可用性の増加が報告されています(Malisonetal., 1995; Muller-Vahletal., 2000; Cheonetal., 2004; Serra-Mestresetal., 2004; Liuetal., 2010)。ただし、相反する結果も報告されています(Singeretal., 2002; Hwangetal., 2008)。TSにおけるセロトニン作動性システムの神経イメージングでは、皮質(眼窩前頭皮質、前帯状回、島皮質、側頭葉、後頭葉皮質、および後頭葉皮質)および皮質下領域(視床、尾状核、および海馬)におけるセロトニン2A受容体の結合増加が報告されています(Haugboletal., 2007)およびストリオイドおよび中脳におけるSERT可用性の増加が報告されています(Wongetal., 2008)。TSにおける前頭-線条体機能不全の証拠が増加しています(レビューについては、Albin & Mink (2006) および Felling & Singer (2011) を参照)。例えば、チック抑制時のCSTC回路の活性化の程度と症状の重症度は負の相関があります(Petersonetal., 1998)。また、前頭前皮質の厚さ(Draganskietal., 2010)および前頭CSTC領域の体積は、TS患者において健康な対照群と比較して減少しています(Draganskietal., 2010)。TS患者の90%以上が併存精神医学的障害を有しており、最も一般的にはOCDまたはADHDです(Robertson, 2011)。TS患者の45〜60%がOCDを併存していると推定されています(Ghanizadeh & Mosallaei, 2009)。OCDと同様に、TSにおける不随意運動症状が運動反応抑制および干渉制御に関連しているかどうかを調査した研究は多数あります。その結果は混在しています。一部の研究では障害されたパフォーマンスが報告されています(Baron-Cohenetal., 1994; Crawfordetal., 2005; Rankinetal., 2006; Channoneletal., 2009; Eicheleetal., 2010)。ただし、これは特に抑制負荷が増加した場合に顕著であり、他の研究では障害が報告されていません(Ozonoffetal., 1994; Ozonoff&Jensen, 1999; Hersheyetal., 2004; Verteetal., 2005; Watkinsetal., 2005; Channoneletal., 2006; RayLietal., 2006; Marshetal., 2007; Razetal., 2009; Sukhodolskyetal., 2010)。これらの研究はしばしばTS患者の併存障害および精神薬物療法の使用を含みます。4つの研究におけるストップシグナルタスクを使用したメタ分析では、軽度の抑制障害が見出されました(Lipszyc & Schachar, 2010)。行動的障害の証拠は明確ではありませんが、抑制関連の脳活性化はTSにおいて変化しているようです(表3を参照)。年齢とともに、TS患者は健康な対照群と比較して干渉制御時にCSTC領域の動員が増加しています(Razetal., 2009)。TS患者における干渉制御時にCSTC領域の過活性化が観察されており、これが補償メカニズムと考えられる可能性があります(Marshetal., 2007)。運動反応抑制時に、TS患者はイベント関連電位(ERP)研究において抑制関連の前頭脳活動の増加を示しています(Johannesetal., 2001)。著者らは、この補償的な脳活性化が、TS患者における反応抑制障害が一貫して観察されていない理由を説明する可能性があると指摘しています。Go/No-Goタスクのパフォーマンス時における脳活性化に差は見られませんでしたが、サンプルサイズは限定的でした(Hersheyetal., 2004)。TSにおける薬理学的治療が行動または機能的反応抑制の尺度に与える直接的な影響を調査した研究はまだありませんが、Wylie et al. (2013) を参照してください。まとめると、行動抑制の障害はTSの一部のサブグループに限定される可能性があります。これは、抑制時の補償的な脳活性化が、一部の患者における行動抑制の障害を conceal する可能性があるためです。CSTC回路におけるドーパミンの増加は、TSにおける反応抑制障害の基礎となっている可能性があります。
抜毛症
抜毛症(TTM)は強迫性スペクトラム障害です(American Psychiatric Association, 2013)。この障害を有する患者は、髪を引っ張る衝動を経験し、これが苦痛および機能障害を引き起こします。TTMとOCDの類似性から、TTMは歴史的にSSRIsで治療されてきました。ただし、当初はTTMに効果的であると考えられていましたが(Steinetal., 1997)、最近の研究ではSSRIsがTTMに対して効果的でないことが示されています(StreichenweinandThornby, 1995; vanMinnenetal., 2003)またはTTM患者の特定のサブグループにのみ効果的であることが示されています(Stanleyetal., 1997a; Gaddeetal., 2007)。最近の臨床試験では、異型抗精神病薬(オランザピンおよびアリピプラゾールなど)による治療が、セロトニン作動性およびドーパミン作動性システムに影響を与えることにより、より有望であることが示されています(VanAmeringenetal., 2010; WhiteandKoran, 2011)。TTMの症状はCSTC回路の機能不全に由来すると推測されています(Mataix-Cols & vandenHeuvel, 2006)。この仮説を支持するように、TTMにおける前頭領域および線条体領域の構造的異常が観察されています(Chamberlainetal., 2008)。TTM患者は、運動反応を抑制すること(すなわち、髪を引っ張ること)に困難をきたし、これが反応抑制障害に関与している可能性があります(Chamberlainetal., 2006a)。TTMにおける反応抑制の研究は限られており、結果は混在しています。関連する認知制御タスクのパフォーマンスは変更されていませんが、TTM患者はストループタスクにおける干渉制御の障害を示しています(Stanleyetal., 1997b; Bohneetal., 2005)。行動取消の障害が報告されており、その障害の程度は疾患の重症度と相関しています(Chamberlainetal., 2006a)。行動抑制の障害は、発症早期の患者の特定のサブグループに限定されているようです(Bohneetal., 2008)。TTMにおける反応抑制障害の神経または薬理学的基盤はまだ完全に解明されていません。これは、TTM患者群において抑制関連の神経イメージング研究がまだ実施されていないためです。さらに、TTMにおける薬理学的治療が反応抑制に与える影響を調査した研究もまだありません。ただし、TTM患者は、抑制関連の脳領域における構造的異常を示しています。例えば、線条体、IFG、補足運動野(SMA)、および前頭領域において(Grachev, 1997; O’Sullivanetal., 1997; Chamberlainetal., 2008)。これらは、TTMにおける反応抑制障害の基礎となっている可能性があります。
注意欠如・多動性障害
注意欠如・多動性障害(ADHD)は、多動、不注意、および衝動性によって特徴づけられる神経精神障害です(American Psychiatric Association, 2013)。この障害は一般的であり、学齢期の児童のほぼ10%に影響を及ぼすと考えられています(Froehlichetal., 2007)。ADHDの神経薬理学は複雑であり、まだ十分に理解されていません。現在の証拠は、ADHDがノルアドレナリンおよびドーパミン作動性システムの障害によって特徴づけられていることを示唆しています(レビューについては、McAlonanetal. (2009) を参照)。ただし、一部の研究ではセロトニン作動性系の追加的関与を示唆しています(Oadesetal., 2002)。ADHDに対する薬物療法(メチルフェニデート、アンフェタミン、またはアトモキセチンによる)は、症状の治療に効果的であることが示されています。これは、細胞外ドーパミンおよびノルアドレナリンレベルの増加によるものと考えられています(Prince, 2008 でレビューを参照)。ADHD患者は、干渉制御タスクのいくつか(シモンタスクおよびフランカータスクを含む)において行動的障害を示しています(Rubiaetal., 2011a; Sebastianetal., 2012)。ADHD患者における干渉制御時に、ACC、IFG、視床、SMA、被殻、および下頭頂葉皮質の活性化が減少していることが観察されています(表4を参照)。最近のメタ分析では、ADHD患者におけるストループタスクおよびシモンタスクのパフォーマンス時に、CSTC領域(右IFG、島皮質、右尾状核、左下頭頂葉皮質、および左ACC)の活性化が減少していることが明らかになりました(Hartetal., 2012)。また、前頭-線条体結合性の減少および前頭-頭頂葉結合性の変化が、ADHDにおける反応抑制障害に関与している可能性があります(Cubilloetal., 2010)。薬理学的研究では、メチルフェニデートおよびアトモキセチンの投与が、行動取消を改善することが示されています(Aronetal., 2003a; Chamberlainetal., 2007a; DeVitoetal., 2009; Coghilletal., 2013)および行動保持を改善することが示されています(Vaidyaetal., 1998)。これにより、ノルアドレナリンおよびドーパミンの障害が、ADHDにおける運動反応抑制障害の基礎となっている可能性が示唆されます。さらに、メチルフェニデートの投与は、ADHD患者におけるGo/No-Goタスクのパフォーマンス時の前頭葉および線条体領域の活性化の増加を示しています(Vaidyaetal., 1998)。メチルフェニデートはまた、ADHD患者におけるストップシグナルタスクのパフォーマンス時の前頭葉の未活性化を正常化します(Rubiaetal., 2011b)。アトモキセチンおよびメチルフェニデートの効果を直接比較した研究では、両薬物がストップシグナルタスクのパフォーマンス時の左前頭葉の未活性化を正常化することが示されています。ただし、右前頭葉の未活性化の正常化はメチルフェニデートのみによって行われます(Cubilloetal., 2014)。ADHDにおける反応抑制障害と関連しているいくつかの候補遺伝子が研究で報告されています。ADHD患者において、ドーパミンD4受容体遺伝子(DRD4)の多型(機能活動の減少と関連しています;Asgharietal., 1995)は、抑制成分のあるタスクでのパフォーマンスの変更と関連していることが関連しています(Langleyetal., 2004; Bellgroveetal., 2005;ただし、ストループタスクでのパフォーマンスの低下と関連していることが関連しています;Looetal., 2008)。また、ADHD患者において、ドーパミントランスポーター遺伝子(DaT gene)の多型(発現の増加と関連している)は、調整されたGo/no-gotaskにおける前頭葉および線条体領域の脳活性化の増加および側頭葉、前運動皮質、および頭頂葉皮質の脳活性化の増加と関連しています(Brookesetal., 2011;ただし、機能活動の減少と関連している多型では、線条体領域の活性化の増加が関連しています;Durstonetal., 2008)。さらに、MAO-A遺伝子の多型(MAO-A活性の低下と関連している)は、ADHD患者におけるストップシグナルタスク時の右IFGの活性化の減少と関連していることが関連しています(Nymbergetal., 2013)。ただし、ADHD患者におけるMAO-A遺伝子の遺伝子型は、干渉抑制とは関連していないことが関連しています(Liuetal., 2011)。まとめると、ADHDにおいて、干渉制御および運動反応抑制の両方における行動的障害が顕著であり、これはCSTC領域における体積の減少および低活性化と関連しています。遺伝子関連研究の結果は、抑制パフォーマンスの低下が、CSTC回路におけるドーパミン伝達の減少と関連している可能性を示唆しています。
障害間の比較と統合
議論されたすべての障害は、特定の衝動または反応を抑制できないことを示す症状を示しています。したがって、反応抑制タスクは、これらの障害の一部の症状に寄与する神経底盤を研究するための非常に適切な操作化です。これまでにレビューしてきた文献から、全体として、強迫性障害または関連障害を有する患者は、抑制障害を示し、これはタスク関連の脳活性化の変化と伴っていることを結論付けることができます。これらの脳領域が健康な対照群と比較して過活性化か低活性化かは、主にタスクの複雑さに依存します。一般的に、患者は抑制関連の脳領域を追加で動員することで行動を補償しています。これにより、行動パフォーマンスはしばしば正常ですが、これはあまり複雑でないタスク(例えば、フランカータスクおよびシモンタスク)においてのみ成り立ちます。抑制負荷が増加すると(例えば、Go/no-goおよびStop-signalタスクにおいて)、これらの補償メカニズムが機能不全となり、患者は行動的障害を示し始め、抑制関連の神経回路の活性化が減少します。この現象は健康な対照群でも観察されています(Sebastianetal., 2013a)。ただし、健康な対照群はより高い抑制負荷までストレスを与えることができますが、パフォーマンスの低下を伴います。つまり、患者は健康な対照群よりも低い抑制負荷において、行動パフォーマンスの障害および補償的な活性化の失敗を示します。この現象は、OCD患者およびその無影響の兄弟姉妹、およびADHD患者においても観察されています。
[Image blocked: 図2: タスク負荷と抑制関連神経回路活性の逆U字型関係におけるシフト。抑制関連神経回路活性はタスク負荷とともに徐々に増加します(緑から赤へのグラデーション)。ただし、タスクの要求が高すぎると、補償的活性が機能不全となり(実線赤)、行動パフォーマンスが障害されます。強迫性障害または関連障害を有する患者において、行動パフォーマンスの障害および補償的神経回路活性の失敗は、健康な対照群よりも低いタスク負荷で発生します(これは、逆U字型曲線を左にシフトさせることで視覚化されます)。]
図2: タスク負荷と抑制関連神経回路活性の逆U字型関係におけるシフト。抑制関連神経回路活性はタスク負荷とともに徐々に増加します(緑から赤へのグラデーション)。ただし、タスクの要求が高すぎると、補償的活性が機能不全となり(実線赤)、行動パフォーマンスが障害されます。強迫性障害または関連障害を有する患者において、行動パフォーマンスの障害および補償的神経回路活性の失敗は、健康な対照群よりも低いタスク負荷で発生します(これは、逆U字型曲線を左にシフトさせることで視覚化されます)。
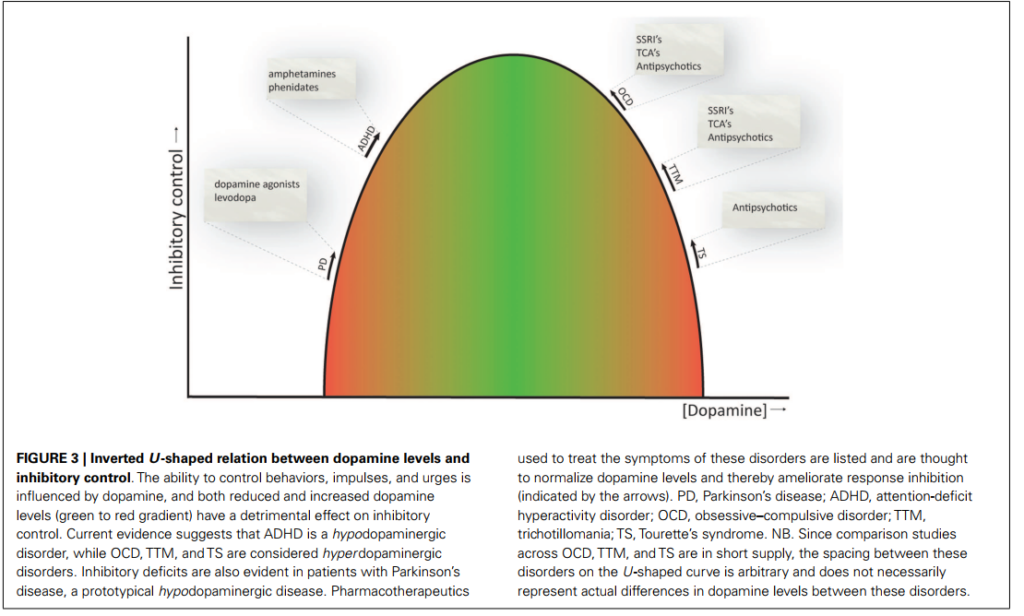
[Image blocked: 図3: ドパミンレベルと抑制制御の逆U字型関係。行動、衝動、および切迫感を制御する能力はドパミンによって影響を受けます。ドパミンレベルの低下および増加(緑から赤へのグラデーション)の両方が、抑制制御に悪影響を及ぼします。現在の証拠は、ADHDが低ドパミン作動性障害であることを示唆しており、一方、OCD、TTM、およびTSは高ドパミン作動性障害であることを示唆しています。抑制制御の障害は、パーキンソン病患者においても見られており、これは典型的な低ドパミン作動性障害です。パーキンソン病の症状を治療するために使用される薬物療法は、ドパミンレベルを生理学的レベルに正常化すると考えられており、それによって抑制制御を改善すると考えられています(矢印で示されます)。PD、パーキンソン病;ADHD、注意欠如・多動性障害;OCD、強迫性障害;TTM、抜毛症;TS、トゥレット症候群。NB. OCD、TTM、およびTS間の比較研究が限られているため、これらの障害のU字型曲線間の間隔は任意であり、これらの障害間の実際のドパミンレベルの違いを必ずしも反映しているわけではありません。]
図3: ドパミンレベルと抑制制御の逆U字型関係。行動、衝動、および切迫感を制御する能力はドパミンによって影響を受けます。ドパミンレベルの低下および増加(緑から赤へのグラデーション)の両方が、抑制制御に悪影響を及ぼします。現在の証拠は、ADHDが低ドパミン作動性障害であることを示唆しており、一方、OCD、TTM、およびTSは高ドパミン作動性障害であることを示唆しています。抑制制御の障害は、パーキンソン病患者においても見られており、これは典型的な低ドパミン作動性障害です。パーキンソン病の症状を治療するために使用される薬物療法は、ドパミンレベルを生理学的レベルに正常化すると考えられており、それによって抑制制御を改善すると考えられています(矢印で示されます)。PD、パーキンソン病;ADHD、注意欠如・多動性障害;OCD、強迫性障害;TTM、抜毛症;TS、トゥレット症候群。NB. OCD、TTM、およびTS間の比較研究が限られているため、これらの障害のU字型曲線間の間隔は任意であり、これらの障害間の実際のドパミンレベルの違いを必ずしも反映しているわけではありません。
低ドパミン作動性障害としてのADHDと、高ドパミン作動性障害としてのOCD、TTM、およびTSの考え方は、現在利用可能な薬物療法と一致しています。これらの薬物療法の神経生物学的メカニズムは、ドパミンレベルを生理学的レベルに戻すことに依存していると考えられています(Abi-Dargham & Laruelle, 2005; Gerlachetal., 2013)。さらに、OCDの第一選択薬物療法であるSSRIsおよび三環系抗うつ薬は、セロトニンシグナルのアップレギュレーションによりドパミンレベルを正常化する可能性があります。これにより、ドパミンに抑制効果がもたらされます(Boureau & Dayan, 2010)。図3は、ドパミンレベルと抑制制御の間の提案された関係の概念的表現を提供します。この関係は、ドパミンレベルと作業記憶機能の逆U字型関係(Cools & D’Esposito, 2011)と類似しています。ただし、提案された関係は完全な物語を提供するわけではなく、このレビューで議論されている一部の結果を理解するための枠組みとしてのみ意図されています。ドパミンは前頭前皮質および線条体において異なる効果を持ち、異なる発火モード(すなわち、トニックおよびファasic)およびセロトニン、ノルアドレナリン、およびグルタミン酸システム、さらにエストロゲンなどのホルモンとの高度に複雑な相互作用があります(Boureau & Dayan, 2010; Cools & D’Esposito, 2011; de Bartolomeisetal., 2013)。これにより、これらの神経伝達物質の脳活動および行動への影響を明確に理解することは困難になります。要約すると、強迫性スペクトラム障害における反応抑制の機能的および行動的障害は、タスク要求と抑制関連神経回路活性の関係におけるシフトとして概念化することができます。このシフトの原因およびこれがこれらの障害の症状の基礎となっている可能性は現在不明ですが、ドパミンが重要な役割を果たしていることを postul しました。
結論と今後の方向性
本レビューの目的は、強迫性スペクトラム障害(OCD、ADHD、TS、およびTTM)における抑制障害の神経的、薬理学的、および遺伝的基盤を調査した研究の概要を提供することであり、特にOCDに焦点を当てました。我々は、機能的および行動的に障害された反応抑制がこれらの障害間で共有されている特徴であり、これらの障害の症状に関連する機能不全の少なくとも一部の基礎となっていることを示してきました。神経イメージング研究は、抑制関連の脳領域がADHDおよびOCDにおいて一般的に低活性化であることを示唆しています(ただし、これはタスク負荷に依存します)。一方、TSにおける研究では混合した結果が得られています。現時点では、TTMにおける反応抑制の神経相関に関する研究はまだ出版されていないことを私たちの知る限りでは確認しています。ドパミンおよびセロトニンシグナルは、反応抑制において重要であり、これらの神経伝達物質の機能不全は、強迫性スペクトラム障害において頻繁に観察されています。ただし、ほとんどの神経伝達物質に関する神経イメージング研究は、(慢性)薬物療法を受けている患者を対象として行われてきました。これにより、薬物と放射性リガンドの競合による直接的な影響、または脳が薬物療法の効果に適応することによる間接的な影響(例えば、特定の結合部位での影響)が生じる可能性があります(Wang et al., 2013)。疾患そのものの病態生理学をより深く理解し、新しい治療標的を特定するためには、薬物を服用していない患者を対象としたさらなる研究が必要です。これらの患者を治療開始後に追跡調査することにより、治療が反応抑制障害および障害特異的な症状に与える影響についての洞察を得ることができます。最後に、関連する精神医学的障害間における抑制障害の病態生理学を比較する研究は相対的に不足しています。このような研究を行うことにより、衝動性および強迫性症状に関連する共通および特異的な疾患バイオマーカーを特定することが可能になります。
著者の貢献
すべての著者は本レビューの設計に関与しました。Laura S. van VelzenおよびChris Vriendは原稿の最初のバージョンを作成しました。すべての著者は文献の解釈に貢献し、以前のドラフトを修正し、最終バージョンの原稿およびその提出を承認しました。
謝辞
本論文は、Brain and Behavior Research FoundationのNARSAD若手研究者賞(Odile A. vanden Heuvel博士に授与)およびオランダ科学研究機構(NWO-ZonMw VENI助成金916.86.036、Odile A. vanden Heuvel博士)による助成金によって支援されました。
参考文献
Abi-Dargham, A., & Laruelle, M. (2005). Mechanisms of action of second-generation antipsychotic drugs in schizophrenia: insights from brain imaging studies. European Psychiatry, 20, 15–27. doi:10.1016/j.eurpsy.2004.11.003
Albin, R. L., & Mink, J. W. (2006). Recent advances in Tourette syndrome research. Trends in Neuroscience, 29, 175–182. doi:10.1016/j.tins.2006.01.001
Alderson, R. M., Rapport, M. D., & Kofler, M. J. (2007). Attention-deficit/hyperactivity disorder and behavioral inhibition: a meta-analytic review of the stop-signal paradigm. Journal of Abnormal Child Psychology, 35, 745–758. doi:10.1007/s10802-007-9131-6
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annual Review of Neuroscience, 9, 357–381. doi:10.1146/annurev.ne.09.030186.002041
American Psychiatric Association. (2013). Diagnostic and Statistical Manual of Mental Disorders, 5th Edition. Arlington: American Psychiatric Publishing.
Anderson, I. M., Clark, L., Elliott, R., Kulkarni, B., Williams, S. R., & Deakin, J. F. (2002). 5-HT(2C) receptor activation by m-chlorophenylpiperazine detected in humans with fMRI. Neuroreport, 13, 1547–1551. doi:10.1097/00001756-200208270-00012
Aron, A. R. (2011). From reactive to proactive and selective control: developing a richermodel for stopping inappropriate responses. Biological Psychiatry, 69, e55–e68. doi:10.1016/j.biopsych.2010.07.024
Aron, A. R., Dowson, J. H., Sahakian, B. J., & Robbins, T. W. (2003a). Methylphenidate improves response inhibition in adults with attention-deficit/hyperactivity disorder. Biological Psychiatry, 54, 1465–1468. doi:10.1016/S0006-3223(03)00609-7
Aron, A. R., Fletcher, P. C., Bullmore, E. T., Sahakian, B. J., & Robbins, T. W. (2003b). Stop-signal inhibition disrupted by damage to right inferior frontal gyrus in humans. Nature Neuroscience, 6, 115–116. doi:10.1038/nn1003
Asghari, V., Sanyal, S., Buchwaldt, S., Paterson, A., Jovanovic, V., & VanTol, H. H. (1995). Modulation of intracellular cyclic AMP levels by different human opamine D4 receptor variants. Journal of Neurochemistry, 65, 1157–1165. doi:10.1046/j.1471-4159.1995.65031157.x
Baehne, C. G., Ehlis, A. C., Plichta, M. M., Conzelmann, A., Pauli, P., Jacob, C., et al. (2009). TPH2 gene variants modulate response control processes in adult ADHD patients and healthy individuals. Molecular Psychiatry, 14, 1032–1039. doi:10.1038/mp.2008.39
Banich, M. T., Burgess, G. C., Depue, B. E., Ruzic, L., Bidwell, L. C., Hitt-Laustsen, S., et al. (2009). The neural basis of sustained and transient attentional control in young adults with ADHD. Neuropsychologia, 47, 3095–3104. doi:10.1016/j.neuropsychologia.2009.07.005
Bannon, S., Gonsalvez, C. J., Croft, R. J., & Boyce, P. M. (2002). Response inhibition deficits in obsessive-compulsive disorder. Psychiatry Research, 110, 165–174. doi:10.1016/S0165-1781(02)00104-X
Bari, A., Eagle, D. M., Mar, A. C., Robinson, E. S., & Robbins, T. W. (2009). Dissociable effects of noradrenaline, dopamine, and serotonin uptake blockade on stop task performance in rats. Psychopharmacology (Berl.), 205, 273–283. doi:10.1007/s00213-009-1537-0
Bari, A., & Robbins, T. W. (2013). Inhibition and impulsivity: behavioral and neural basis of response control. Progress in Neurobiology, 108, 44–79. doi:10.1016/j.pneurobio.2013.06.005
Baron-Cohen, S., Cross, P., Crowson, M., & Robertson, M. (1994). Can children with Gilles de la Tourette syndrome edit their intentions? Psychological Medicine, 24, 29–40. doi:10.1017/S0033291700026805
Batty, M. J., Liddle, E. B., Pitiot, A., Toro, R., Groom, M. J., Scerif, G., et al. (2010). Cortical gray matter in attention-deficit/hyperactivity disorder: a structural magnetic resonance imaging study. Journal of the American Academy of Child & Adolescent Psychiatry, 49, 229–238. doi:10.1097/00004583-201003000-00006
Bedard, A. C., Schulz, K. P., Cook, E. H. Jr., Fan, J., Clerkin, S. M., Ivanov, I., et al. (2010). Dopamine transporter gene variation modulates activation of striatum in youth with ADHD. Neuroimage, 53, 935–942. doi:10.1016/j.neuroimage.2009.12.041
Bellgrove, M. A., Hawi, Z., Lowe, N., Kirley, A., Robertson, I. H., & Gill, M. (2005). DRD4 gene variants and sustained attention in attention-deficit/hyperactivity disorder (ADHD): effects of associated alleles at the VNTR and -521 SNP. American Journal of Medical Genetics. B Neuropsychiatr. Genet., 136, 81–86. doi:10.1002/ajmg.b.30193
Blasi, G., Goldberg, T. E., Weickert, T., Das, S., Kohn, P., Zoltick, B., et al. (2006). Brain regions underlying response inhibition and interference monitoring and suppression. European Journal of Neuroscience, 23, 1658–1664. doi:10.1111/j.1460-9568.2006.04680.x
Bloch, M., State, M., & Pittenger, C. (2011). Recent advances in Tourette syndrome. Current Opinion in Neurology, 24, 119–125. doi:10.1097/WCO.0b013e328344648c
Bohne, A., Keuthen, N. J., Tuschen-Caffier, B., & Wilhelm, S. (2005). Cognitive inhibition in trichotillomania and obsessive-compulsive disorder. Behavioral Research and Therapy, 43, 923–942. doi:10.1016/j.brat.2004.06.014
Bohne, A., Savage, C. R., Deckersbach, T., Keuthen, N. J., & Wilhelm, S. (2008). Motor inhibition in trichotillomania and obsessive-compulsive disorder. Journal of Psychiatric Research, 42, 141–150. doi:10.1016/j.jpsychires.2006.11.008
Booth, J. R., Burman, D. D., Meyer, J. R., Lei, Z., Trommer, B. L., Davenport, N. D., et al. (2005). Larger deficits in brain networks for response inhibition than for visual selective attention in attention-deficit/hyperactivity disorder (ADHD). Journal of Child Psychology and Psychiatry, 46, 94–111. doi:10.1111/j.1469-7610.2004.00337
Boureau, Y.-L., & Dayan, P. (2010). Opponency revisited: competition and cooperation between dopamine and serotonin. Neuropsychopharmacology, 36, 74–97. doi:10.1038/npp.2010.151
Braet, W., Johnson, K. A., Tobin, C. T., Acheson, R., McDonnell, C., Hait, Z., et al. (2011). fMRI activation during response inhibition and error processing: the role of the DAT1 gene in typically developing adolescents and those diagnosed with ADHD. Neuropsychologia, 49, 1641–1650. doi:10.1016/j.neuropsychologia.2011.01.001
Brookes, K. J., Neale, B. M., Sugden, K., Khan, N., Asherson, P., & D’Souza, U. M. (2007). Relationship between VNTR polymorphism of the human dopamine transporter gene and expression in post-mortem midbrain tissue. American Journal of Medical Genetics. B Neuropsychiatr. Genet., 144, 1070–1078. doi:10.1002/ajmg.b.30572
Burgess, G. C., Depue, B. E., Ruzic, L., Willcutt, E. G., Du, Y. P., & Banich, M. T. (2010). Attentional control activation relates to working memory in attention-deficit/hyperactivity disorder (ADHD). Biological Psychiatry, 67, 632–640. doi:10.1016/j.biopsych.2009.10.036
Buse, J., Schoenefeld, K., Munchau, A., & Roessner, V. (2013). Neuromodulation in Tourette syndrome: dopamine and beyond. Neuroscience and Biobehavioral Review, 37, 1069–1084. doi:10.1016/j.neubiorev.2012.10.004
Bush, G., Frazier, J. A., Rauch, S. L., Seidman, L. J., Whalen, P. J., Jenike, M. A., et al. (1999). Anterior cingulate cortex dysfunction in attention-deficit/hyperactivity disorder revealed by fMRI and the counting Stroop. Biological Psychiatry, 45, 1542–1552. doi:10.1016/S0006-3223(99)00083-9
Cai, W., George, J. S., Verbruggen, F., Chambers, C. D., & Aron, A. R. (2012). The role of the right presupplementary motor area in stopping action: two studies with event-related transcranial magnetic stimulation. Journal of Neurophysiology, 108, 380–389. doi:10.1152/jn.00132.2012
Casey, B. J., Castellanos, F. X., Giedd, J. N., March, W. L., Hamburger, S. D., Schubert, A. B., et al. (1997). Implication of right frontostriatal circuitry in response inhibition and attention-deficit/hyperactivity disorder. Journal of the American Academy of Child & Adolescent Psychiatry, 36, 374–383. doi:10.1097/00004583-199703000-00016
Chamberlain, S. R., Blackwell, A. D., Fineberg, N. A., Robbins, T. W., & Sahakian, B. J. (2005). The neuropsychology of obsessive-compulsive disorder: the importance of failures in cognitive and behavioural inhibition as candidate endophenotypic markers. Neuroscience and Biobehavioral Review, 29, 399–419. doi:10.1016/j.neubiorev.2004.11.006
Chamberlain, S. R., DelCampo, N., Dowson, J., Muller, U., Clark, L., Robbins, T. W., et al. (2007a). Atomoxetine improves response inhibition in adults with attention-deficit/hyperactivity disorder. Biological Psychiatry, 62, 977–984. doi:10.1016/j.biopsych.62,977–984
Chamberlain, S. R., Fineberg, N. A., Blackwell, A. D., Robbins, T. W., & Sahakian, B. J. (2006a). Motor inhibition and cognitive flexibility in obsessive-compulsive disorder and trichotillomania. American Journal of Psychiatry, 163, 1282–1284. doi:10.1176/appi.ajp.163.7.1282
Chamberlain, S. R., Fineberg, N. A., Menzies, L. A., Blackwell, A. D., Bullmore, E. T., Robbins, T. W., et al. (2007b). Impaired cognitive flexibility and motor inhibition in unaffected first-degree relatives of patients with obsessive-compulsive disorder. American Journal of Psychiatry, 164, 335–338. doi:10.1176/appi.ajp.164.2.335
Chamberlain, S. R., Muller, U., Blackwell, A. D., Clark, L., Robbins, T. W., & Sahakian, B. J. (2006b). Neurochemical modulation of response inhibition and probabilistic learning in humans. Science, 311, 861–863. doi:10.1126/science.1121218
Chambers, C. D., Bellgrove, M. A., Stokes, M. G., Henderson, T. R., Garavan, H., & Robertson, I. H. (2006). Executive “brake failure” following deactivation of human frontal lobe. Journal of Cognitive Neuroscience, 18, 444–455. doi:10.1162/089892906775990606
Chambers, C. D., Garavan, H., & Bellgrove, M. A. (2009). Insights into the neural basis of response inhibition from cognitive and clinical neuroscience. Neuroscience and Biobehavioral Review, 33, 631–646. doi:10.1016/j.neubiorev.2008.08.016
Channon, S., Drury, H., Martinos, M., Robertson, M. M., Orth, M., & Crawford, S. (2009). Tourette syndrome (TS): inhibitory performance in adults with uncomplicated TS. Neuropsychology, 23, 359–366. doi:10.1037/a0014552
Channon, S., Gunning, A., Frankl, J., & Robertson, M. M. (2006). Tourette syndrome (TS): cognitive performance in adults with uncomplicated TS. Neuropsychology, 20, 58–65. doi:10.1037/0894-4105.20.1.58
Chen, J., Lipska, B. K., Halim, N., Ma, Q. D., Matsumoto, M., Melhem, S., et al. (2004). Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in post-mortem human brain. American Journal of Human Genetics, 75, 807–821. doi:10.1086/425589
Cheon, K. A., Ryu, Y. H., Namkoong, K., Kim, C. H., Kim, J. J., & Lee, J. D. (2004). Dopamine transporter density of the basal ganglia assessed with [123I] IPTSPECT in drug-naive children with Tourette’s disorder. Psychiatry Research, 130, 85–95. doi:10.1016/j.pscychresns.2003.06.001
Clark, L., Roiser, J. P., Cools, R., Rubinsztein, D. C., Sahakian, B. J., & Robbins, T. W. (2005). Stop-signal response inhibition is not modulated by tryptophan depletion or serotonin transporter polymorphism in healthy volunteers: implications for the 5-HT theory of impulsivity. Psychopharmacology (Berl.), 182, 570–578. doi:10.1007/s00213-005-0104-6
Coghill, D. R., Seth, S., Pedroso, S., Usala, T., Currie, J., & Gagliano, A. (2013). Effects of methylphenidate on cognitive function in children and adolescents with attention-deficit/hyperactivity disorder: evidence from a systematic review and meta-analysis. Biological Psychiatry. doi:10.1016/j.biopsych.2013.10.005
Colzato, L. S., vandenWildenberg, W. P., VanderDoes, A. J., & Hommel, B. (2010). Genetic markers of striatal dopamine predict individual differences in dysfunctional, but not functional impulsivity. Neuroscience, 170, 782–788. doi:10.1016/j.neuroscience.2010.07.050
Congdon, E., Constable, R. T., Lesch, K. P., & Canli, T. (2009). Influence of SLC6A3 and COMT variation on neural activation during response inhibition. Biological Psychology, 81, 144–152. doi:10.1016/j.biopsycho.2009.03.005
Congdon, E., Lesch, K. P., & Canli, T. (2008). Analysis of DRD4 and DAT polymorphisms and behavioral inhibition in healthy adults: implications for impulsivity. American Journal of Medical Genetics. B Neuropsychiatr. Genet., 147, 27–32. doi:10.1002/ajmg.b.30557
Cools, R., & D’Esposito, M. (2011). Inverted-U-shaped dopamine actions on human working memory and cognitive control. Biological Psychiatry, 69, e113–e125. doi:10.1016/j.biopsych.2011.03.028
Cornish, K. M., Manly, T., Savage, R., Swanson, J., Morisano, D., Butler, N., et al. (2005). Association of the dopamine transporter (DAT1) 10/10-repeat genotype with ADHD symptoms and response inhibition in a general population sample. Molecular Psychiatry, 10, 686–698. doi:10.1038/sj.mp.4001641
Crawford, S., Channon, S., & Robertson, M. M. (2005). Tourette syndrome: performance on tests of behavioural inhibition, working memory, and gambling. Journal of Child Psychology and Psychiatry, 46, 1327–1336. doi:10.1111/j.1469-7610.2005.01419.x
Cubillo, A., Halari, R., Ecker, C., Giampietro, V., Taylor, E., & Rubia, K. (2010). Reduced activation and inter-regional functional connectivity of fronto-striatal networks in adults with childhood attention-deficit/hyperactivity disorder (ADHD) and persisting symptoms during tasks of motor inhibition and cogni-tive switching. Journal of Psychiatric Research, 44, 629–639. doi:10.1016/j.jpsychires.2009.11.016
Cubillo, A., Halari, R., Giampietro, V., Taylor, E., & Rubia, K. (2011). Fronto-striatal underactivation during interference inhibition and attention allocation in grownup children with attention-deficit/hyperactivity disorder and persisting symptoms. Psychiatry Research, 193, 17–27. doi:10.1016/j.pscychresns.2010.12.014
Cubillo, A., Smith, A. B., Barrett, N., Giampietro, V., Brammer, M. J., Simons,, A., et al. (2014). Shared and drug-specific effects of atomoxetine and methylphenidate on inhibitory brain dysfunction in medication-naive ADHD boys. Cerebral Cortex, 24, 174–185. doi:10.1093/cercor/bhs296
Cummins, T. D., Hawi, Z., Hocking, J., Strudwick, M., Hester, R., Garavan, H., et al. (2012). Dopamine transporter genotype predicts behavioural and neural measures of response inhibition. Molecular Psychiatry, 17, 1086–1092. doi:10.1038/mp.2011.104
Dalley, J. W., & Roiser, J. P. (2012). Dopamine, serotonin, and impulsivity. Neuroscience, 215, 42–58. doi:10.1016/j.neuroscience.2012.03.065
de Bartolomeis, A., Buonaguro, E., & Iasevoli, F. (2013). Serotonin-glutamate and serotonin-dopamine reciprocal interactions as putative molecular targets for novel antipsychotic treatments: from receptor heterodimers to post-synaptic scaffolding and effector proteins. Psychopharmacology, 225, 1–19. doi:10.1007/s00213-012-2921-8
de Vries, F. E., de Wit, S. J., Cath, D. C., vander Werf, Y. D., vander Borden, V., van Rossum, T. B., et al. (2013). Compensatory frontoparietal activity during working memory: an endophenotype of obsessive-compulsive disorder. Biological Psychiatry. doi:10.1016/j.biopsych.2013.11.021
de Wit, S. J., de Vries, F. E., vander Werf, Y. D., Cath, D. C., Heslenfeld, D. J., Veltman, E. M., et al. (2012). Presupplementary motor area hyperactivity during response inhibition: a candidate endophenotype of obsessive-compulsive disorder. American Journal of Psychiatry, 169, 1100–1108. doi:10.1176/appi.ajp.2012.1201007
Denys, D., de Geus, F., van Megen, H. J., & Westenberg, H. G. (2004a). A double-blind, randomized, placebo-controlled trial of quetiapine addition in patients with obsessive-compulsive disorder refractory to serotonin reuptake inhibitors. Journal of Clinical Psychiatry, 65, 1040–1048. doi:10.4088/JCP.v65n0803
Denys, D., van der Wee, N., Janssen, J., DeGeus, F., & Westenberg, H. G. (2004b). Low level of dopaminergic D2 receptor binding in obsessive-compulsive disorder. Biological Psychiatry, 55, 1041–1045. doi:10.1016/j.biopsych.2004.01.023
Depue, B. E., Burgess, G. C., Bidwell, L. C., Willcutt, E. G., & Banich, M. T. (2010). Behavioral performance predicts gray matter reductions in right inferior frontal gyrus in young adults with combined-type ADHD. Journal of Psychiatric Research, 182, 231–237. doi:10.1016/j.pscychresns.2010.01.012
DeVito, E. E., Blackwell, A. D., Clark, L., Kent, L., Dezsery, A. M., Turner, D. C., et al. (2009). Methylphenidate improves response inhibition but not reflection impulsivity in children with attention-deficit/hyperactivity disorder (ADHD). Psychopharmacology (Berl.), 202, 531–539. doi:10.1007/s00213-008-1337-y
DeVito, T. J., Drost, D. J., Pavlosky, W., Neufeld, R. W., Rajakumar, N., McKinlay, B. D., et al. (2005). Brain magnetic resonance spectroscopy in Tourette’s disorder. Journal of the American Academy of Child & Adolescent Psychiatry, 44, 1301–1308. doi:10.1097/01.chi.0000181046.52078
Dibbets, P., Evers, L., Hurks, P., Marchetta, N., & Jolles, J. (2009). Differences in feedback and inhibition-related neural activity in adult ADHD. Brain and Cognition, 70, 73–83. doi:10.1016/j.bandc.2009.01.001
Dillo, W., Goke, A., Prox-Vagedes, V., Szycik, G. R., Roy, M., Donnerstag, F., et al. (2010). Neuronal correlates of ADHD in adult swith evidence for compensation strategies–a functional MRI study with a Go/No-Go paradigm. German Medical Science, 8, Doc09. doi:10.3205/000098
Donders, F. C. (1969). On the speed of mental processes. Acta Psychologica, 30, 412–431. [Originally published in Dutch, translated by W. G. Koster]
Draganski, B., Martino, D., Cavanna, A. E., Hutton, C., Orth, M., Robertson, M. M., et al. (2010). Multispectral brain morphometry in Tourette syndrome persisting into adulthood. Brain, 133 (Pt12), 3661–3675. doi:10.1093/brain/awq300
Drueke, B., Boecker, M., Schlaegel, S., Moeller, O., Hiemke, C., Grunder, G., et al. (2010). Serotonergic modulation of response inhibition and re-engagement? Results of a study in healthy human volunteers. Human Psychopharmacology, 25, 472–480. doi:10.1002/hup.1141
Durston, S. (2003). A review of the biological bases of ADHD: what have we learned from imaging studies? Mental Retardation and Developmental Disability Reviews, 9, 184–195. doi:10.1002/mrdd.10079
Durston, S., Fossella, J. A., Mulder, M. J., Casey, B. J., Ziermans, T. B., Vessaz, M. N., et al. (2008). Dopamine transporter genotype conveys familial risk of attention-deficit/hyperactivity disorder through striatal activation. Journal of the American Academy of Child & Adolescent Psychiatry, 47, 61–67. doi:10.1097/chi.0b013e31815a5f17
Durston, S., Tottenham, N. T., Thomas, K. M., Davidson, M. C., Eigsti, I. M., Yang, Y., et al. (2003). Differential patterns of striatal activation in young children with and without ADHD. Biological Psychiatry, 53, 871–878. doi:10.1006/S0006-3223(02)01904-2
Eagle, D. M., Bari, A., & Robbins, T. W. (2008). The neuropsychopharmacology of action inhibition: cross-species translation of the stop-signal and go/no-go tasks. Psychopharmacology (Berl.), 199, 439–456. doi:10.1007/s00213-008-1127-6
Eagle, D. M., Lehmann,, O., Theobald, D. E., Pena, Y., Zakaria, R., Ghosh, R., et al. (2009). Serotonin depletion impairs waiting but not stop-signal reaction time in rats: implications for the theories of the role of 5-HT in behavioral inhibition. Neuropsychopharmacology, 34, 1311–1321. doi:10.1038/npp.2008.202
Eagle, D. M., Tufft, M. R., Goodchild, H. L., & Robbins, T. W. (2007). Differential effects of modafinil and methylphenidate on stop-signal reaction time task performance in the rat and interactions with the dopamine receptor antagonist cis-flupenthixol. Psychopharmacology (Berl.), 192, 193–206. doi:10.1007/s00213-007-0701-7
Eichele, H., Eichele, T., Hammar, A., Freyberger, H. J., Hugdahl, K., and Plessen, K. J. (2010). Go/NoGo performance in boys with Tourette syndrome. Child Neuropsychology, 16, 162–168. doi:10.1080/09297040903150182
Evers, E. A., vander Veen, F. M., van Deursen, J. A., Schmitt, J. A., Deutz, N. E., and Jolles, J. (2006). The effect of acute tryptophan depletion on the BOLD response during performance monitoring and response inhibition in healthy male volunteers. Psychopharmacology (Berl.), 187, 200–208. doi:10.1007/s00213-006-0411-6
Felling, R. J., & Singer, H. S. (2011). Neurobiology of Tourette syndrome: current status and need for further investigation. Journal of Neuroscience, 31, 12387–12395. doi:10.1523/JNEUROSCI.0150-11.2011
Fineberg, N. A., Sivakumaran, T., Roberts, A., and Gale, T. (2005). Adding quetiapine to SRI in treatment-resistant obsessive-compulsive disorder: a randomized controlled treatment study. International Journal of Clinical Psychopharmacology, 20, 223–226. doi:10.1097/00004850-200507000-00005
Fitzgerald, K. D., Welsh, R. C., Gehring, W. J., Abelson, J. L., Himle, J. A., et al. (2005). Error-related hyperactivity of the anterior cingulate cortex in obsessive-compulsive disorder. Biological Psychiatry, 57, 287–294. doi:10.1016/j.biopsych.2004.10.038
Floden, D., & Stuss, D. T. (2006). Inhibitory control is slowed in patients with right superior medial frontal damage. Journal of Cognitive Neuroscience, 18, 1843–1849. doi:10.1162/jocn.2006.18.11.1843
Frodl, T., & Skokauskas, N. (2012). Meta-analysis of structural MRI studies in children and adolescents with attention-deficit/hyperactivity disorder indicates treatment effects. Acta Psychiatrica Scandinavica, 125, 114–126. doi:10.1111/j.1600-0447.2011.01786.x
Froehlich, T. E., Lanphear, B. P., Epstein, J. N., Barbaresi, W. J., Katasi, S. K., and Kahn, R. S. (2007). Prevalence, recognition, and treatment of attention-deficit/hyperactivity disorder in a national sample of US children. Archive of Pediatrics and Adolescent Medicine, 161, 857–864. doi:10.1001/archpedi.161.9.857
Gadde, K. M., Ryan Wagner, H. II, Connor, K. M., and Foust, M. S. (2007). Escitalopram treatment of trichotillomania. International Journal of Clinical Psychopharmacology, 22, 39–42. doi:10.1097/01.yic.0000224799.59524.50
Gerlach, M., Grunblatt, E., Lange, K. W. (2013). Is the treatment with psycho-stimulants in children and adolescents with attention-deficit/hyperactivity disorder harmful for the dopaminergic system? Attention Deficit Hyperactivity Disorder, 5, 71–81. doi:10.1007/s12402-013-0105-y
Ghahremani, D. G., Lee, B., Robertson, C. L., Tabibnia, G., Morgan, A. T., DeShetler, N., et al. (2012). Striatal dopamine D(2)/D(3) receptors mediate response inhibition and related activity in frontostriatal neural circuitry in humans. Journal of Neuroscience, 32, 7316–7324. doi:10.1523/JNEUROSCI.4284-11.2012
Ghanizadeh, A., & Mosallaei, S. (2009). Psychiatric disorders and behavioral problems in children and adolescents with Tourette syndrome. Brain Development, 31, 15–19. doi:10.1016/j.braindev.2008.03.010
Gilbert, D. L., Christian, B. T., Gelfand, M. J., Shi, B., Mantil, J., and Sallee, F. R. (2006). Altered mesolimbocortical and thalamic dopamine in Tourette syndrome. Neurology, 67, 1695–1697. doi:10.1212/01.wnl.0000242733.18534.2c
Gottesman, I. I., & Gould, T. D. (2003). The endophenotype concept in psychiatry: etymology and strategic intentions. American Journal of Psychiatry, 160, 636–645. doi:10.1176/appi.ajp.160.4.636
Grachev, I. D. (1997). MRI-based morphotopographic parcellation of human neocortex in trichotillomania. Psychiatry Clinical Neuroscience, 51, 315–321. doi:10.1111/j.1440-1819.1997.tb03205
Hart, H., Radua, J., Nakao, T., Mataix-Cols, D., and Rubia, K. (2012). Meta-analysis of functional magnetic resonance imaging studies of inhibition and attention in attention-deficit/hyperactivity disorder: exploring task-specific, stimulant medication, and age effects. Archive of General Psychiatry, 70, 1–14. doi:10.1001/jamapsychiatry.2013.277
Haugbol, S., Pinborg, L. H., Regeur, L., Hansen, E. S., Bolwig, T. G., Nielsen, F. A., et al. (2007). Cerebral 5-HT2A receptor binding is increased in patients with Tourette syndrome. International Journal of Neuropsychopharmacology, 10, 245–252. doi:10.1017/S146114570600
